
Short and Long-Term Effects of Early and Late Weaning on Atlantic Cod, Gadus morhua


Abstract
1. Introduction
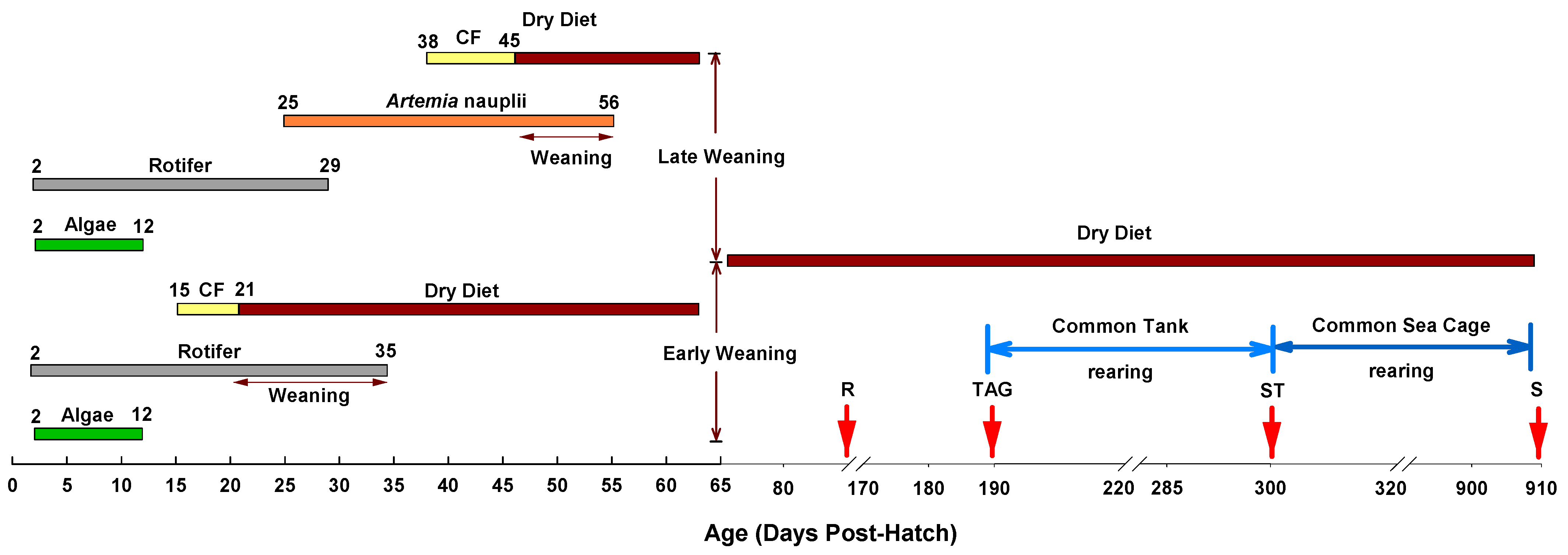
2. Materials and Methods
2.1. Gamete Collection and Egg Incubation
2.2. Larval Rearing
2.3. Data Collection and Analysis
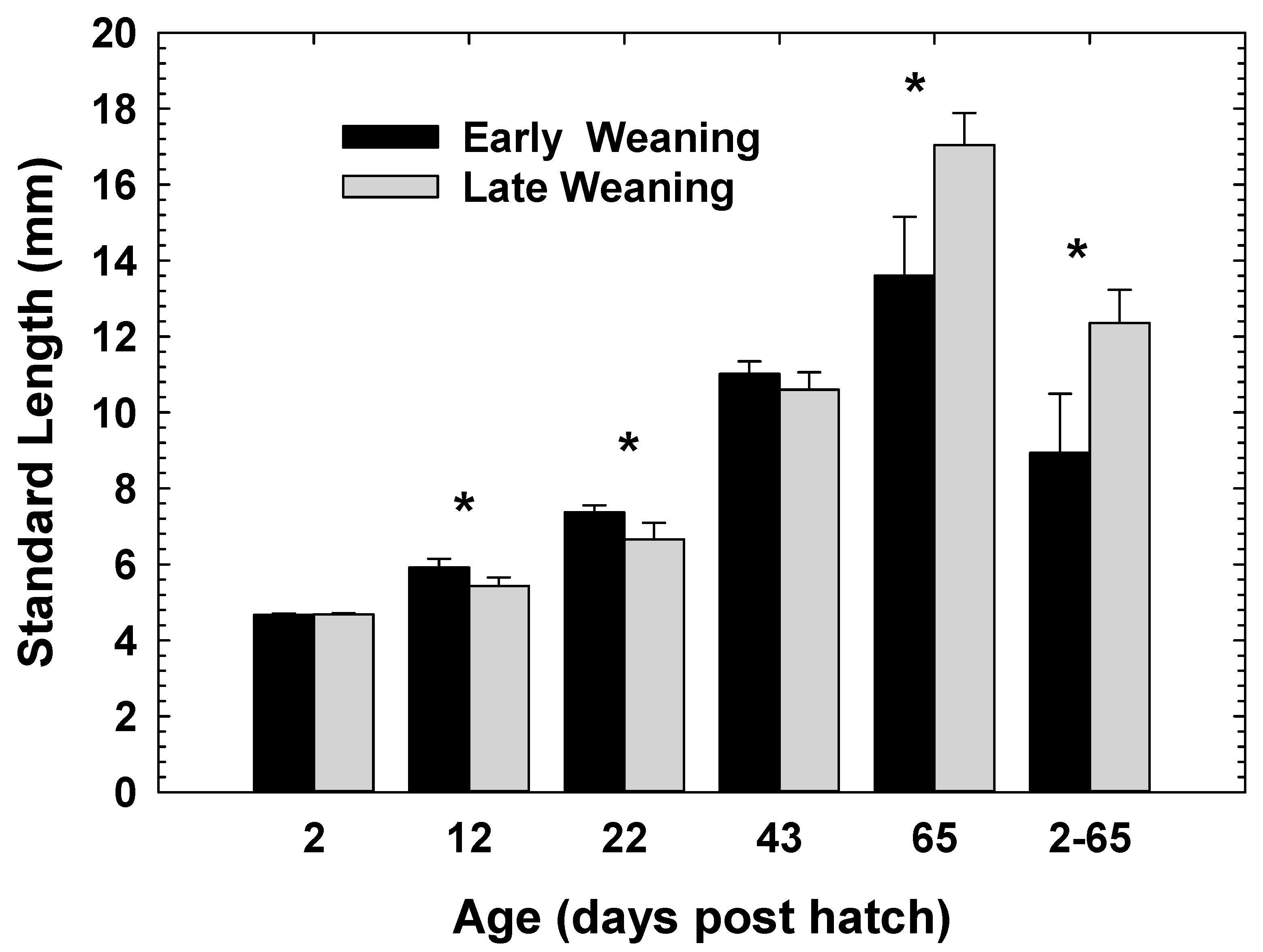
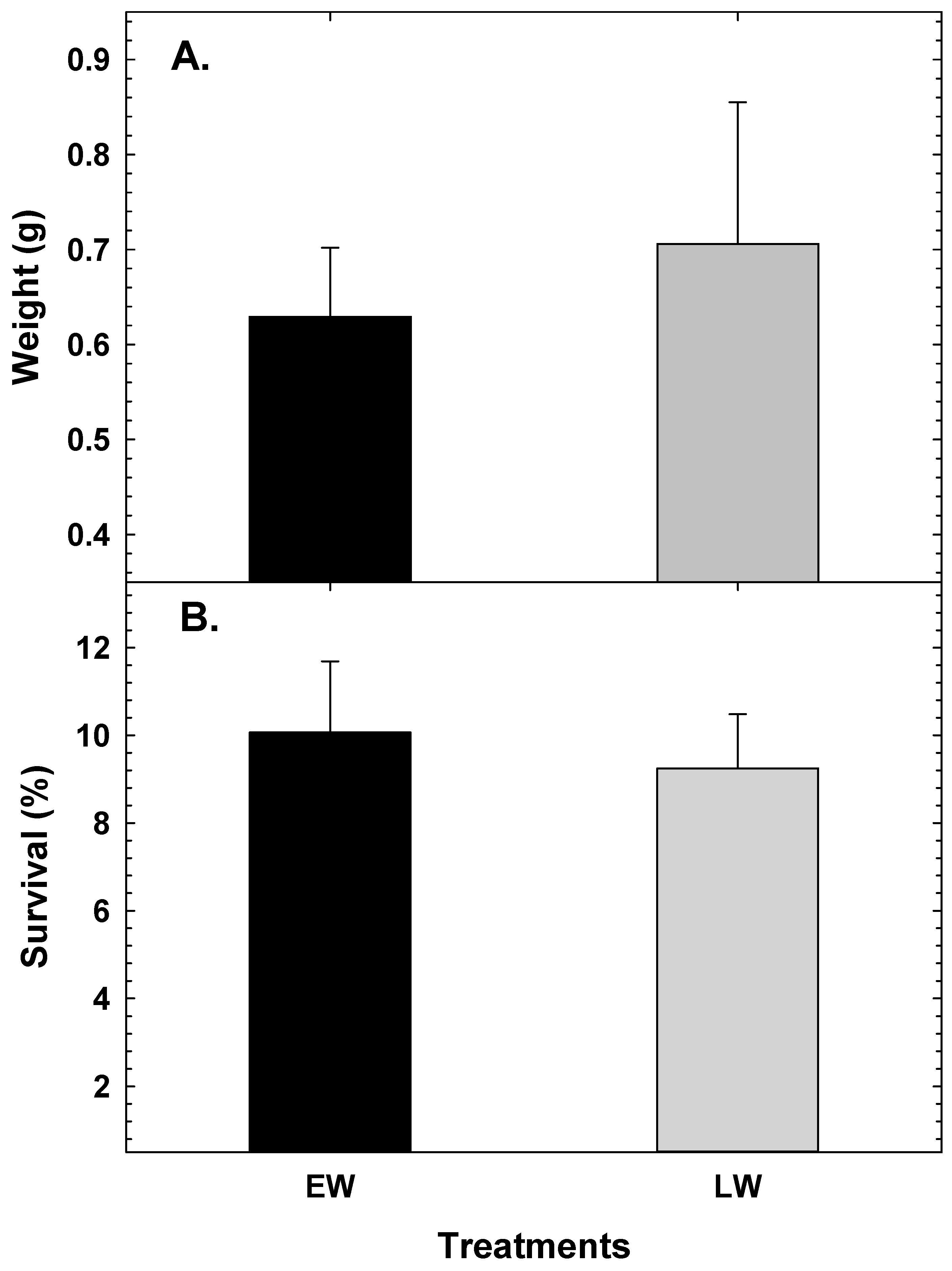
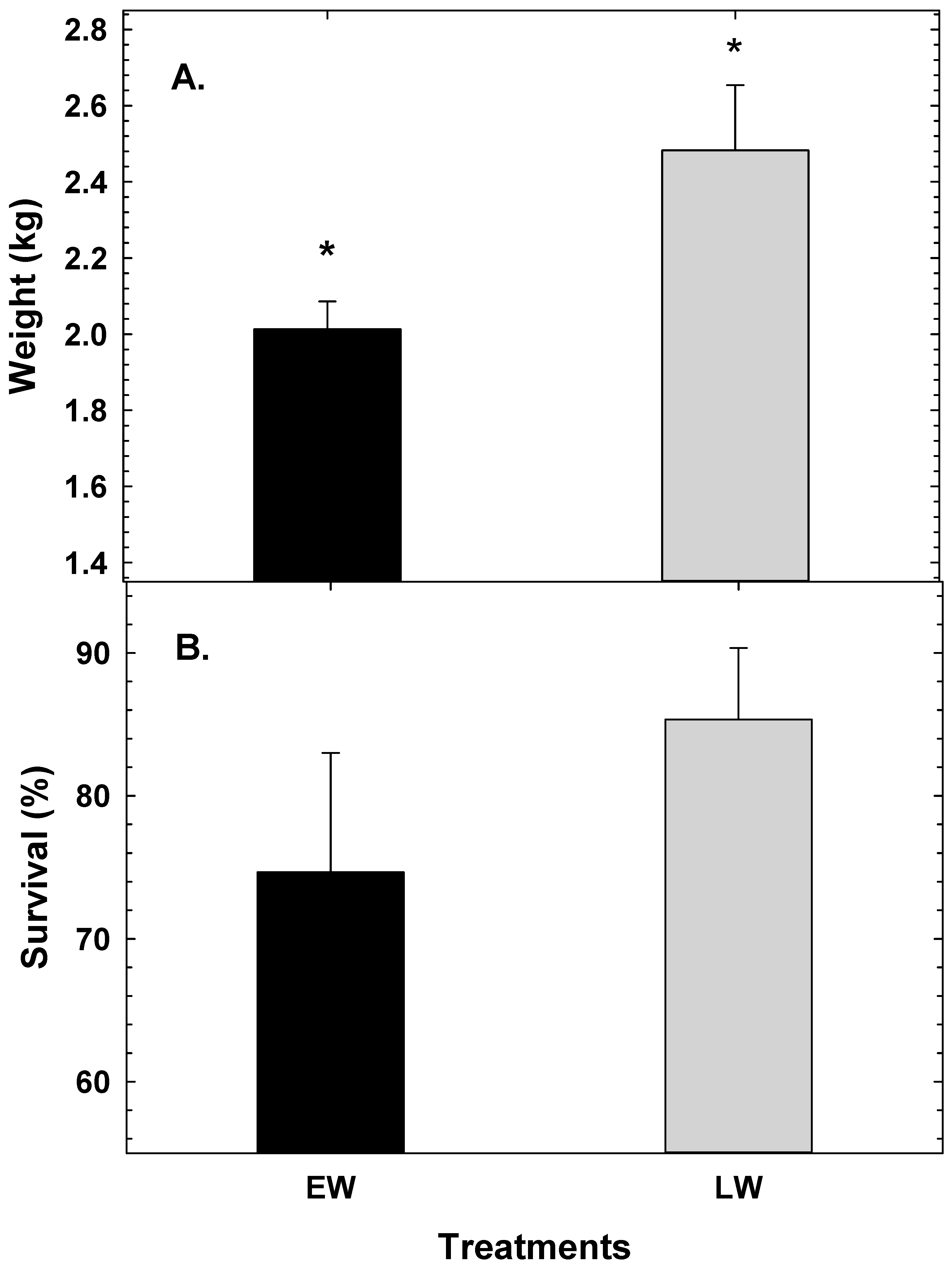
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cahu, C.; Infante, J.Z. Substitution of live food by formulated diets in marine fish larvae. Aquaculture 2001, 200, 161–180. [Google Scholar] [CrossRef]
- Kolkovski, S.; Tandler, A.; Izquierdo, M.S. The effects of live food and dietary digestive enzymes on the efficiency of microdiets for seabass Dicentrarchus labrax. larvae. Aquaculture 1997, 148, 313–322. [Google Scholar] [CrossRef]
- Moguel-Hernández, I.; Peña, R.; Andree, K.B.; Tovar-Ramirez, D.; Bonacic, K.; Dumas, S.; Gisbert, E. Ontogeny changes and weaning effects in gene expression patterns of digestive enzymes and regulatory digestive factors in spotted rose snapper (Lutjanus guttatus) larvae. Fish Physiol. Biochem. 2016, 42, 1319–1334. [Google Scholar] [CrossRef] [PubMed]
- Sorgeloos, P.; Dhert, P.; Candreva, P. Use of the brine shrimp, Artemia spp., in marine fish larviculture. Aquaculture 2001, 200, 147–159. [Google Scholar]
- Navarro, J.C.; Henderson, R.J.; McEvoy, L.A.; Bell, M.V.; Amat, F. Lipid conversions during enrichment of Artemia. Aquaculture 1999, 174, 155–166. [Google Scholar] [CrossRef]
- Baskerville-Bridges, B.; Kling, L.J. Early weaning of Atlantic cod (Gadus morhua) larvae onto a microparticulate diet. Aquaculture 2000, 189, 109–117. [Google Scholar]
- Næss, T.; Hamre, K.; Holm, J.C. Successful early weaning of Atlantic halibut (Hippoglossus hippoglossus L.) in small shallow raceway systems. Aquacult. Res. 2001, 32, 163–168. [Google Scholar]
- Fletcher Jr, R.C.; Roy, W.; Davie, A.; Taylor, J.; Robertson, D.; Migaud, H. Evaluation of new microparticulate diets for early weaning of Atlantic cod (Gadus morhua): Implications on larval performances and tank hygiene. Aquaculture 2007, 263, 35–51. [Google Scholar]
