
Reproductive Ecology of the Chilean Round Ray (Urotrygon chilensis, Günther, 1872) in the Southern Gulf of California



Abstract
1. Introduction
2. Materials and Methods
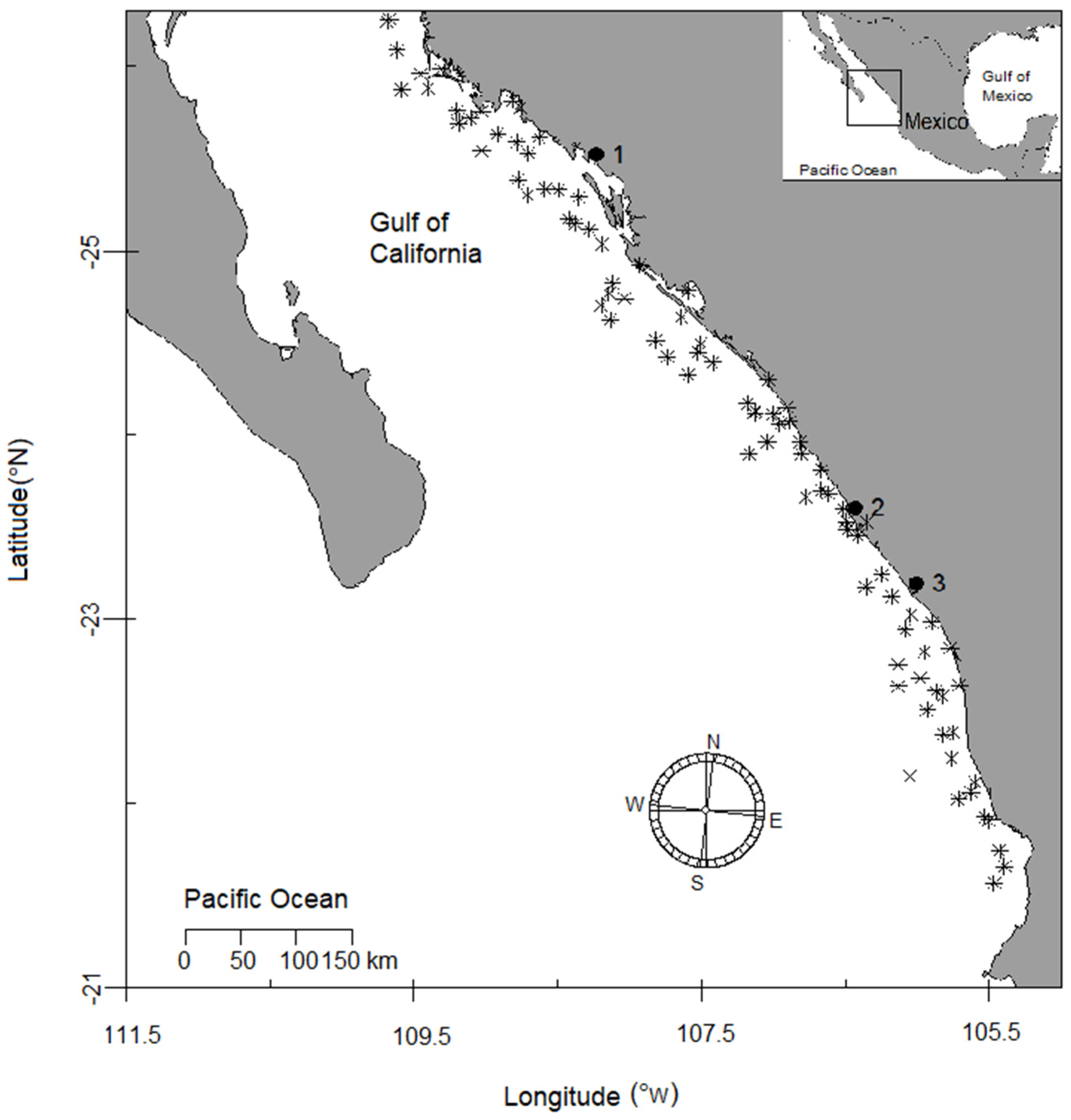
2.1. Studied Area and Sampling Procedure
2.2. Length Analysis
2.3. Reproduction
2.3.1. Reproductive Cycle
2.3.2. Size at Maturity
2.3.3. Length at Birth, Gestation Period, Fecundity, and Embryos Sex Proportion
3. Results
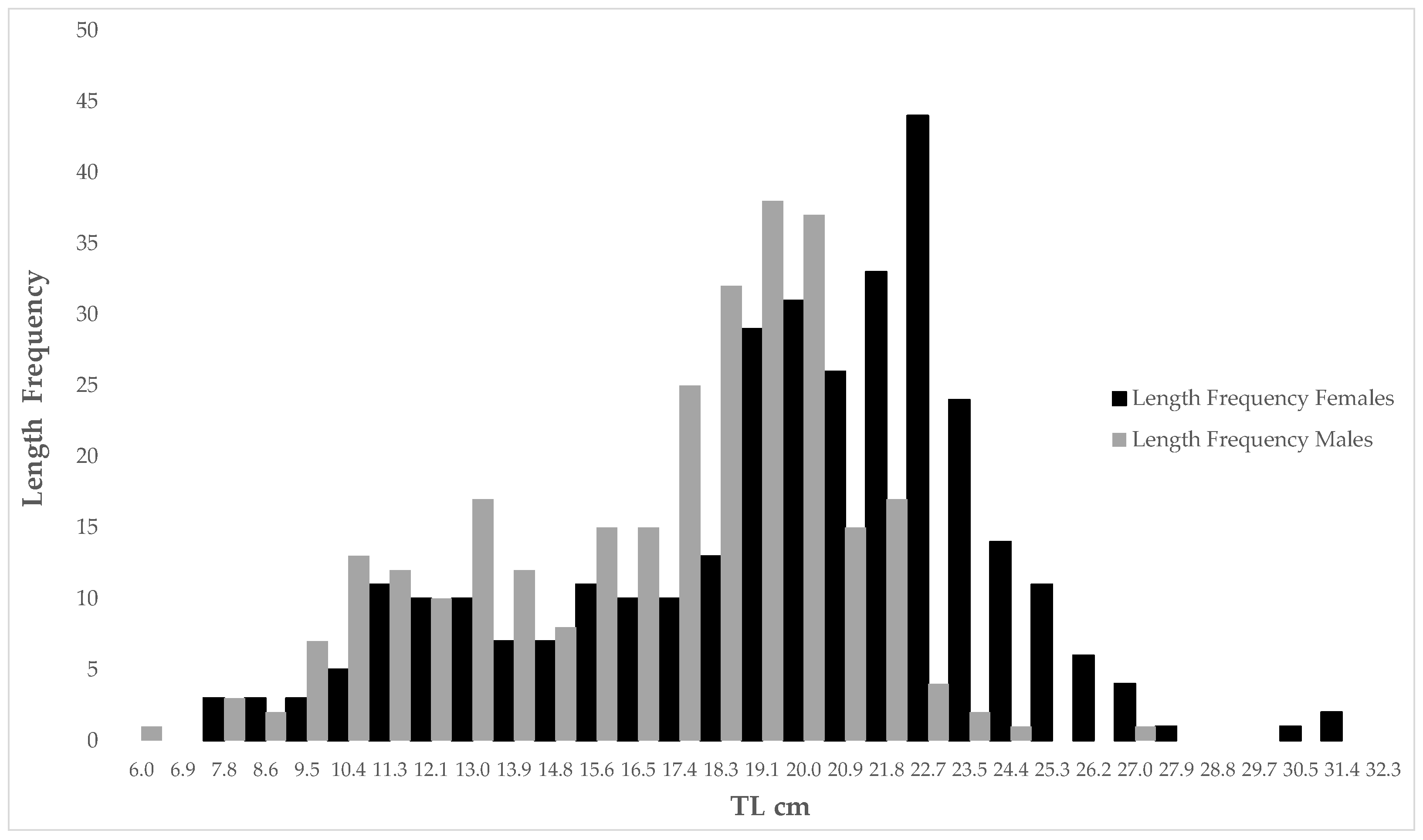
3.1. Length Analysis
3.2. Reproduction
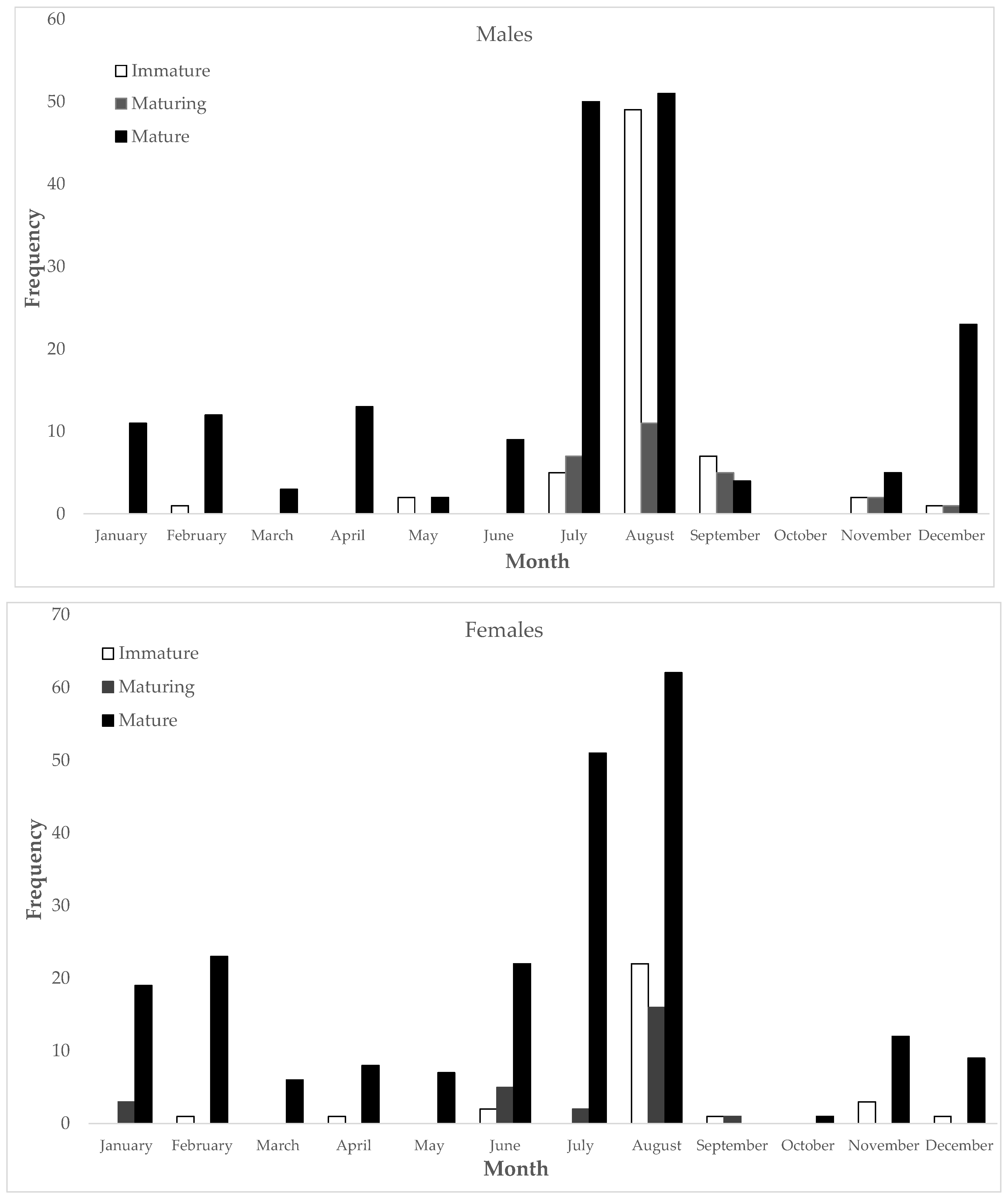
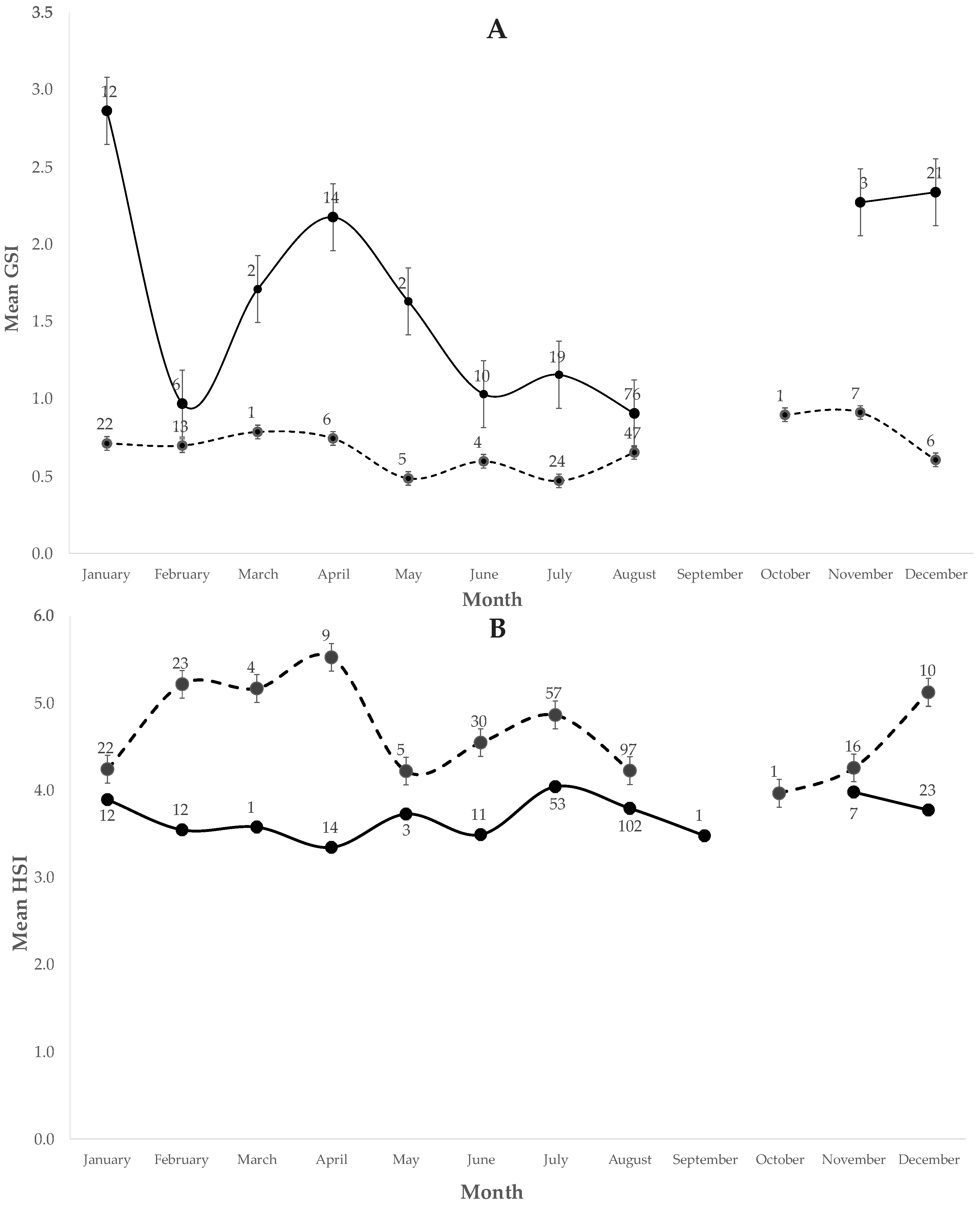
3.2.1. Reproductive Cycle
GSI and HSI
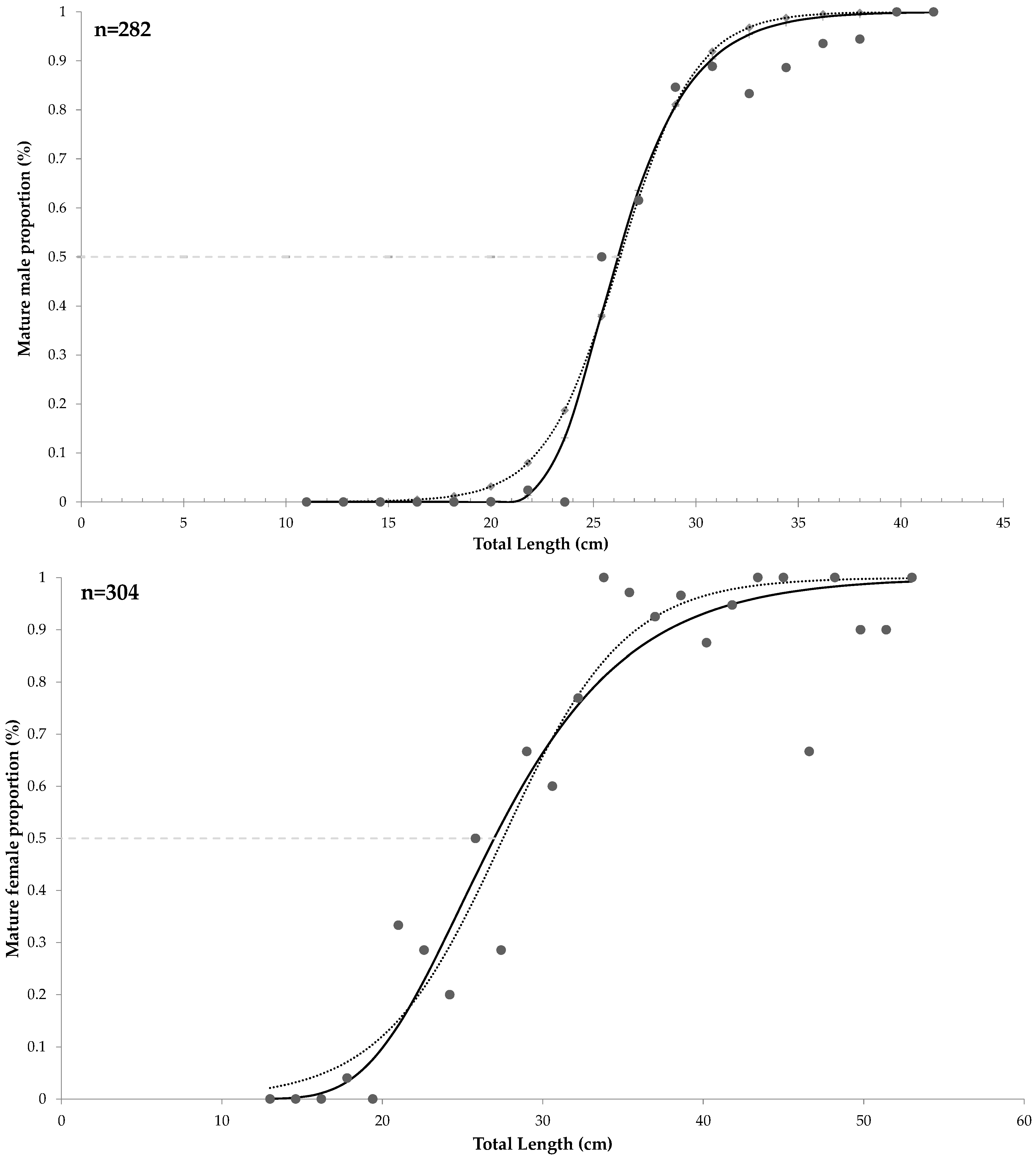
3.2.2. Size at Maturity
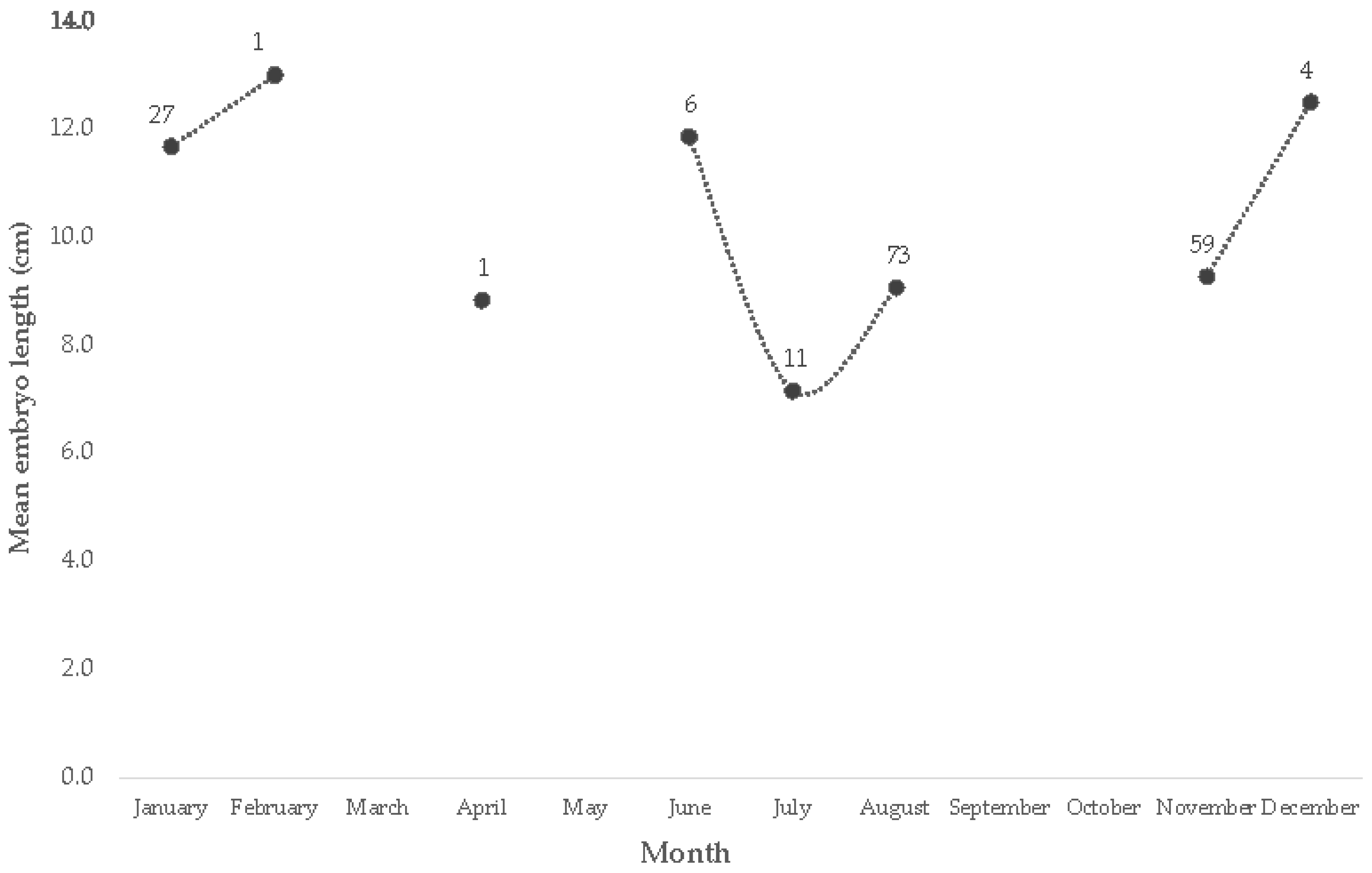
3.2.3. Length at Birth, Gestation Period, Fecundity, and Embryo Sex Proportion
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dulvy, N.K.; Fowler, S.L.; Musick, J.A.; Cavanagh, R.D.; Kyne, P.M.; Harrison, L.R.; Carlson, J.K.; Davidson, L.N.K.; Fordham, S.V.; Francis, M.P.; et al. Extinction Risk and Conservation of the World’s Sharks and Rays. eLife 2014, 3, e00590. [Google Scholar] [CrossRef]
- Consales, G.; Marsili, L. Assessment of the conservation status of Chondrichthyans: Underestimation of the pollution threat. Eur. Zool. J. 2021, 88, 165–180. [Google Scholar] [CrossRef]
- Márquez-Farías, J.F.; Blanco-Parra, M.D. Sustentabilidad y Pesca Responsable en México, Evaluación y Manejo; INAPESCA, SAGARPA: México City, México, 2006; ISBN 958800684X. [Google Scholar]
- Amezcua, F.; Madrid-Vera, J.; Aguirre-Villaseñor, H. Effect of the artisanal shrimp fishery on the ichthyofauna in the coastal lagoon of Santa María la Reforma, southeastern Gulf of California [Efecto de la pesca artesanal de camarón sobre la ictiofauna en el sistema lagunar de Santa María la Reforma, suroe. Cienc. Mar. 2006, 32, 97–109. Available online: https://www.scopus.com/inward/record.uri?eid=2-s2.0-33645522240&partnerID=40&md5=ffbb5f8324f63356834390997c948c1e (accessed on 16 February 2023). [CrossRef]
- Amezcua, F.; Ramirez, M.; Flores-Verdugo, F. Classification and Comparison of Five Estuaries in the Southeast Gulf of California Based on Environmental Variables and Fish Assemblages. Bull. Mar. Sci. 2019, 95, 139–159. [Google Scholar] [CrossRef]
