Effect of Host-Associated Bacillus-Supplemented Artificial Diets on Growth, Survival Rate, and Gene Expression in Early-Stage Eel Larvae (Anguilla japonica)

 , , , and
, , , and
Abstract
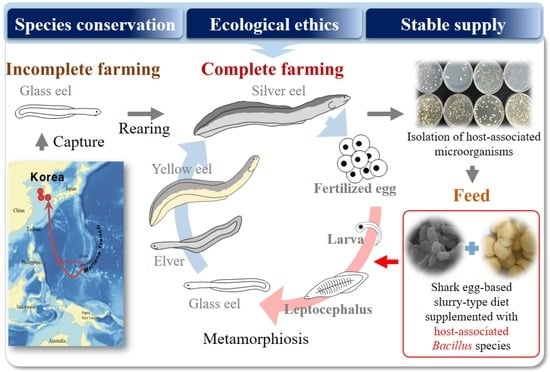
1. Introduction
2. Materials and Methods
2.1. Bacterial Isolation and Identification
2.2. Experimental Artificial Diet Preparation
2.3. Rearing System and Condition for the Feeding Trial
2.4. Growth and Survival Rate
2.5. Gene Expression Analysis
2.6. Statistical Analysis
3. Results and Discussion
3.1. Bacterial Isolation and Identification
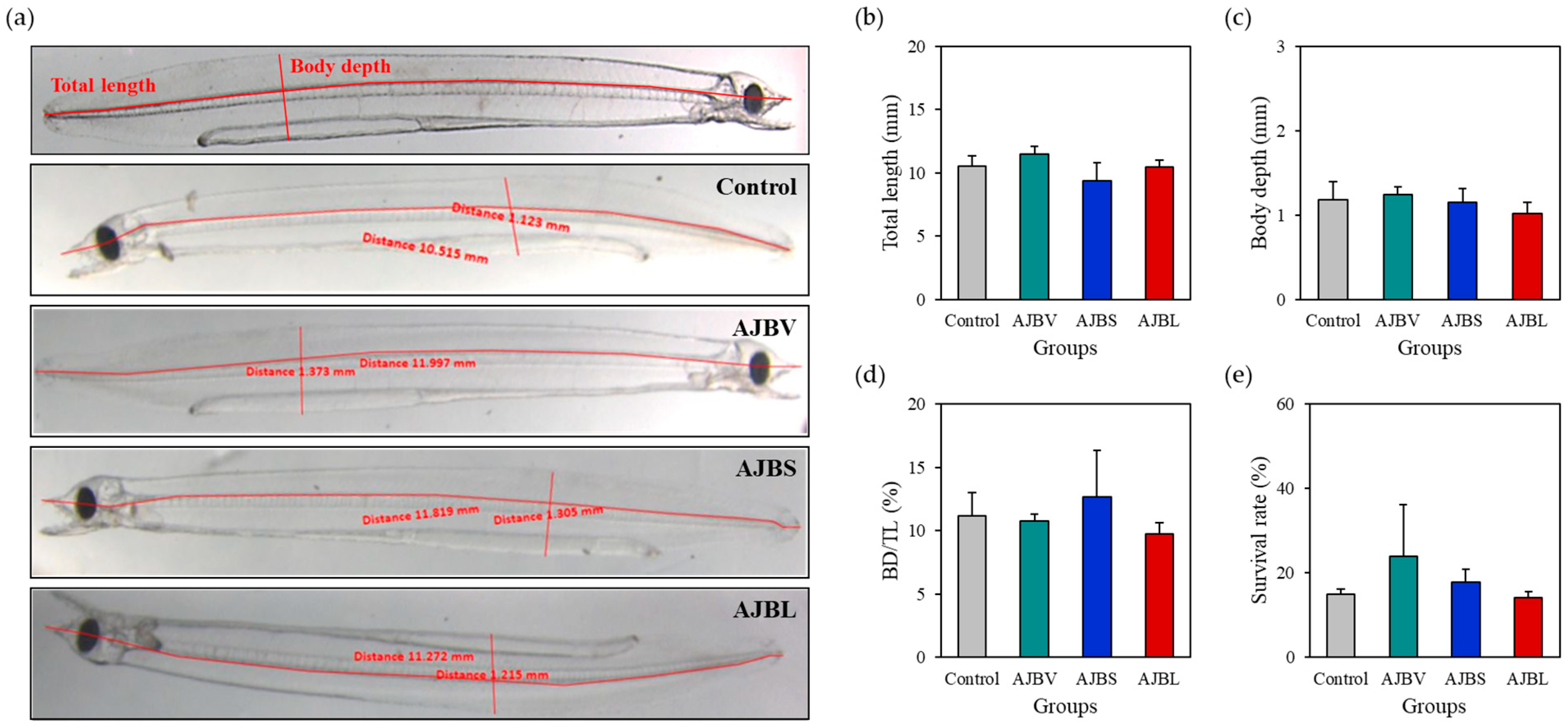
3.2. Growth and Survival Rate
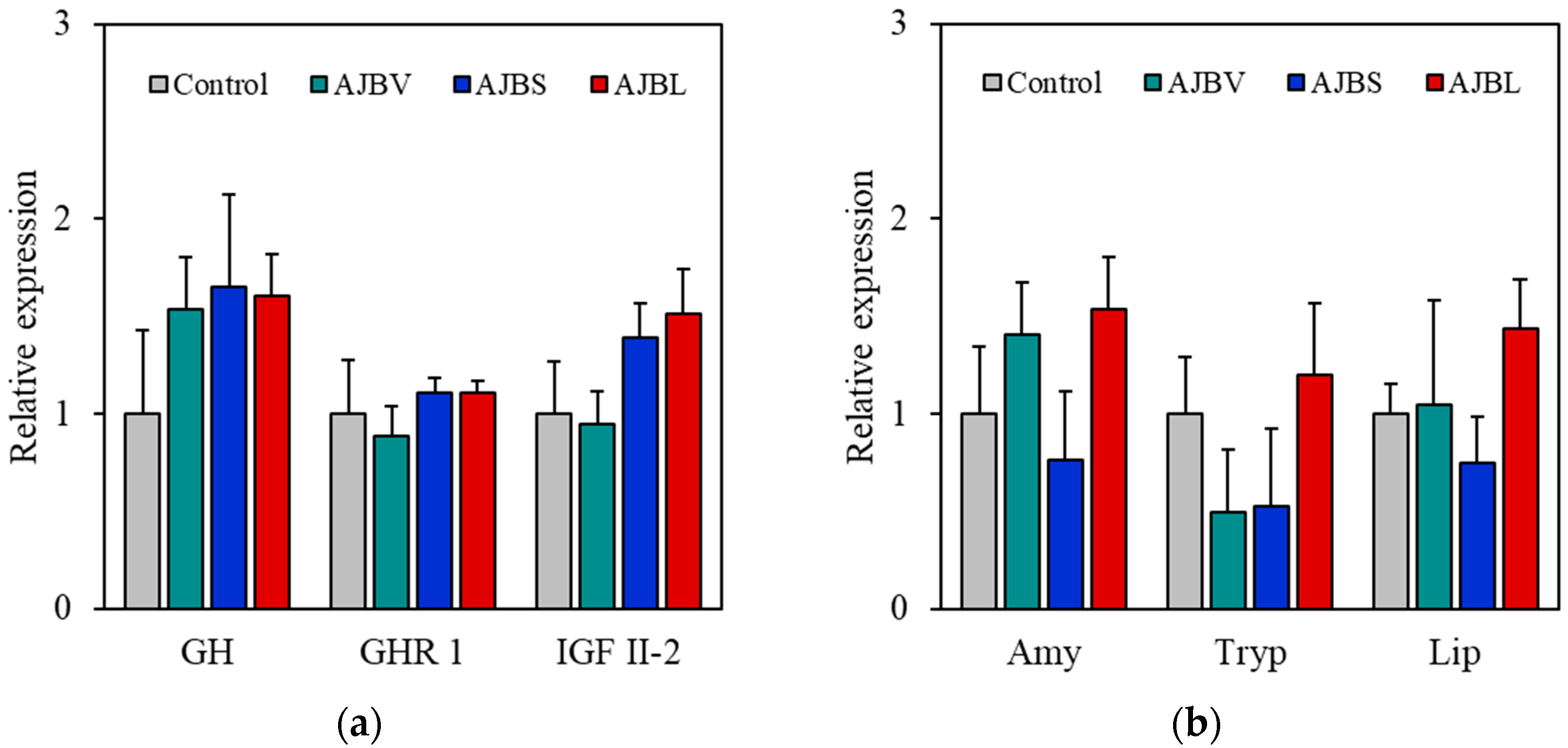
3.3. Gene Expression Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Kagawa, H.; Tanaka, H. The first success of glass eel production in the world: Basic biology on fish reproduction advances new applied technology in aquaculture. Fish Physiol. Biochem. 2005, 31, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, T.; Kurokawa, T. Influence of salinity on morphological deformities in cultured larvae of Japanese eel, Anguilla japonica, at completion of yolk resorption. Aquaculture 2009, 293, 113–118. [Google Scholar] [CrossRef]
- Jang, W.J.; Kim, S.K. Effect of Bacillus sp. supplementation diet on survival rate and microbiota composition in artificially produced eel larvae (Anguilla japonica). Front. Microbiol. 2022, 13, 891070. [Google Scholar] [CrossRef]
- Kurokawa, T.; Okamoto, T. Influence of water temperature on morphological deformities in cultured larvae of Japanese eel, Anguilla japonica, at completion of yolk resorption. J. World Aquac. Soc. 2008, 39, 726–735. [Google Scholar] [CrossRef]
- Harrison, A.J.; Walker, A.M. A review of glass eel migratory behaviour, sampling techniques and abundance estimates in estuaries: Implications for assessing recruitment, local production and exploitation. Rev. Fish Biol. Fish. 2014, 24, 967–983. [Google Scholar] [CrossRef]
- Hibiya, T. Success in collecting fully matured eel eggs. Aquaculture 1970, 3, 12–15. [Google Scholar]