- Bonaldo, A.; Parma, L.; Badiani, A.; Serratore, P.; Gatta, P.P. Very early weaning of common sole (Solea solea L.) larvae by means of different feeding regimes and three commercial microdiets: Influence on performances, metamorphosis development and tank hygiene. Aquaculture 2011, 321, 237–244. [Google Scholar] [CrossRef]
- Brown, J.A.; Minkoff, G.; Puvanendran, V. Larviculture of Atlantic cod (Gadus morhua)—Progress, protocols and problems. Aquaculture 2003, 227, 357–372. [Google Scholar] [CrossRef]
- Rosenlund, G.; Halldórsson, Ó. Cod juvenile production: Research and commercial developments. Aquaculture 2007, 268, 188–194. [Google Scholar] [CrossRef]
- Polanco, J.F.; Bjørndal, T. Aquaculture diversification in Europe: The Kingdom of Spain and the Kingdom of Norway. In Planning for Aquaculture Diversification: The Importance of Climate Change and Other Driver; Harvey, B., Soto, D., Carolsfeld, J., Beveridge, M., Bartley, D.M., Eds.; FAO Fisheries and Aquaculture Proceedings No. 47; FAO: Rome, Italy, 2017; pp. 37–45. [Google Scholar]
- Bangera, R.; Ødegård, J.; Præbel, A.K.; Mortensen, A.; Nielsen, H.M. Genetic correlations between growth rate and resistance to vibriosis and viral nervous necrosis in Atlantic cod (Gadus morhua L.). Aquaculture 2011, 317, 67–73. [Google Scholar] [CrossRef]
- Bangera, R.; Ødegård, J.; Mikkelsen, H.; Nielsen, H.M.; Seppola, M.; Puvanendran, V.; Gjøen, H.M.; Hansen, Ø.J.; Mortensen, A. Genetic analysis of francisellosis field outbreak in Atlantic cod (Gadus morhua L.) using an ordinal threshold model. Aquaculture 2014, 420, S50–S56. [Google Scholar] [CrossRef]
- Bangera, R.; Drangsholt, T.M.; Nielsen, H.M.; Sae-Lim, P.; Ødegård, J.; Puvanendran, V.; Hansen, Ø.J.; Mortensen, A. Genotype by environment interaction for growth in Atlantic cod (Gadus morhua L.) in four farms of Norway. J. Mar. Sci. Eng. 2015, 3, 412–427. [Google Scholar] [CrossRef]
- Hansen, Ø.J.; Puvanendran, V.; Jøstensen, J.P.; Ous, C. Effects of dietary levels and ratio of phosphatidylcholine and phosphatidylinositol on the growth, survival and deformity levels of Atlantic cod larvae and early juveniles. Aquacult. Res. 2011, 42, 1026–1033. [Google Scholar] [CrossRef]
