
Zootechnical Brown Trout (Salmo trutta L. 1758) Ovarian Fluid Fails to Upregulate the Swimming Performances of Native Mediterranean Brown Trout (Salmo cettii Rafinesque, 1810) Sperm in the Biferno River

 , ,
, ,  , and
, and
Abstract
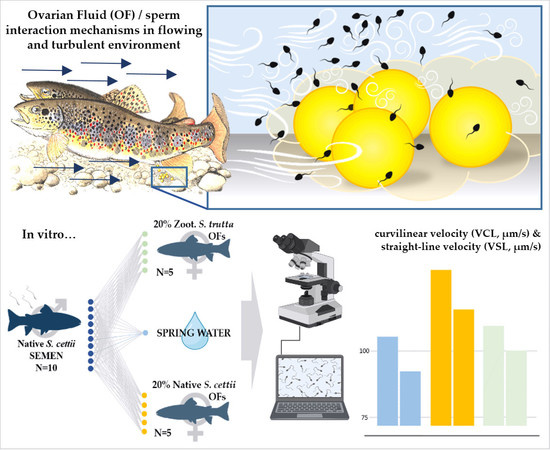
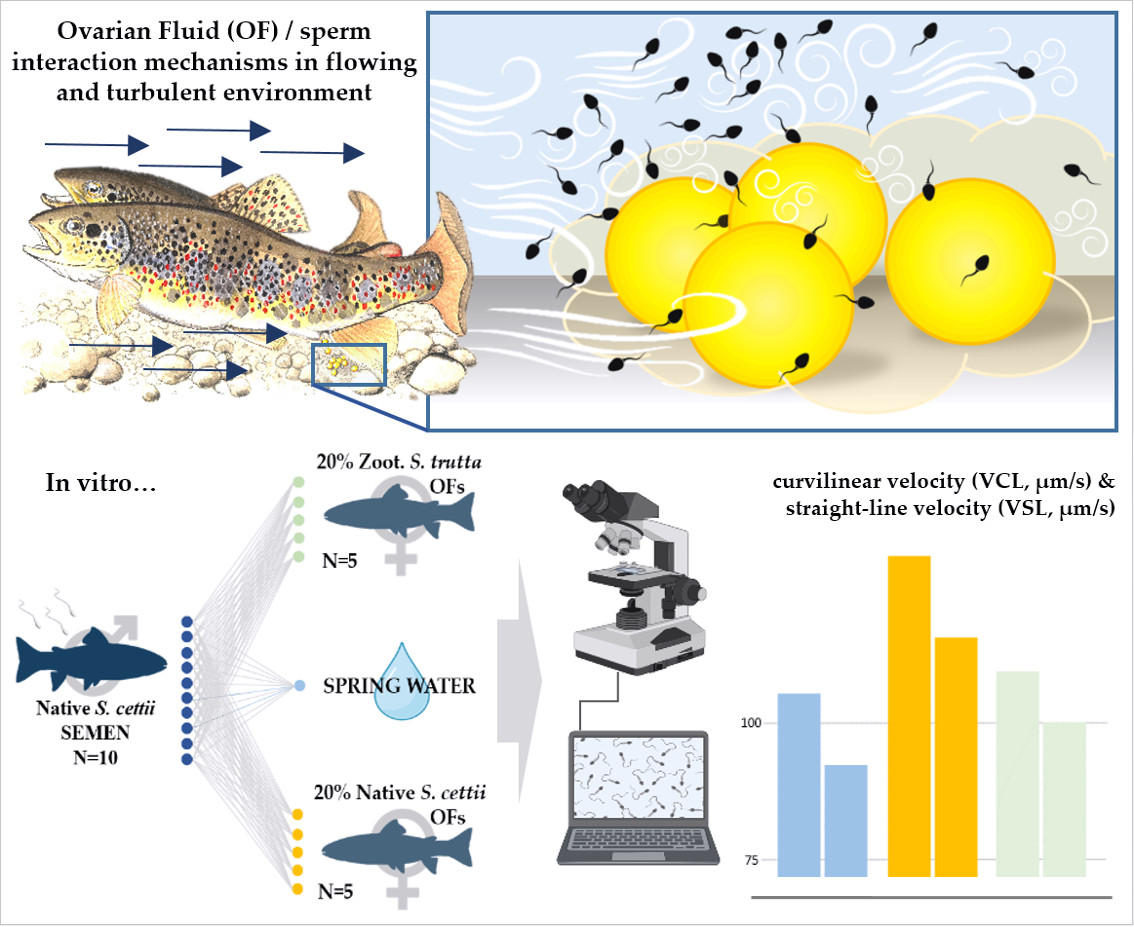
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Ovarian Fluid and Sperm Collection and Analytical Measurements
2.3. Experimental Design
2.4. Sperm Motility Analysis
2.5. Statistical Analysis
3. Results
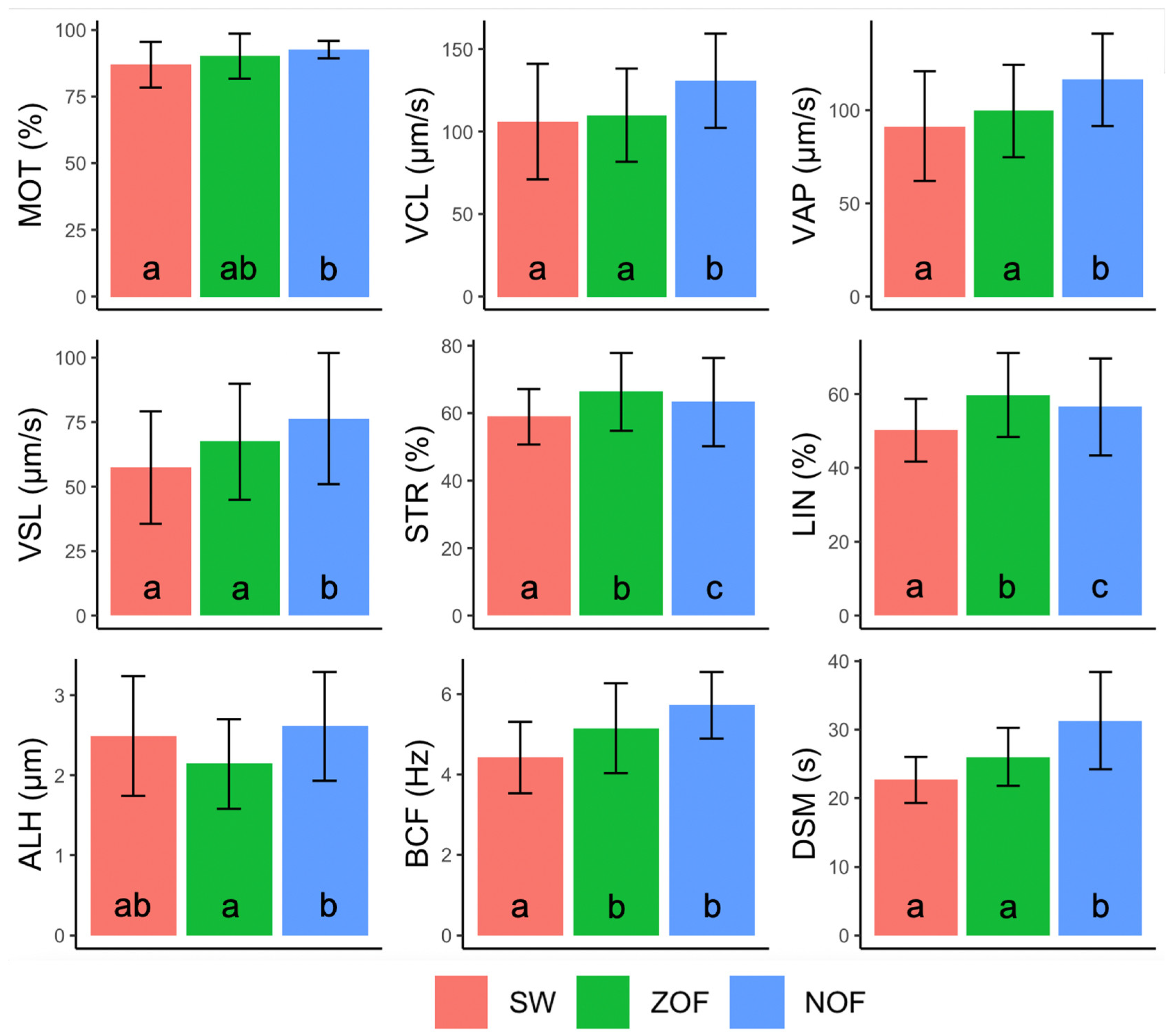
3.1. Analytical Measurements of Semen and Ovarian Fluid
3.2. Comparison between Spring Water, ZOF and NOF as Activation Media
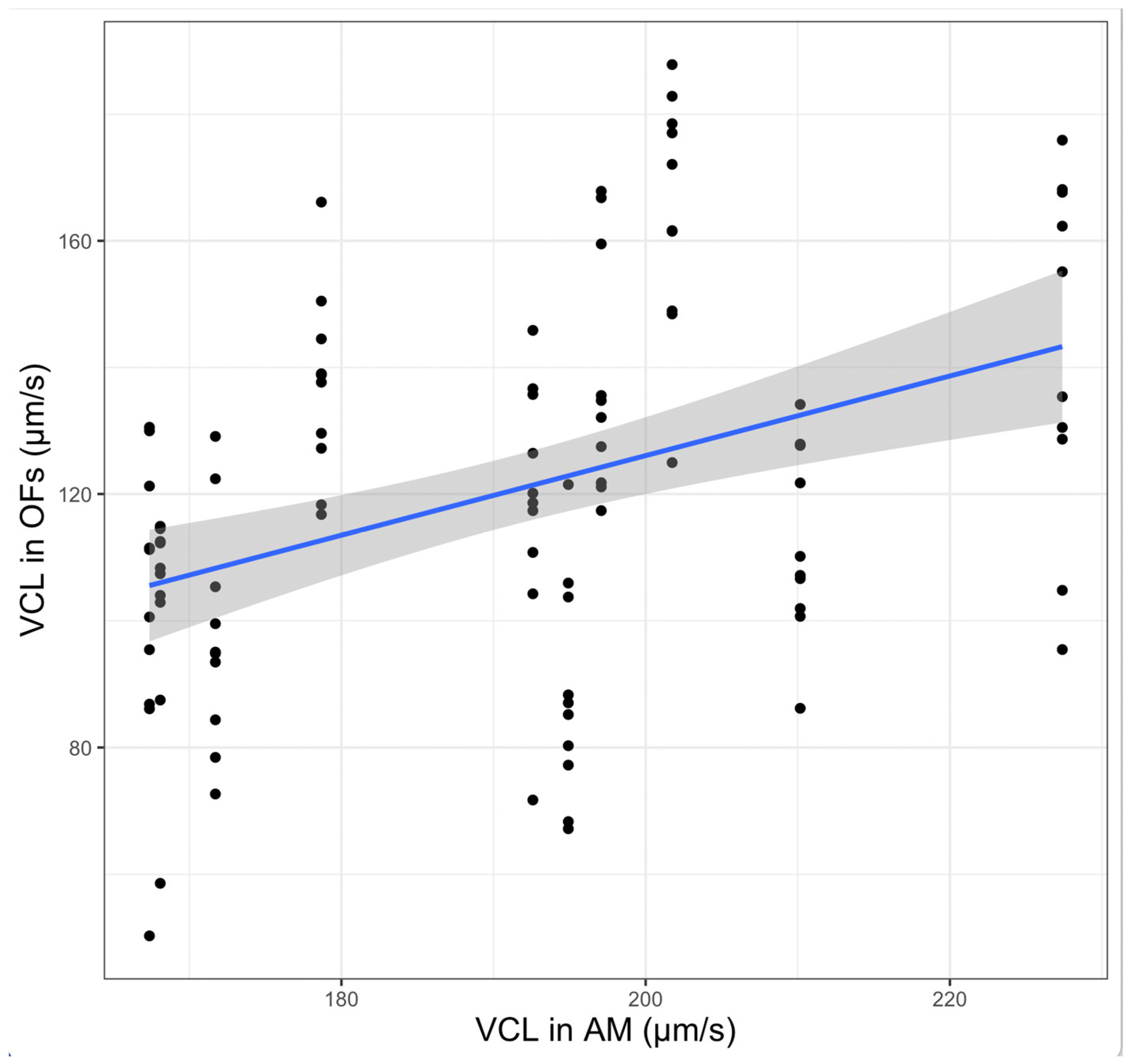
3.3. Exploring the Effects of Specific Males and Females Used in the Experiment
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding

Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Morisawa, M. Initiation mechanism of sperm motility at spawning in teleosts. Zool. Sci. 1985, 2, 605–615. [Google Scholar]
- Billard, R.; Bry, C.; Gillet, C. Stress, environment and reproduction in teleost fish. In Stress and Fish; Pickering, A.D., Ed.; Academic Press: London, UK, 1981; pp. 185–208. [Google Scholar]
- Petersen, C.W.; Warner, R.R.; Shapiro, D.Y.; Marconato, A. Components of fertilization success in the bluehead wrasse, Thalassoma bifasciatum. Behav. Ecol. 2001, 12, 237–245. [Google Scholar] [CrossRef]
- Hoysak, D.J.; Liley, N.R. Fertilization dynamics in sockeye salmon and a comparison of 450 sperm from alternative male phenotypes. J. Fish Biol. 2001, 58, 1286–1300. [Google Scholar] [CrossRef]
- Liao, W.B.; Huang, Y.; Zeng, Y.; Zhong, M.J.; Luo, Y.; Lüpold, S. Ejaculate evolution in external fertilizers: Influenced by sperm competition or sperm limitation? Evolution 2018, 72, 4–17. [Google Scholar] [CrossRef] [PubMed]
- Kholodnyy, V.; Gadêlha, H.; Cosson, J.; Boryshpolets, S. How do freshwater fish sperm find the egg? The physicochemical factors guiding the gamete encounters of externally fertilizing freshwater fish. Rev. Aquac. 2020, 12, 1165–1192. [Google Scholar] [CrossRef]