- Yamamoto, K.; Yamauchi, K. Sexual maturation of Japanese eel and production of eel larvae in the aquarium. Nature 1974, 251, 220–222. [Google Scholar] [CrossRef]
- Shin, M.G.; Kim, S.K. Histological development of the digestive system in artificially produced Anguilla japonica larvae. Korean J. Fish. Aquat. Sci. 2021, 54, 298–310. [Google Scholar] [CrossRef]
- Tsukamoto, K. Aquaculture production of glass eels as a possible conservation measure for freshwater eels. In Proceedings of the 144th Annual Meeting of the American Fisheries Society 2014, Quebec City, QC, Canada, 17–21 August 2014. [Google Scholar]
- Hasan, M.T.; Jang, W.J. Effects of immunostimulants, prebiotics, probiotics, synbiotics, and potentially immunoreactive feed additives on olive flounder (Paralichthys olivaceus) aquaculture: A review. Rev. Fish. Sci. Aquac. 2019, 27, 417–437. [Google Scholar] [CrossRef]
- Mladineo, I.; Bušelić, I. Autochthonous bacterial isolates successfully stimulate in vitro peripheral blood leukocytes of the European sea bass (Dicentrarchus labrax). Front. Microbiol. 2016, 7, 1244. [Google Scholar] [CrossRef]
- Hoseinifar, S.H.; Sun, Y.Z. Probiotics as means of diseases control in aquaculture, a review of current knowledge and future perspectives. Front. Microbiol. 2018, 9, 2429. [Google Scholar] [CrossRef] [PubMed]
- Jang, W.J.; Lee, S.J. Characterization of a Bacillus sp. KRF-7 isolated from the intestine of rockfish and effects of dietary supplementation with mannan oligosaccharide in rockfish aquaculture. Fish Shellfish Immunol. 2021, 119, 182–192. [Google Scholar] [CrossRef] [PubMed]
- Van Doan, H.; Hoseinifar, S.H. Host-associated probiotics: A key factor in sustainable aquaculture. Rev. Fish. Sci. Aquac. 2020, 28, 16–42. [Google Scholar] [CrossRef]
- Kuebutornye, F.K.; Abarike, E.D. A review on the application of Bacillus as probiotics in aquaculture. Fish Shellfish Immunol. 2019, 87, 820–828. [Google Scholar] [CrossRef] [PubMed]