- Hansen, Ø.J.; Puvanendran, V.; Mortensen, A. Importance of broodstock holding temperature on fecundity and egg quality in three groups of photo-manipulated Atlantic cod broodstock. Aquacult. Res. 2012, 44, 140–150. [Google Scholar] [CrossRef]
- Hansen, Ø.J.; Puvanendran, V.; Bangera, R. Do maternal age and experience contribute to better growth, survival and disease resistance of offspring in Atlantic cod (Gadus morhua)? Aquacult. Int. 2015, 23, 1157–1164. [Google Scholar] [CrossRef]
- Hansen, Ø.J.; Puvanendran, V.; Bangera, R. Broodstock diet with water and astaxanthin improve condition and egg output of brood fish and larval survival in Atlantic cod, Gadus morhua L. Aquacult. Res. 2016, 47, 819–829. [Google Scholar] [CrossRef]
- Hansen, Ø.J.; Puvanendran, V.; Jøstensen, J.P.; Falk-Petersen, I.B. Early introduction of an inert diet and unenriched Artemia enhances growth and quality of Atlantic cod (Gadus morhua) larvae. Aquacult. Nutr. 2018, 24, 102–111. [Google Scholar] [CrossRef]
- Puvanendran, V.; Mortensen, A.; Johansen, L.H.; Kettunen, A.; Hansen, Ø.J.; Henriksen, E.; Heide, M. Development of cod farming in Norway: Past and current biological and market status and future prospects and directions. Rev. Aquacult. 2022, 14, 308–342. [Google Scholar] [CrossRef]
- Callan, C.; Jordaan, A.; Kling, L.J. Reducing Artemia use in the culture of Atlantic cod (Gadus morhua). Aquaculture 2003, 219, 585–595. [Google Scholar] [CrossRef]
- Hansen, Ø.J.; Puvanendran, V. Fertilization success and blastomere morphology as predictors of egg and juvenile quality for domesticated Atlantic cod, Gadus morhua, broodstock. Aquacult. Res. 2010, 41, 1791–1798. [Google Scholar] [CrossRef]
- Björnsson, B.; Ólafsdóttir, S.R. Effects of water quality and stocking density on growth performance of juvenile cod (Gadus morhua L.). ICES J. Mar. Sci. 2006, 63, 326–334. [Google Scholar] [CrossRef]
- Brekken, B. Kvantitativ Bestemmelse av Frie Aminosyrer; SSF-Rapport: Stockholm, Sweden, 1989; A-142; 13p. [Google Scholar]
- Cohen, S.A.; Meys, M.; Tarvin, T.L. The Pico Tag® Method. A Manual of Advanced Techniques for Amino Acid Analysis. In Water Division of Millipore WM02, Rev 1; Millipore Corporation: Milford, MA, USA, 1989; 123p. [Google Scholar]
