Complete Mitochondrial Genome and Phylogenetic Position of Chirolophis wui (Perciformes: Stichaeidae)

 ,
,  , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
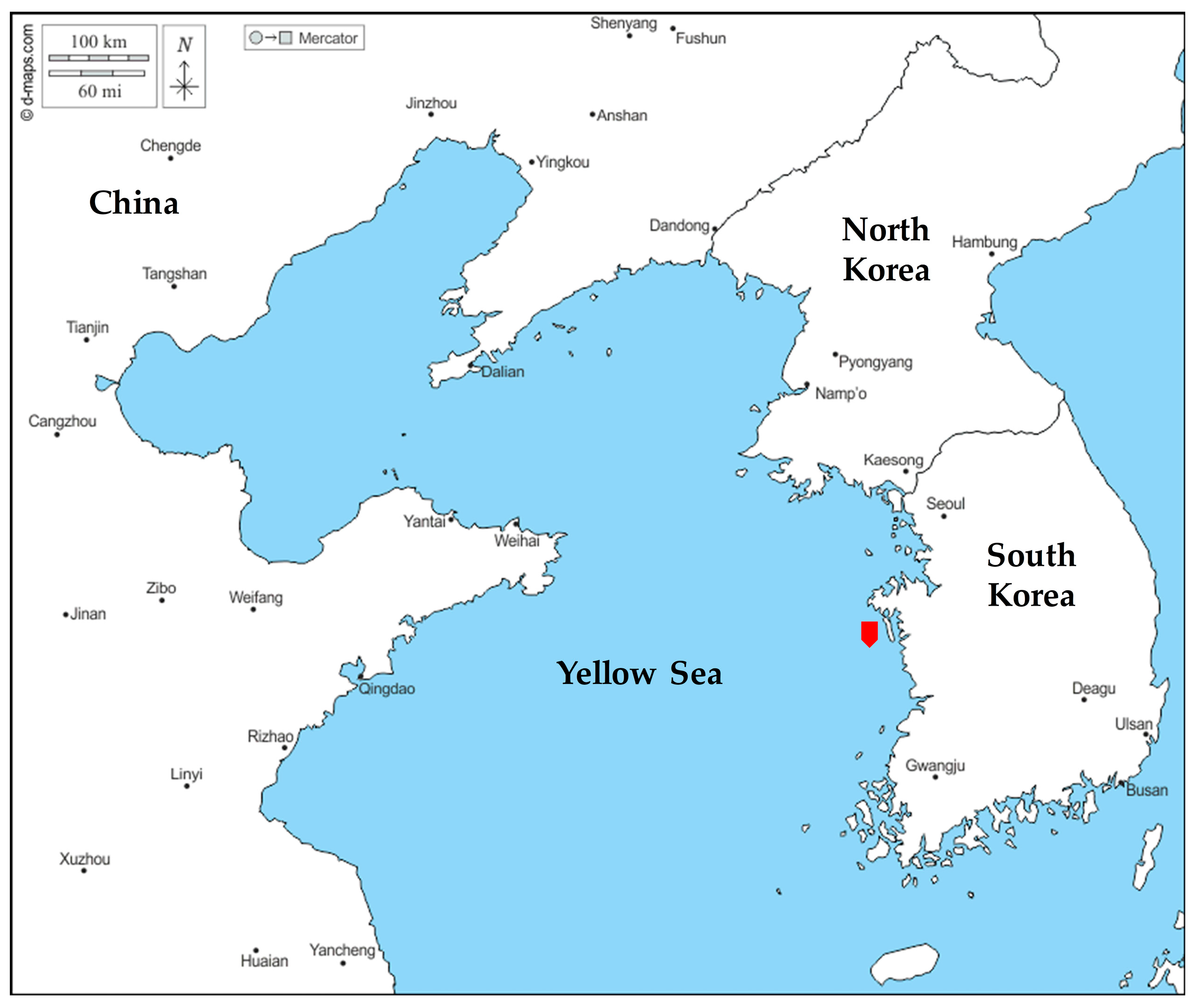
2.1. Sample and DNA Extraction
2.2. Next-Generation Sequencing and Mitogenome Assembly