- Walker, T.I. Reproduction in Fisheries Science. In Reproductive Biology and Phylogeny of Chondrichthyes: Sharks, Batoids and Chimaeras; Science Publishers: Enfield, CT, USA, 2005; pp. 81–127. [Google Scholar]
- Walker, T.I. Reproduction of Chondrichthyans. In Reproduction in Aquatic Animals; Springer: Singapore, 2020; pp. 193–223. [Google Scholar]
- Conrath, C.L. Reproductive Biology. In Management Techniques for Elasmobranch Fisheries FAO; Food & Agriculture Org.: Roma, Italy, 2005; Volume 1, p. 491. ISBN 9780124409057. [Google Scholar]
- García-Rodríguez, A.; Hernández-Herrera, A.; Galván-Magaña, F.; Ceballos-Vázquez, B.P.; Pelamatti, T.; Tovar-Ávila, J. Estimation of the Size at Sexual Maturity of the Bat Ray (Myliobatis californica) in Northwestern Mexico through a Multi-Model Inference. Fish. Res. 2020, 231, 105712. [Google Scholar] [CrossRef]
- Berkeley, S.A.; Chapman, C.; Sogard, S.M.; Ecology, S.; May, N. Maternal Age as a Determinant of Larval Growth and Survival in a Marine Fish, Sebastes melanops Maternal Age as a Determinant of Larval Growth and Survival in a Marine Fish, Sebastes melanops. Ecology 2012, 85, 1258–1264. [Google Scholar] [CrossRef]
- Birkeland, C.; Dayton, P.K. The importance in fishery management of leaving the big ones. Trends Ecol. Evol. 2005, 20, 356–358. [Google Scholar] [CrossRef] [PubMed]
- Conover, D.O.; Munch, S.B. Sustaining fisheries yields over evolutionary time scales. Science 2002, 297, 94–96. [Google Scholar] [CrossRef]
- Hsieh, C.H.; Reiss, C.S.; Hunter, J.R.; Beddington, J.R.; May, R.M.; Sugihara, G. Fishing elevates variability in the abundance of exploited species. Nature 2006, 443, 859–862. [Google Scholar] [CrossRef]
- Anderson CN, K.; Hsieh, C.; Sandin, S.A.; Hewitt, R.; Hollowed, A.; Beddington, J.; May, R.M.; Sugihara, G. Why fishing magnifies fluctuations in fish abundance. Nature 2008, 452, 835–839. [Google Scholar] [CrossRef]
- Stenseth, N.C.; Rouyer, T. Ecology: Destabilized fish stocks. Nature 2008, 452, 825–826. [Google Scholar] [CrossRef]
- Walker, T.I. Spatial and Temporal Variation in the Reproductive Biology of Gummy Shark. Mar. Freshw. Res. 2007, 58, 67–97. [Google Scholar] [CrossRef]