- Jang, W.J.; Lee, J.M. Effects of probiotic supplementation of a plant-based protein diet on intestinal microbial diversity, digestive enzyme activity, intestinal structure, and immunity in olive flounder (Paralichthys olivaceus). Fish Shellfish Immunol. 2019, 92, 719–727. [Google Scholar] [CrossRef]
- Liu, H.; Wang, S. Dietary administration of Bacillus subtilis HAINUP40 enhances growth, digestive enzyme activities, innate immune responses and disease resistance of tilapia, Oreochromis niloticus. Fish Shellfish Immunol. 2017, 60, 326–333. [Google Scholar] [CrossRef]
- Cha, J.H.; Rahimnejad, S. Evaluations of Bacillus spp. as dietary additives on growth performance, innate immunity and disease resistance of olive flounder (Paralichthys olivaceus) against Streptococcus iniae and as water additives. Aquaculture 2013, 402, 50–57. [Google Scholar] [CrossRef]
- Cerezuela, R.; Guardiola, F.A. Increases in immune parameters by inulin and Bacillus subtilis dietary administration to gilthead seabream (Sparus aurata L.) did not correlate with disease resistance to Photobacterium damselae. Fish Shellfish Immunol. 2012, 32, 1032–1040. [Google Scholar] [CrossRef]
- Hasan, M.T.; Jang, W.J. Synergistic effects of dietary Bacillus sp. SJ-10 plus β-glucooligosaccharides as a synbiotic on growth performance, innate immunity and streptococcosis resistance in olive flounder (Paralichthys olivaceus). Fish Shellfish Immunol. 2018, 82, 544–553. [Google Scholar] [CrossRef]
- Jang, W.J.; Hasan, M.T. Comparison of spore or vegetative Bacillus sp. supplementation on physiological changes and gut microbiota of the olive flounder (Paralichthys olivaceus). Aquaculture 2021, 535, 736355. [Google Scholar] [CrossRef]
- Jang, W.J.; Lee, K.B. Characteristics and biological control functions of Bacillus sp. PM8313 as a host-associated probiotic in red sea bream (Pagrus major) aquaculture. Anim. Nutr. 2023, 12, 20–31. [Google Scholar] [CrossRef] [PubMed]
- Jang, W.J.; Jeon, M.H. Dietary supplementation of Bacillus sp. PM8313 with β-glucan modulates the intestinal microbiota of red sea bream (Pagrus major) to increase growth, immunity, and disease resistance. Front. Immunol. 2022, 13, 960554. [Google Scholar] [CrossRef] [PubMed]