- Lahnsteiner, F. The influence of ovarian fluid on the gamete physiology in the Salmonidae. Fish Physiol. Biochem. 2002, 27, 49–59. [Google Scholar] [CrossRef]
- Elofsson, H.; Van Look, K.J.W.; Sundell, K.; Sundh, H.; Borg, B. Stickleback sperm saved by salt in ovarian fluid. J. Exp. Biol. 2006, 209, 4230–4237. [Google Scholar] [CrossRef]
- Lehnert, S.J.; Butts, I.A.E.; Flannery, E.W.; Peters, K.M.; Heath, D.D.; Pitcher, T.E. Effects of ovarian fluid and genetic differences on sperm performance and fertilization success of alternative reproductive tactics in Chinook salmon. J. Evol. Biol. 2017, 30, 1236–1245. [Google Scholar] [CrossRef]
- Rosengrave, P.; Taylor, H.; Montgomerie, R.; Metcalf, V.; McBride, K.; Gemmell, N.J. Chemical composition of seminal and ovarian fluids of chinook salmon (Oncorhynchus tshawytscha) and their effects on sperm motility traits. Comp. Biochem. Physiol. 2009, 152, 123–129. [Google Scholar] [CrossRef]
- Turner, E.; Montgomerie, R. Ovarian fluid enhances sperm movement in Arctic charr. J. Fish Biol. 2002, 60, 1570–1579. [Google Scholar] [CrossRef]
- Urbach, D.; Folstad, I.; Rudolfsen, G. Effects of ovarian fluid on sperm velocity in Arctic charr (Salvelinus alpinus). Behav. Ecol. Sociobiol. 2005, 57, 438–444. [Google Scholar] [CrossRef]
- Dietrich, G.J.; Wojtczak, M.; Słowińska, M.; Dobosz, S.; Kuźmiński, H.; Ciereszko, A. Effects of ovarian fluid on motility characteristics of rainbow trout (Oncorhynchus mykiss Walbaum) spermatozoa. J. Appl. Ichthyol. 2008, 24, 503–507. [Google Scholar] [CrossRef]
- Diogo, P.; Soares, F.; Dinis, M.T.; Cabrita, E. The influence of ovarian fluid on Solea senegalensis sperm motility. J. Appl. Ichthyol. 2010, 26, 690–695. [Google Scholar] [CrossRef]
- Galvano, P.M.; Johnson, K.; Wilson, C.C.; Pitcher, T.E.; Butts, I.A. Ovarian fluid influences sperm performance in lake trout, Salvelinus namaycush. Reprod. Biol. 2013, 13, 172–175. [Google Scholar] [CrossRef]