- Mejía-Falla, P.A.; Navia, A.F.; Cortés, E. Reproductive Variables of Urotrygon rogersi (Batoidea: Urotrygonidae): A Species with a Triannual Reproductive Cycle in the Eastern Tropical Pacific Ocean. J. Fish Biol. 2012, 80, 1246–1266. [Google Scholar] [CrossRef] [PubMed]
- Walker, T.I. Reproduction in aquatic animals. In Reproduction in Aquatic Animals: From Basic Biology to Aquaculture Technology; Springer Nature: Berlin/Heidelberg, Germany, 2020. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Model Selection and Inference: A Practical Information-Theoretic Approach; Springer: Berlin/Heidelberg, Germany, 2002; Volume 65, ISBN 0387953647. [Google Scholar]
- Chen, Y.; Jackson, D.A.; Harvey, H.H. A Comparison of Methods Fitting von Bertalanffy Growth Curve. J. Fish. Soc. Taiwan 1984, 11, 10–19. [Google Scholar]
- Hadon, M. Modelling and Quantitative Methods in Fisheries, 2nd ed.; Chapman & Hall, CRC Press: Boca Raton, FL, USA, 2011. [Google Scholar]
- Kelleher, K. Discards in the World’S Marine Fisheries; An Update; FAO Fisheries Technical Paper; Food and Agriculture Organization of the United Nations: Rome, Italy, 2005; Volume 470. [Google Scholar]
- Stobutzki, I.; Miller, M.; Jones, P.; Salini, J. Bycatch diversity and variation in a tropical Australian penaeid fishery; the implications for monitoring. Fish. Res. 2001, 53, 283–301. [Google Scholar] [CrossRef]
- Acal, D.E.; Arias, A. Evaluation of the Demerso-Pelagic Resources Available to Bottom Trawl Nets in the South of Pacific Mexico. Cienc. Mar. 1990, 16, 93–129. [Google Scholar] [CrossRef]
- D’Iglio, C.; Albano, M.; Famulari, S.; Spanò, N.; Rinelli, P.; Savoca, S.; Capillo, G. Basic Intersexuality (Abnormal Hermaphroditism) in the Blackmouth Catshark, Galeus melastomus, (Rafinesque, 1810), from the Southern Tyrrhenian Sea (Central Mediterranean Sea). Fishes 2022, 7, 120. [Google Scholar] [CrossRef]
- Mulas, A.; Bellodi, A.; Porcu, C.; Cau, A.; Coluccia, E.; Riccardo, D.; Marongiu, M.; Pesci, P.; Follesa, M. Living naked: First case of lacking of skin related structures in an Elasmobranch, the blackmouth catshark (Galeus melastomus). J. Fish Biol. 2020, 97, 1252–1256. [Google Scholar] [CrossRef] [PubMed]