- Won, S.; Hamidoghli, A. Effects of Bacillus subtilis WB60 and Lactococcus lactis on growth, immune responses, histology and gene expression in Nile tilapia, Oreochromis niloticus. Microorganisms 2020, 8, 67. [Google Scholar] [CrossRef] [PubMed]
- Won, S.; Hamidoghli, A. Evaluation of potential probiotics Bacillus subtilis WB60, Pediococcus pentosaceus, and Lactococcus lactis on growth performance, immune response, gut histology and immune-related genes in whiteleg shrimp, Litopenaeus vannamei. Microorganisms 2020, 8, 281. [Google Scholar] [CrossRef]
- Olmos, J.; López, L.M. Bacillus subtilis effects on growth performance and health status of Totoaba macdonaldi fed with high levels of soy protein concentrate. Animals 2022, 12, 3422. [Google Scholar] [CrossRef]
- Ringø, E.; Harikrishnan, R.; Soltani, M.; Ghosh, K. The effect of gut microbiota and probiotics on metabolism in fish and shrimp. Animals 2022, 12, 3016. [Google Scholar] [CrossRef]
- Kim, S.K.; Lee, B.I. Development of the slurry type diet for the growing leptocephalus, eel larvae (Anguilla japonica). J. Fish. Mar. Sci. Edu. 2014, 26, 1209–1216. [Google Scholar]
- Kurokawa, T.; Koshio, M. Distribution of pepsinogen- and ghrelin-producing cells in the digestive tract of Japanese eel (Anguilla japonica) during metamorphosis and the adult stage. Gen. Comp. Endocrinol. 2011, 173, 475–482. [Google Scholar] [CrossRef]
- Murashita, K.; Furuita, H. Partial characterization and ontogenetic development of pancreatic digestive enzymes in Japanese eel Anguilla japonica larvae. Fish Physiol. Biochem. 2013, 39, 895–905. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient | Content |
|---|---|
| Shark egg | 50 g |
| Krill meal | 6 g |
| Soybean peptide | 3 g |
| Fishmeal | 3 g |
| Vitamin Mix | 0.3 g |
| Host-associated Bacillus | 1 × 1011 CFU/mL |
| Gene | Sense | Sequence (5′-3′) | Size (bp) | Access No. |
|---|---|---|---|---|
| β-actin | F | TGT GGA TCA GCA AGC AGG AG | 110 | GU001950.1 |
| R | CAG TTT TGA GTC GGC GTG TG | |||