- Butts, I.A.E.; Johnson, K.; Wilson, C.C.; Pitcher, T.E. Ovarian fluid enhances sperm velocity based on relatedness in lake trout, Salvelinus namaycush. Theriogenology 2012, 78, 2105–2109. [Google Scholar] [CrossRef]
- Butts, I.A.E.; Prokopchuk, G.; Kašpar, V.; Cosson, J.; Pitcher, T.E. Ovarian fluid impacts flagellar beating and biomechanical metrics of sperm between alternative reproductive tactics. J. Exp. Biol. 2017, 220, 2210–2217. [Google Scholar] [CrossRef]
- Poli, F.; Immler, S.; Gasparini, C. Effects of ovarian fluid on sperm traits and its implications for cryptic female choice in zebrafish. Behav. Ecol. 2019, 30, 1298–1305. [Google Scholar] [CrossRef]
- Rosengrave, P.; Gemmell, N.J.; Metcalf, V.; McBride, K.; Montgomerie, R. A mechanism for cryptic female choice in chinook salmon. Behav. Ecol. 2008, 19, 1178–1185. [Google Scholar] [CrossRef]
- Beirão, J.; Purchase, C.F.; Wringe, B.F.; Fleming, I.A. Wild Atlantic cod sperm motility is negatively affected by ovarian fluid of farmed females. Aquac. Environ. Interact. 2014, 5, 61–70. [Google Scholar] [CrossRef]
- Rosengrave, P.; Montgomerie, R.; Gemmell, N. Cryptic female choice enhances fertilization success and embryo survival in chinook salmon. Proc. R. Soc. B 2016, 283, 20160001. [Google Scholar] [CrossRef]
- Cherr, G.N.; Morisawa, M.; Vines, C.A.; Yoshida, K.; Smith, E.H.; Matsubara, T.; Pillai, M.C.; Griffin, F.J.; Yanagimachi, R. Two egg-derived molecules in sperm motility initiation and fertilization in the Pacific herring (Clupea pallasi). Int. J. Dev. Biol. 2008, 52, 743–752. [Google Scholar] [CrossRef] [PubMed]
- Hatef, A.; Niksirat, H.; Alavi, S.M. Composition of ovarian fluid in endangered Caspian brown trout, Salmo trutta caspius, and its effects on spermatozoa motility and fertilizing ability compared to freshwater and a saline medium. Fish Physiol. Biochem. 2009, 35, 695–700. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.D.; Niu, H.X.; Meng, Z.; Liu, X.F.; Lei, J.L. Biochemical composition of the ovarian fluid and its effects on the fertilization capacity of turbot Scophthalmus maximus during the spawning season. J. Fish Biol. 2015, 86, 1612–1620. [Google Scholar] [CrossRef] [PubMed]