- Amezcua-Linares, F.; Cruz-González, K.A.; Flores-de-Santiago, F.; Muro, V.; Green, L.; Amezcua, F. Are the Temporal Changes Observed in the Reproductive Biology of the Estuarine Conguito Sea Catfish Related to Increased Small-Scale Fishing Effort on the Northwestern Pacific Coast of Mexico? Mar. Coast. Fish. 2022, 14, e10217. [Google Scholar] [CrossRef]
- Christensen, V.; Guénette, S.; Heymans, J.J.; Walters, C.J.; Watson, R.; Zeller, D.; Pauly, D. Hundred-year decline of North Atlantic predatory fishes. Fish Fish. 2003, 4, 1–24. [Google Scholar] [CrossRef]
- Cortés, E. Life History Patterns and Correlations in Sharks. Rev. Fish. Sci. 2000, 8, 299–344. [Google Scholar] [CrossRef]
- Torres-Huerta, A.M.; López-Pérez, R.A.; Tapia-García, M.; Gracía, A. Distribution of Batoid Demersal Assemblages on the Continental Shelf of the Gulf of Tehuantepec. Mar. Freshw. Res. 2019, 70, 1445–1458. [Google Scholar] [CrossRef]
- Ordoñez-Lopez, J. Aspectos Reproductivos y Alimenticios de La Raya Pinta Urotrygon chilensis (Gunther, 1871) (Pisces—Usolophidae) En El Pacifico Central Mexicano. Bachelor’s Thesis, Universidad Nacional Autónoma de México, Mexico City, Mexico, 2004. [Google Scholar]
- Guzmán-Castellanos, A.B. Historia de Vida de la Raya Redonda Urotrygon chilensis (Günther, 1872) en el Sureste del Pácifico Mexicano. Ph.D. Thesis, Centro de Investigaciones Biológicas del Noroeste, La Paz, Mexico, 2015. [Google Scholar]
- Trinnie, F.I.; Walker, T.I.; Jones, P.L.; Laurenson, L.J. Reproductive Cycle of Urolophus Cruciatus in South-Eastern Australia: Does the Species Exhibit Obligate or Facultative Diapause? Mar. Biol. 2016, 163, 226. [Google Scholar] [CrossRef]
- Trinnie, F.I.; Walker, T.I.; Jones, P.L.; Laurenson, L.J. Asynchrony and Regional Differences in the Reproductive Cycle of the Greenback Stingaree Urolophus viridis from South-Eastern Australia. Environ. Biol. Fishes 2014, 98, 425–441. [Google Scholar] [CrossRef]
- Trinnie, F.I.; Walker, T.I.; Jones, P.L.; Laurenson, L.J. Biennial Reproductive Cycle in an Extensive Matrotrophic Viviparous Batoid: The Sandyback Stingaree Urolophus bucculentus from South-Eastern Australia. J. Fish Biol. 2012, 80, 1267–1291. [Google Scholar] [CrossRef]