- Conover, W.J.; Iman, R.L. Rank transformations as a bridge between parametric and nonparametric statistics. Am. Stat. 1981, 35, 124–129. [Google Scholar]
- Faulk, C.K.; Holt, G.J. Early weaning of southern flounder, Paralichthys lethostigma, larvae and ontogeny of selected digestive enzymes. Aquaculture 2009, 296, 213–218. [Google Scholar] [CrossRef]
- Parma, L.; Bonaldo, A.; Massi, P.; Yúfera, M.; Martínez-Rodríguez, G.; Gatta, P.P. Different early weaning protocols in common sole (Solea solea L.) larvae: Implications on the performances and molecular ontogeny of digestive enzyme precursors. Aquaculture 2013, 414, 26–35. [Google Scholar] [CrossRef]
- Mata-Sotres, J.A.; Lazo, J.P.; Baron-Sevilla, B. Effect of age on weaning success in totoaba (Totoaba macdonaldi) larval culture. Aquaculture 2015, 437, 292–296. [Google Scholar] [CrossRef]
- Nhu, V.C.; Dierckens, K.; Nguyen, H.T.; Hoang, T.M.T.; Le, T.L.; Tran, M.T.; Nys, C.; Sorgeloos, P. Effect of early co-feeding and different weaning diets on the performance of cobia (Rachycentron canadum) larvae and juveniles. Aquaculture 2010, 305, 52–58. [Google Scholar] [CrossRef]
- Nguyen, H.Q.; Reinertsen, H.; Wold, P.A.; Tran, T.M.; Kjørsvik, E. Effects of early weaning strategies on growth, survival and digestive enzyme activities in cobia (Rachycentron canadum L.) larvae. Aquacult. Int. 2011, 19, 63–78. [Google Scholar] [CrossRef]
- Folkvord, A.; Otterå, H. Effects of initial size distribution, day length, and feeding frequency on growth, survival, and cannibalism in juvenile Atlantic cod (Gadus morhua L.). Aquaculture 1993, 114, 243–260. [Google Scholar] [CrossRef]
- Puvanendran, V.; Laurel, B.J.; Brown, J.A. Cannibalism of Atlantic cod Gadus morhua larvae and juveniles on first-week larvae. Aquat. Biol. 2008, 2, 113–118. [Google Scholar] [CrossRef]
- Canavate, J.P.; Fernández-Dıaz, C. Influence of co-feeding larvae with live and inert diets on weaning the sole Solea senegalensis onto commercial dry feeds. Aquaculture 1999, 174, 255–263. [Google Scholar] [CrossRef]