- Alonzo, S.H.; Stiver, K.A.; Marsh-Rollo, S.E. Ovarian fluid allows directional cryptic female choice despite external fertilization. Nat. Commun. 2016, 16, 12452. [Google Scholar] [CrossRef]
- Yeates, S.E.; Diamond, S.E.; Einum, S.; Emerson, B.C.; Holt, W.V.; Gage, M.J. Cryptic choice of conspecific sperm controlled by the impact of ovarian fluid on sperm swimming. Behav. Evol. 2013, 67, 3523–3536. [Google Scholar] [CrossRef]
- Beirão, J.; Purchase, C.F.; Wringe, B.F.; Fleming, I.A. Inter-population ovarian fluid variation differentially modulates sperm motility in Atlantic cod Gadus morhua. J. Fish Biol. 2015, 87, 54–68. [Google Scholar] [CrossRef]
- Zadmajid, V.; Myers, J.N.; Sørensen, S.R.; Butts, I.A.E. Ovarian fluid and its impacts on spermatozoa performance in fish: A review. Theriogenology 2019, 1, 144–152. [Google Scholar] [CrossRef]
- Splendiani, A.; Ruggeri, P.; Giovannotti, M.; Pesaresi, S.; Occhipinti, G.; Fioravanti, T.; Lorenzoni, M.; Nisi Cerioni, P.; Caputo Barucchi, V. Alien brown trout invasion of the Italian peninsula: The role of geological, climate and anthropogenic factors. Biol. Invasions 2016, 18, 2029–2044. [Google Scholar] [CrossRef]
- Berrebi, P. Three brown trout Salmo trutta lineages in Corsica described through allozyme variation. J. Fish Biol. 2015, 86, 60–73. [Google Scholar] [CrossRef]
- Sabatini, A.; Cannas, R.; Follesa, M.C.; Palmas, F.; Manunza, A.; Matta, G.; Pendugiu, A.A.; Serra, P.; Cau, A. Genetic characterization and artificial reproduction attempt of endemic Sardinian trout Salmo trutta L., 1758 (Osteichthyes, Salmonidae): Experiences in captivity. Ital. J. Zool. 2011, 78, 20–26. [Google Scholar] [CrossRef]
- Splendiani, A.; Fioravanti, T.; Giovannotti, M.; Olivieri, L.; Ruggeri, P.; Nisi Cerioni, P.; Vanni, S.; Enrichetti, F.; Caputo Barucchi, V. Museum samples could help to reconstruct the original distribution of Salmo trutta complex in Italy. J. Fish Biol. 2017, 90, 2443–2451. [Google Scholar] [CrossRef] [PubMed]