- White, W.T.; Platell, M.E.; Potter, I.C. Relationship between Reproductive Biology and Age Composition and Growth in Urolophus iobatus (Batoidea: Urolophidae). Mar. Biol. 2001, 138, 135–147. [Google Scholar] [CrossRef]
- Trinnie, F.I.; Walker, T.I.; Jones, P.L.; Laurenson, L.J. Regional Differences in the Reproductive Parameters of the Sparsely-Spotted Stingaree, Urolophus paucimaculatus, from South-Eastern Australia. Mar. Freshw. Res. 2014, 65, 943–958. [Google Scholar] [CrossRef]
- Fowler, A.J.; McLeay, L.; Short, D.A. Spatial Variation in Size and Age Structures and Reproductive Characteristics of the King George Whiting (Percoidei: Sillaginidae) in South Australian Waters. Mar. Freshw. Res. 2000, 51, 11–22. [Google Scholar] [CrossRef]
- Brown-Peterson, N.J.; Overstreet, R.M.; Lotz, J.M.; Franks, J.S.; Burns, K.M. Reproductive Biology of Cobia, Rachycentron canadum, from Coastal Waters of the Southern United States. Fish. Bull. 2001, 99, 15–28. [Google Scholar]
- Tovar-Ávila, J.; Walker, T.I.; Day, R.W. Reproduction of Heterodontus portusjacksoni in Victoria, Australia: Evidence of Two Populations and Reproductive Parameters for the Eastern Population. Mar. Freshw. Res. 2007, 58, 956–965. [Google Scholar] [CrossRef]
- Yiğin, C.Ç.; İşmen, A. Reproductive Biology of Spiny Dogfish Squalus acanthias, in the North Aegean Sea. Turk. J. Fish. Aquat. Sci. 2013, 13, 447–452. [Google Scholar] [CrossRef]
- Braccini, J.M.; Bronwyn, M.G.; Walker, T.I. Determining Reproductive Parameters for Population Assessments of Chondrichthyan Species with Asynchronous Ovulation and Parturition: Piked Spurdog (Squalus megalops) as a Case Study. Mar. Freshw. Res. 2006, 57, 105–119. [Google Scholar] [CrossRef]
- Garcés-García, K.C.; Tovar-Ávila, J.; Vargas-Trejo, B.; Chávez-Arrenquín, D.A.; Walker, T.I.; Day, R.W. Elasmobranch bycatch by prawn trawls in the Gulf of California: First comprehensive analysis and the effect of fish escape devices. Fish. Res. 2020, 230, 105639. [Google Scholar] [CrossRef]
- Yokota, L.; Lessa, R. A nursery Area for Sharks and Rays in Northeastern Brazil. Environ. Biol. Fishes 2006, 75, 349–360. [Google Scholar] [CrossRef]
- Horie, T.; Tanaka, S. Reproduction and Food Habits of Two Species of Sawtail Catsharks, Galeus eastmani and G. nipponensis, in Suruga Bay, Japan. Fish. Sci. 2000, 66, 812–825. [Google Scholar] [CrossRef]