- Pousão-Ferreira, P.; Santos, P.; Carvalho, A.P.; Morais, S.; Narciso, L. Effect of an experimental microparticulate diet on the growth, survival and fatty acid profile of gilthead seabream (Sparus aurata L.) larvae. Aquacult. Int. 2003, 11, 491–504. [Google Scholar] [CrossRef]
- Cahu, C.L.; Infante, J.Z. Early weaning of sea bass (Dicentrarchus labrax) larvae with a compound diet: Effect on digestive enzymes. Comp. Biochem. Physiol. 1994, 109, 213–222. [Google Scholar] [CrossRef]
- Ma, Z.; Qin, J.G.; Hutchinson, W.; Chen, B.N.; Song, L. Responses of digestive enzymes and body lipids to weaning times in yellowtail kingfish Seriola lalandi (Valenciennes, 1833) larvae. Aquacult. Res. 2014, 45, 973–982. [Google Scholar] [CrossRef]
- Guerreiro, I.; de Vareilles, M.; Pousão-Ferreira, P.; Rodrigues, V.; Dinis, M.T.; Ribeiro, L. Effect of age-at-weaning on digestive capacity of white seabream (Diplodus sargus). Aquaculture 2010, 300, 194–205. [Google Scholar] [CrossRef]
- Kolkovski, S.; Koven, W.; Tandler, A. The mode of action of Artemia in enhancing utilization of microdiet by gilthead seabream Sparus aurata larvae. Aquaculture 1997, 155, 193–205. [Google Scholar] [CrossRef]
- Naz, M.; Turkmen, M. Digestive enzymes and hormones in gilthead seabream larvae (Sparus aurata) fed Artemia nauplii enriched with free histidine. Isr. J. Aquac. 2008, 60, 230–236. [Google Scholar] [CrossRef]
- Naz, M.; Türkmen, M. The changes in digestive enzymes and hormones of gilthead seabream larvae (Sparus aurata, L 1758) fed on Artemia nauplii enriched with free methionine. Aquacult. Int. 2009, 17, 243–256. [Google Scholar] [CrossRef]
- Batten, T.F.C.; Camere, M.L.; Moons, L.; Vandesande, F. Comparative distribution of neuropeptide-immunoreactive systems in the brain of the greenmolly, Poecillia latipinna. J. Comp. Neurol. 1990, 302, 893–919. [Google Scholar] [CrossRef]
- Thorndyke, M.; Holmgren, S. Bombesin potentiates the effect of acetylcholine on isolated strips of fish stomach. Regul. Pept. 1990, 30, 125–135. [Google Scholar] [CrossRef] [PubMed]
- McDonald, T.J.; Jornvall, H.; Nilsson, G.; Vagne, M.; Ghatei, M.; Bloom, S.R.; Mutt, V. Characterization of a gastrin releasing peptide from porcine non-antral gastric tissue. Biochem. Biophys. Res. Commun. 1979, 90, 227–233. [Google Scholar] [CrossRef]