- Di Iorio, M.; Esposito, S.; Rusco, G.; Roncarati, A.; Miranda, M.; Gibertoni, P.P.; Cerolini, S.; Iaffaldano, N. Semen cryopreservation for the Mediterranean brown trout of the Biferno River (Molise-Italy): Comparative study on the effects of basic extenders and cryoprotectants. Sci. Rep. 2019, 9, 9703. [Google Scholar] [CrossRef] [PubMed]
- Rusco, G.; Di Iorio, M.; Gibertoni, P.P.; Esposito, S.; Penserini, M.; Roncarati, A.; Cerolini, S.; Iaffaldano, N. Optimization of Sperm Cryopreservation Protocol for Mediterranean Brown Trout: A Comparative Study of Non-Permeating Cryoprotectants and Thawing Rates In Vitro and In Vivo. Animals 2019, 9, 304. [Google Scholar] [CrossRef]
- Rusco, G.; Di Iorio, M.; Iampietro, R.; Esposito, S.; Gibertoni, P.P.; Penserini, M.; Roncarati, A.; Iaffaldano, N. A Simple and Efficient Semen Cryopreservation Method to Increase the Genetic Variability of Endangered Mediterranean Brown Trout Inhabiting Molise Rivers. Animals 2020, 10, 403. [Google Scholar] [CrossRef] [PubMed]
- Rusco, G.; Di Iorio, M.; Iampietro, R.; Roncarati, A.; Esposito, S.; Iaffaldano, N. Cryobank of Mediterranean Brown Trout Semen: Evaluation of the Use of Frozen Semen up to Six Hours Post-Collection. Fishes 2021, 6, 26. [Google Scholar] [CrossRef]
- Ferguson, A.; Reed, T.E.; Cross, T.F.; McGinnity, P.; Prodöhl, P.A. Anadromy, potamodromy and residency in brown trout Salmo trutta: The role of genes and the environment. J. Fish Biol. 2019, 95, 692–718. [Google Scholar] [CrossRef]
- Di Iorio, M.; Rusco, G.; Esposito, S.; D′Andrea, M.; Roncarati, A.; Iaffaldano, N. The role of semen cryobanks for protecting endangered native salmonids: Advantages and perspectives as outlined by the LIFE Nat.Sal.Mo. project on Mediterranean brown trout (Molise region-Italy). Front. Mar. Sci. 2023, 9, 1075498. [Google Scholar] [CrossRef]
- Palombo, V.; De Zio, E.; Salvatore, G.; Esposito, S.; Iaffaldano, N.; D′Andrea, M. Genotyping of Two Mediterranean Trout Populations in Central-Southern Italy for Conservation Purposes Using a Rainbow-Trout-Derived SNP Array. Animals 2021, 11, 1803. [Google Scholar] [CrossRef]