- Lombardi-Carlson, L.A.; Cortés, E.; Parsons, G.R.; Manire, C.A. Latitudinal Variation in Life-History Traits of Bonnethead Sharks, Sphyrna tiburo, (Carcharhiniformes: Sphyrnidae) from the Eastern Gulf of Mexico. Mar. Freshw. Res. 2003, 54, 875–883. [Google Scholar] [CrossRef]
- Last, P.R.; McEachran, J.D. Round rays, Family Urotrygonidae. In Rays of the World; Last, P.R., White, W.T., de Carvalho, M.R., Séret, B., Stehmann, M.F.W., Naylor, G.J.P., Eds.; CSIRO Publishing: Melbourne, Australia, 2016; pp. 656–673. [Google Scholar]
- López-Martínez, J.; Hernández-vázquez, S.; Morales-azpeitia, R. Peces de La Fauna de Acompañamiento En La Pesca Industrial de Camarón En El Golfo de California, México. Rev. Biol. Trop. 2010, 58, 925–942. [Google Scholar] [CrossRef]
- Brander, K. Disappearance of Common Skate Raia batis from Irish Sea. Nature 1981, 290, 48–49. [Google Scholar] [CrossRef]
- Jennings, S.; Kaiser, M.J. The Effects of Fishing on Marine Ecosystems. Adv. Mar. Biol. 1998, 34, 201–212. [Google Scholar] [CrossRef]
- Stevens, J.D.; Bonfil, R.; Dulvy, N.K.; Walker, P.A. The Effects of Fishing on Sharks, Rays, and Chimaeras (Chondrichthyans), and the Implications for Marine Ecosystems. ICES J. Mar. Sci. 2000, 57, 476–494. [Google Scholar] [CrossRef]
- Hanchet, S. Reproductive Biology of Squalus Acanthias from the East Coast, South Island, New Zealand. N. Z. J. Mar. Freshw. Res. 1988, 22, 537–549. [Google Scholar] [CrossRef]
- Peres, M.B.; Vooren, C.M. Sexual Development, Reproductive Cycle, and Fecundity of the School Shark, Galeorhinus galeus off Southern Brazil. Fish. Bull. 1991, 17, 352–367. [Google Scholar]
- Cervantes-Hernández, P.; Ramos-Cruz, S. Evaluación Del Estado de La Pesquería de Camarón En El Golfo de Tehuantepec. Hidrobiológica 2006, 16, 233–239. [Google Scholar]
- Shin, Y.J.; Rochet, M.J.; Jennings, S.; Field, J.G.; Gislason, H. Using Size-Based Indicators to Evaluate the Ecosystem Effects of Fishing. ICES J. Mar. Sci. 2005, 62, 384–396. [Google Scholar] [CrossRef]
- Kendall, N.W.; Hard, J.J.; Quinn, T.P. Quantifying Six Decades of Fishery Selection for Size and Age at Maturity in Sockeye Salmon. Evol. Appl. 2009, 2, 523–536. [Google Scholar] [CrossRef] [PubMed]