- Curnow, J.; King, J.; Partridge, G.; Kolkovski, S. Effects of two commercial microdiets on growth and survival of barramundi (Lates calcarifer Bloch) larvae within various early weaning protocols. Aquacult. Nutr. 2006, 12, 247–255. [Google Scholar] [CrossRef]
- Koedijk, R.M.; Folkvord, A.; Foss, A.; Pittman, K.; Stefansson, S.O.; Handeland, S.; Imsland, A.K. The influence of first-feeding diet on the Atlantic cod Gadus morhua phenotype: Survival, development and long-term consequences for growth. J. Fish Biol. 2010, 77, 1–19. [Google Scholar] [CrossRef]
- Imsland, A.K.; Foss, A.; Koedijk, R.; Folkvord, A.; Stefansson, S.O.; Jonassen, T.M. Short-and long-term differences in growth, feed conversion efficiency and deformities in juvenile Atlantic cod (Gadus morhua) startfed on rotifers or zooplankton. Aquacult. Res. 2006, 37, 1015–1027. [Google Scholar] [CrossRef]
- Aragão, C.; Conceição, L.E.; Dinis, M.T.; Fyhn, H.J. Amino acid pools of rotifers and Artemia under different conditions: Nutritional implications for fish larvae. Aquaculture 2004, 234, 429–445. [Google Scholar] [CrossRef]
- Naz, M. The changes in the biochemical compositions and enzymatic activities of rotifer (Brachionus plicatilis, Müller) and Artemia during the enrichment and starvation periods. Fish Physiol. Biochem. 2008, 34, 391–404. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zheng, S.; Wu, G. Nutrition and functions of amino acids in fish. In Amino Acids in Nutrition and Health, Advances in Experimental Medicine and Biology; Wu, G., Ed.; Springer: Cham, Switzerland, 2021; pp. 133–168. [Google Scholar]
- Salze, G.P.; Davis, D.A. Taurine: A critical nutrient for future fish feeds. Aquaculture 2015, 437, 215–229. [Google Scholar] [CrossRef]
- Kim, S.-K.; Takeuchi, T.; Akimoto, A.; Furuita, H.; Yamamoto, T.; Yokoyama, M.; Murata, Y. Effect of taurine supplemented practical diet on growth performance and taurine contents in whole body and tissues of juvenile Japanese flounder Paralichthys olivaceus. Fish. Sci. 2005, 71, 627–632. [Google Scholar] [CrossRef]