- Škraba Jurlina, D.; Marić, A.; Mrdak, D.; Kanjuh, T.; Špelić, I.; Nikolić, V.; Piria, M.; Simonović, P. Alternative life-history in native trout (Salmo spp.) suppresses the invasive effect of alien trout strains introduced into streams in the Western part of the Balkans. Front. Ecol. Evol. 2020, 8, 188. [Google Scholar] [CrossRef]
- Toews, D.P.; Brelsford, A. The biogeography of mitochondrial and nuclear discordance in animals. Mol. Ecol. 2012, 21, 3907–3930. [Google Scholar] [CrossRef]
- Lorenzoni, M.; Carosi, A.; Giovannotti, M.; La Porta, G.; Splendiani, A.; Barucchi, V.C. Ecology and conservation of the Mediterranean trout in the central Apennines (Italy). J. Limnol. 2019, 78. [Google Scholar] [CrossRef]
- D′Agaro, E.; Gibertoni, P.; Marroni, F.; Messina, M.; Tibaldi, E.; Esposito, S. Genetic and Phenotypic Characteristics of the Salmo trutta Complex in Italy. Appl. Sci. 2022, 12, 3219. [Google Scholar] [CrossRef]
- Polgar, G.; Iaia, M.; Righi, T.; Volta, P. The Italian Alpine and Subalpine trouts: Taxonomy, Evolution, and Conservation. Biology 2022, 11, 576. [Google Scholar] [CrossRef] [PubMed]
- Purchase, C.F.; Rooke, A.C. Freezing ovarian fluid does not alter how it affects fish sperm swimming performance: Creating a cryptic female choice ‘spice rack’ for use in split-ejaculate experimentation. J. Fish Biol. 2020, 96, 693–699. [Google Scholar] [CrossRef] [PubMed]
- Nynca, J.; Ciereszko, A. Measurement of concentration and viability of brook trout (Salvelinus fontinalis) spermatozoa using computer-aided fluorescent microscopy. Aquacilture 2009, 292, 256–258. [Google Scholar] [CrossRef]
- Billard, R. Reproduction in rainbow trout: Sex differentiation, dynamics of gametogenesis, biology and preservation of gametes. Aquaculture 1992, 100, 263–298. [Google Scholar] [CrossRef]
- Dadras, H.; Dzyuba, B.; Cosson, J.; Golpour, A.; Siddique, M.A.M.; Linhart, O. Effect of water temperature on the physiology of fish spermatozoon function: A brief review. Aquac. Res. 2016, 48, 729–740. [Google Scholar] [CrossRef]