- Ruggerone, G.T.; Zimmermann, M.; Myers, K.W.; Nielsen, J.L.; Rogers, D.E. Competition between Asian Pink Salmon (Oncorhynchus gorbuscha) and Alaskan Sockeye Salmon (O. nerka) in the North Pacific Ocean. Fish. Oceanogr. 2003, 12, 209–219. [Google Scholar] [CrossRef]
- Marteinsdottir, G.; Begg, G.A. Essential Relationships Incorporating the Influence of Age, Size and Condition on Variables Required for Estimation of Reproductive Potential in Atlantic Cod Gadus morhua. Mar. Ecol. Prog. Ser. 2002, 235, 235–256. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Maturity Stage | Females | Males |
|---|---|---|
| (GS = 1) Ovaries are thin tissue strips with epigonal gland predominant and small white undeveloped follicles. (OGS = 1) Oviducal glands are indistinct from the anterior oviducts. (US = 1) Uteri are uniformly thin tubular structures. | (GS = 1) Testes are thin tissue strips with epigonal gland predominant. (CS = 1) Claspers are small, and pliable with no calcification. |
| (GS = 2) Ovaries are thickened strips with small white and yoking medium-size follicles. (OGI = 2) Oviducal glands are distinct but only partly formed. (US = 2) Uteri are thin tubular structures partly enlarged posteriorly. | (GS = 2) Testes are thickened strips with epigonal gland tissue extensive. (CS = 2) Claspers are enlarged but partly calcified. |
| (GS = 3) Ovaries are enlarged and thickened containing large follicles with yellowish yolk. (GOS = 3) Oviducal glands are heart shaped. (US = 3) Uteri are uniformly enlarged and widened tubular structures but no eggs or embryos are visible (Sexually inactive). (US = 4) Macroscopically visible eggs are present (sexually active). (US = 5) Embryos of any size are present (sexually active). (US = 6) No eggs or embryos but uteri walls are distended (in recovery or postpartum) (Sexually inactive). | (GS = 3) Testes are enlarged and predominant with epigonal gland tissue negligible. (CS = 3) Claspers are enlarged, rigid and fully calcified. |
| Model | TL50 (cm) | AICc | Δi | wi (%) |
|---|---|---|---|---|
| Females | ||||
| Gompertz | 24.8 | −63.4 | 0.89 | 0.38 |
| Brouwer and Griffiths † | 27.5 | −64.3 | 0 | 0.61 |
| Males | ||||