- Matsunari, H.; Yamamoto, T.; Kim, S.-K.; Goto, T.; Takeuchi, T. Optimum dietary taurine level in casein-based diet for juvenile red sea bream Pagrus major. Fish. Sci. 2008, 74, 347–353. [Google Scholar] [CrossRef]
- Takagi, S.; Murata, H.; Goto, T.; Endo, M.; Yamashita, H.; Ukawa, M. Taurine is an essential nutrient for yellowtail Seriola quinqueradiata fed non-fish meal diets based on soy protein concentrate. Aquaculture 2008, 280, 198–205. [Google Scholar] [CrossRef]
- Karlsen, Ø.; van der Meeren, T.; Rønnestad, I.; Mangor-Jensen, A.; Galloway, T.F.; Kjørsvik, E.; Hamre, K. Copepods enhance nutritional status, growth and development in Atlantic cod (Gadus morhua L.) larvae—Can we identify the underlying factors? PeerJ 2015, 3, e902. [Google Scholar] [CrossRef] [PubMed]
- Sutherland, R. Economics of Potential Systems for Farmed Production of Cod. 1998. 36p. Available online: https://www.seafish.org/media/Publications/EconomicsOfPotentialforFarmedProductionofCod.pdf (accessed on 25 April 2023).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | Unit | Amount |
|---|---|---|
| Protein | % | 58.9 |
| Fat | % | 16.5 |
| Ash | % | 9.4 |
| Vit. A | % | 0.005 |
| Vit. C | % | 0.073 |
| Vit. D | % | 0.000015 |
| Vit. E | % | 0.03 |
| Taurine | % | 0.4 |
| Energy | Jkg−1 | 1605 |
| DHA | % fat | 13.7 |
| EPA | % fat | 7.0 |
| ARA | % fat | 0.43 |
| PL | % fat | 32.1 |
| DHA | % PL | 22.9 |
| EPA | % PL | 9.9 |
| ARA | % PL | 0.54 |
| Lipid classes in total lipid | ||
| PC | % fat | 18.8 |
| 2-LPC | % fat | 1.6 |
| PI | % fat | 2.57 |
| PE | % fat | 5.87 |
| PA | % fat | 1.0 |
| FAA (g. 100 g Sample −1) | Rotifer | Artemia |
|---|---|---|
| Creatinine | 0.004 | 0.0035 |
| Aspartic acid | 0.0255 * | 0.0475 * |
| Glutamic acid | 0.065 * | 0.074 * |
| Hydroxyproline | ND | 0.001 |
| Serine | 0.0435 * | 0.0455 * |
| Asparagine | 0.0345 * | 0.04 * |
| Glycine | 0.0235 * | 0.029 * |
| Glutamine | 0.0735 * | 0.0685 * |
| 3-amino-propanic acid | 0.001 | 0.001 |
| Taurine | 0.011 * | 0.0345 * |
| Histidine | 0.017 * | 0.023 * |
| 4-amino-butanoic acid | 0.001 | 0.001 |
| Citrulline | 0.004 * | 0.001 * |
| Threonine | 0.0285 * | 0.041 * |
| Alanine | 0.0565 * | 0.0675 * |