- Petersen, C.W.; Warner, R.R.; Cohen, S.; Hess, H.C.; Sewell, A.T. Variable pelagic fertilization success: Implications for mate choice and spatial patterns of mating. Ecology 1992, 73, 391–401. [Google Scholar] [CrossRef]
- Petersen, C.W.; Warner, R.R. Sperm competition in fishes. In Sperm Competition and Sexual Selection; Birkhead, T.R., MØller, A.P., Eds.; Academic Press: San Jose, CA, USA, 1998; pp. 435–463. [Google Scholar]
- Kime, D.E.; Van Look, K.J.W.; McAllister, B.G.; Huyskens, G.; Rurangwa, E.; Ollevier, F. Computer-assisted sperm analysis (CASA) as a tool for monitoring sperm quality in fish. Comp. Biochem. Physiol. 2001, 130, 425–433. [Google Scholar] [CrossRef]
- Yanagimachi, R.; Cherr, G.; Matsubara, T.; Andoh, T.; Harumi, T.; Vines, C.; Pillai, M.; Griffin, F.; Matsubara, H.; Weatherby, T. Sperm attractant in the micropyle region of fish and insect eggs. Biol. Reprod. 2013, 88, 1–11. [Google Scholar] [CrossRef]
- Yanagimachi, R.; Harumi, T.; Matsubara, H.; Yan, W.; Yuan, S.; Hirohashi, N.; Iida, T.; Yamaha, E.; Arai, K.; Matsubara, T. Chemical and physical guidance of fish spermatozoa into the egg through the micropyle. Biol. Reprod. 2017, 96, 780–799. [Google Scholar] [CrossRef]
- Lahnsteiner, F.; Berger, B.; Weismann, T.; Patzner, R.A. Determination of semen quality of the rainbow trout, Oncorhynchus mykiss, by sperm motility, seminal plasma parameters, and spermatozoal metabolism. Aquaculture 1998, 163, 163–181. [Google Scholar] [CrossRef]
- Gage, M.J.G.; Macfarlane, C.P.; Yeates, S.; Ward, R.G.; Searle, J.B.; Parker, G.A. Spermatozoal traits and sperm competition in Atlantic salmon: Relative sperm velocity is the primary determinant of fertilization success. Curr. Biol. 2004, 14, 44–47. [Google Scholar] [CrossRef] [PubMed]
- Linhart, O.; Rodina, M.; Gela, D.; Kocour, M.; Vandeputte, M. Spermatozoal competition in common carp (Cyprinus carpio): What is the primary determinant of competition success? Reprod. Colch. 2005, 130, 705–711. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Male ID | Female ID | N-Z Group | |