| Gompertz † | 25.3 | −65.1 | 0 | 0.96 |
| Brouwer and Griffiths | 26.3 | −58.5 | 6.63 | 0.03 |
| Month | (mm) | SD | n |
|---|---|---|---|
| January | 13 | - | 1 |
| February | 13.7 | 5.0 | 12 |
| March | 8.8 | 1.0 | 4 |
| April | 4.3 | 0.3 | 3 |
| May | - | - | 0 |
| June | 19.4 | 8.1 | 7 |
| July | 20.8 | 4.5 | 6 |
| August | 19.6 | 4.1 | 17 |
| September | - | - | 0 |
| October | 6.0 | - | 1 |
| November | 24.2 | 13.9 | 4 |
| December | 21.3 | 11.8 | 3 |
| Sex | TL cm | n | SR | SM | F | RP | BL | Region | SY | |
|---|---|---|---|---|---|---|---|---|---|---|
| Avg. | Max. | |||||||||
| ♀ | 14.5–44.5 | 100 | 0.96:1 | 25.2 | 1.7 | 4 | Spring | 14–15 | MG | 2004 |
| ♂ | 17.6–36.2 | 104 | 26.5 | |||||||
| ♀ | 10.9–39.5 | 490 | 1:1/6.01:1 | 25.3 | 5 | Spring/summer | 10–14.5 | GT | 2012 | |
| ♂ | 10.9–33.8 | 211 | - | |||||||
| ♀ | 11.4–51.6 | 320 | 1:1 | 27.5 | 2.14 | 6 | Winter/spring | 11.2–15.6 | SGC | 2022 |
| ♂ | 11.2–41.3 | 286 | 25.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alvarez-Fuentes, C.J.; Tovar-Ávila, J.; Payan-Alejo, J.; Chávez-Arrenquín, D.A.; Salgado-Ugarte, I.H.; Amezcua, F. Reproductive Ecology of the Chilean Round Ray (Urotrygon chilensis, Günther, 1872) in the Southern Gulf of California. Fishes 2023, 8, 193. https://doi.org/10.3390/fishes8040193
Alvarez-Fuentes CJ, Tovar-Ávila J, Payan-Alejo J, Chávez-Arrenquín DA, Salgado-Ugarte IH, Amezcua F. Reproductive Ecology of the Chilean Round Ray (Urotrygon chilensis, Günther, 1872) in the Southern Gulf of California. Fishes. 2023; 8(4):193. https://doi.org/10.3390/fishes8040193
Chicago/Turabian StyleAlvarez-Fuentes, Carlos J., Javier Tovar-Ávila, Jorge Payan-Alejo, Darío A. Chávez-Arrenquín, Isaias H. Salgado-Ugarte, and Felipe Amezcua. 2023. "Reproductive Ecology of the Chilean Round Ray (Urotrygon chilensis, Günther, 1872) in the Southern Gulf of California" Fishes 8, no. 4: 193. https://doi.org/10.3390/fishes8040193
APA StyleAlvarez-Fuentes, C. J., Tovar-Ávila, J., Payan-Alejo, J., Chávez-Arrenquín, D. A., Salgado-Ugarte, I. H., & Amezcua, F. (2023). Reproductive Ecology of the Chilean Round Ray (Urotrygon chilensis, Günther, 1872) in the Southern Gulf of California. Fishes, 8(4), 193. https://doi.org/10.3390/fishes8040193