| Carnosine | ND | ND |
| Arginine | 0.057 * | 0.088 * |
| Proline | 0.03 * | 0.044 * |
| Tyrosine | 0.0315 * | 0.05 * |
| Valine | 0.034 * | 0.0555 * |
| Methionine | 0.021 * | 0.0265 * |
| Cysteine | ND | ND |
| Isoleucine | 0.0285 * | 0.0475 * |
| Leucine | 0.0635 * | 0.086 * |
| Phenylalanine | 0.041 * | 0.052 * |
| Tryptophan | 0.007 * | 0.011 * |
| Ornithine | 0.004 | 0.005 |
| Lysine | 0.0795 * | 0.119 * |
| Categories | |
|---|---|
| Number of Artemia needed (billion) | 1.9 |
| Number of Artemia cysts in 1 kg (million) | 320 |
| Amount of Artemia cyst needed (kg) | 65 |
| Price of Artemia cyst (USD/kg) | 96 |
| Total cost of Artemia cyst (USD) | 6240 |
| Artemia enrichment cost (USD) | 3000 |
| Artemia capital cost (tanks, equipments, energy, etc.; USD) | 10,000 |
| Total Artemia cost (USD) | 19,240 |
| Labour cost (2 persons for 1 month; USD) | 7000 |
| Total Artemia Production cost (USD) | 26,240 |
| Categories | |
|---|---|
| Average weight (kg) of adult cod from LW | 2.48 |
| Average weight (kg) of adult cod from EW | 2.01 |
| Average difference in weight (kg) | 0.47 |
| Number of fish at slaughter (80% survival) | 900,000 |
| Total weight gain using LW (kg) | 423,000 |
| Estimated gate price of the cod (USD) | 4 |
| Total price gain using LW (USD) | 1,692,000 |
| Total profit (minus Artemia production cost; USD) | 1,665,760 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Puvanendran, V.; Hansen, Ø.J. Short and Long-Term Effects of Early and Late Weaning on Atlantic Cod, Gadus morhua. Fishes 2023, 8, 312. https://doi.org/10.3390/fishes8060312
Puvanendran V, Hansen ØJ. Short and Long-Term Effects of Early and Late Weaning on Atlantic Cod, Gadus morhua. Fishes. 2023; 8(6):312. https://doi.org/10.3390/fishes8060312
Chicago/Turabian StylePuvanendran, Velmurugu, and Øyvind J. Hansen. 2023. "Short and Long-Term Effects of Early and Late Weaning on Atlantic Cod, Gadus morhua" Fishes 8, no. 6: 312. https://doi.org/10.3390/fishes8060312
APA StylePuvanendran, V., & Hansen, Ø. J. (2023). Short and Long-Term Effects of Early and Late Weaning on Atlantic Cod, Gadus morhua. Fishes, 8(6), 312. https://doi.org/10.3390/fishes8060312