|---|---|---|---|
| Sperm Trait | Significance | Significance | Significance |
| MOT (%) | *** | * | |
| VCL (µm/s) | *** | ** | *** |
| VAP (µm/s) | *** | ** | *** |
| VSL (µm/s) | *** | *** | |
| STR (%) | *** | * | ** |
| LIN (%) | *** | ** | |
| ALH (µm) | *** | *** | |
| BCF (Hz) | *** | * | |
| DSM (s) | *** | *** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rusco, G.; Di Iorio, M.; Esposito, S.; Gibertoni, P.; Antenucci, E.; Palombo, V.; Roncarati, A.; Iaffaldano, N. Zootechnical Brown Trout (Salmo trutta L. 1758) Ovarian Fluid Fails to Upregulate the Swimming Performances of Native Mediterranean Brown Trout (Salmo cettii Rafinesque, 1810) Sperm in the Biferno River. Fishes 2023, 8, 190. https://doi.org/10.3390/fishes8040190
Rusco G, Di Iorio M, Esposito S, Gibertoni P, Antenucci E, Palombo V, Roncarati A, Iaffaldano N. Zootechnical Brown Trout (Salmo trutta L. 1758) Ovarian Fluid Fails to Upregulate the Swimming Performances of Native Mediterranean Brown Trout (Salmo cettii Rafinesque, 1810) Sperm in the Biferno River. Fishes. 2023; 8(4):190. https://doi.org/10.3390/fishes8040190
Chicago/Turabian StyleRusco, Giusy, Michele Di Iorio, Stefano Esposito, Pierpaolo Gibertoni, Emanuele Antenucci, Valentino Palombo, Alessandra Roncarati, and Nicolaia Iaffaldano. 2023. "Zootechnical Brown Trout (Salmo trutta L. 1758) Ovarian Fluid Fails to Upregulate the Swimming Performances of Native Mediterranean Brown Trout (Salmo cettii Rafinesque, 1810) Sperm in the Biferno River" Fishes 8, no. 4: 190. https://doi.org/10.3390/fishes8040190
APA StyleRusco, G., Di Iorio, M., Esposito, S., Gibertoni, P., Antenucci, E., Palombo, V., Roncarati, A., & Iaffaldano, N. (2023). Zootechnical Brown Trout (Salmo trutta L. 1758) Ovarian Fluid Fails to Upregulate the Swimming Performances of Native Mediterranean Brown Trout (Salmo cettii Rafinesque, 1810) Sperm in the Biferno River. Fishes, 8(4), 190. https://doi.org/10.3390/fishes8040190