| Growth hormone | F | TGC ACA AAG TGG AGA CCT ACC | 118 | M24066.1 |
| R | TTA ACA CAG GAC CGA AGC CC | |||
| Growth hormone receptor 1 | F | TCG CTG TTG ACA ACT TTG CG | 147 | AB180476.1 |
| R | ACA GGC AAG GGG TGA AGA TG | |||
| Insulin-like growth factors II-2 | F | ACC TGC GTA AGG ACA GCA AA | 157 | AB353117.1 |
| R | CTG CTGGTG GGT CTG CTA AA | |||
| Amylase | F | GTG GAG AAC CCA TTA CGG CA | 154 | AB070721.1 |
| R | CCA TCC TTC ACC CCA GGT TC | |||
| Trypsin | F | CCG AGC TTC CAA GGT TCT CC | 137 | AB519643.1 |
| R | GGG TTC ATA GTG TTG CCC CA | |||
| Lipase | F | GGG TTT GCT GGG AGC TAC TT | 126 | EU715406.1 |
| R | GCG TCC ACA AAT ATG GCG TC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jang, W.J.; Kim, S.-K.; Park, S.Y.; Kim, D.P.; Heo, Y.-J.; Kim, H.; Lee, S.-J.; Shin, M.G.; Lee, E.-W.; Lee, S.; et al. Effect of Host-Associated Bacillus-Supplemented Artificial Diets on Growth, Survival Rate, and Gene Expression in Early-Stage Eel Larvae (Anguilla japonica). Fishes 2023, 8, 247. https://doi.org/10.3390/fishes8050247
Jang WJ, Kim S-K, Park SY, Kim DP, Heo Y-J, Kim H, Lee S-J, Shin MG, Lee E-W, Lee S, et al. Effect of Host-Associated Bacillus-Supplemented Artificial Diets on Growth, Survival Rate, and Gene Expression in Early-Stage Eel Larvae (Anguilla japonica). Fishes. 2023; 8(5):247. https://doi.org/10.3390/fishes8050247
Chicago/Turabian StyleJang, Won Je, Shin-Kwon Kim, So Young Park, Dong Pil Kim, Yun-Jy Heo, Haham Kim, Su-Jeong Lee, Min Gyu Shin, Eun-Woo Lee, Seunghyung Lee, and et al. 2023. "Effect of Host-Associated Bacillus-Supplemented Artificial Diets on Growth, Survival Rate, and Gene Expression in Early-Stage Eel Larvae (Anguilla japonica)" Fishes 8, no. 5: 247. https://doi.org/10.3390/fishes8050247
APA StyleJang, W. J., Kim, S.-K., Park, S. Y., Kim, D. P., Heo, Y.-J., Kim, H., Lee, S.-J., Shin, M. G., Lee, E.-W., Lee, S., & Lee, J. M. (2023). Effect of Host-Associated Bacillus-Supplemented Artificial Diets on Growth, Survival Rate, and Gene Expression in Early-Stage Eel Larvae (Anguilla japonica). Fishes, 8(5), 247. https://doi.org/10.3390/fishes8050247