2.3. Mitogenome Annotation, Sequence Analysis, and Phylogenetic Analysis
3. Results and Discussion
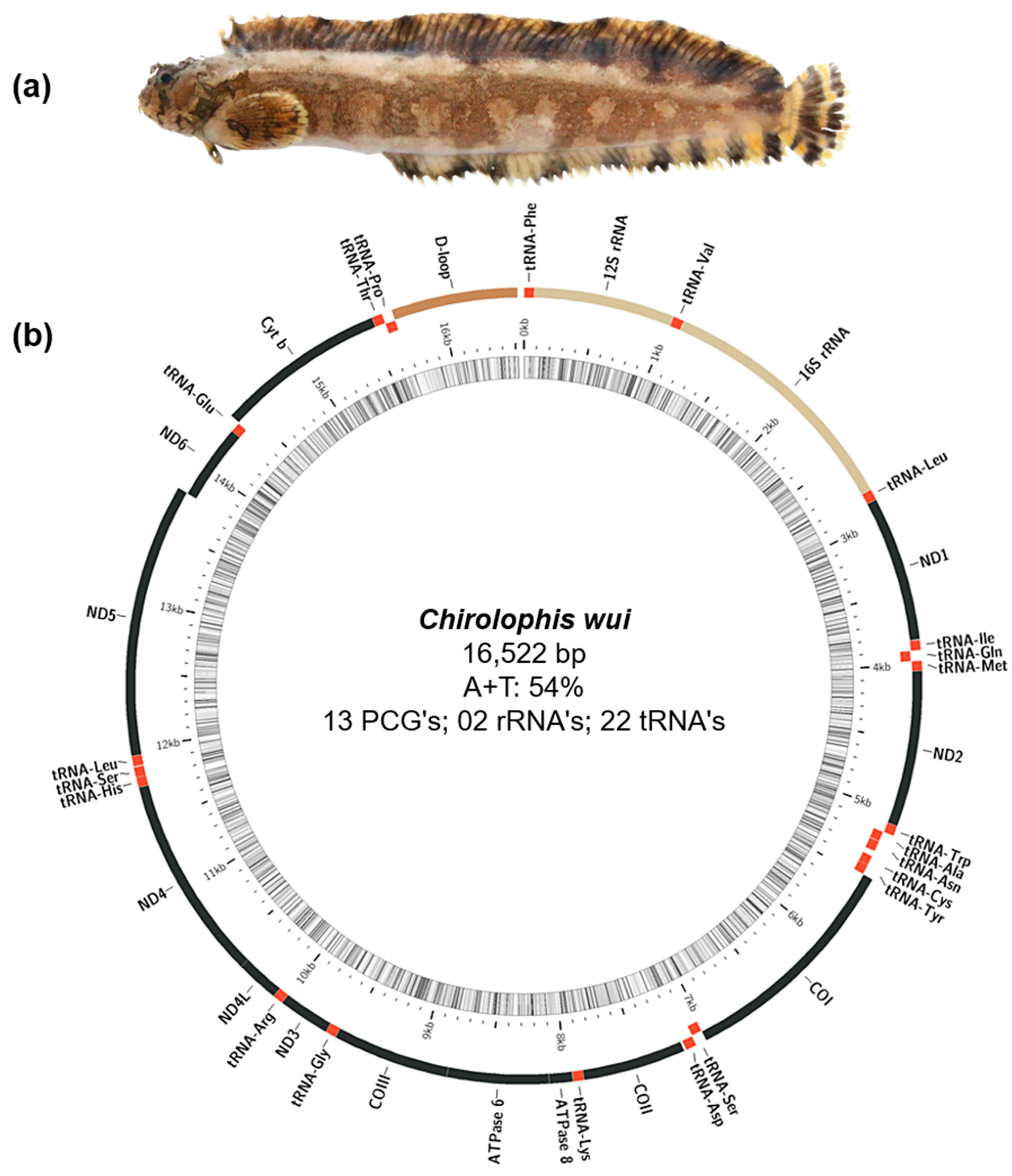
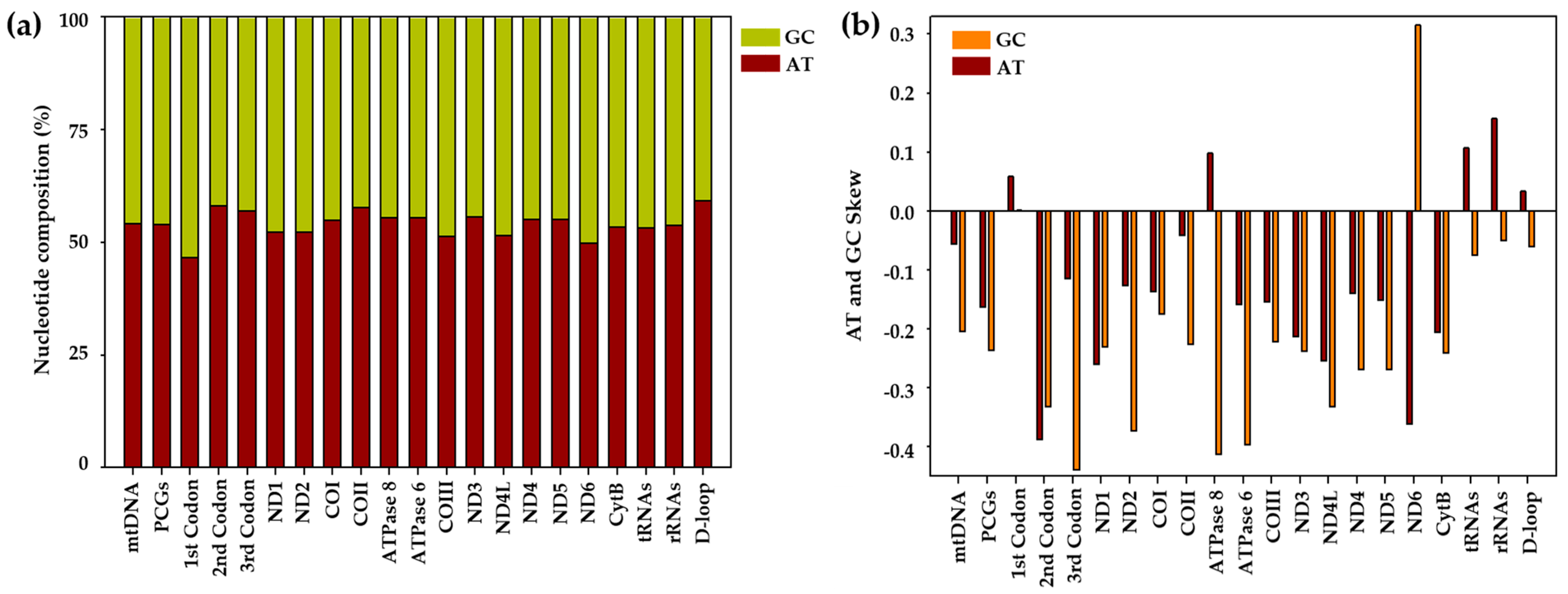
3.1. Mitochondrial Genome Assembly, Annotation, and Sequence Analysis
3.2. Protein Coding Genes
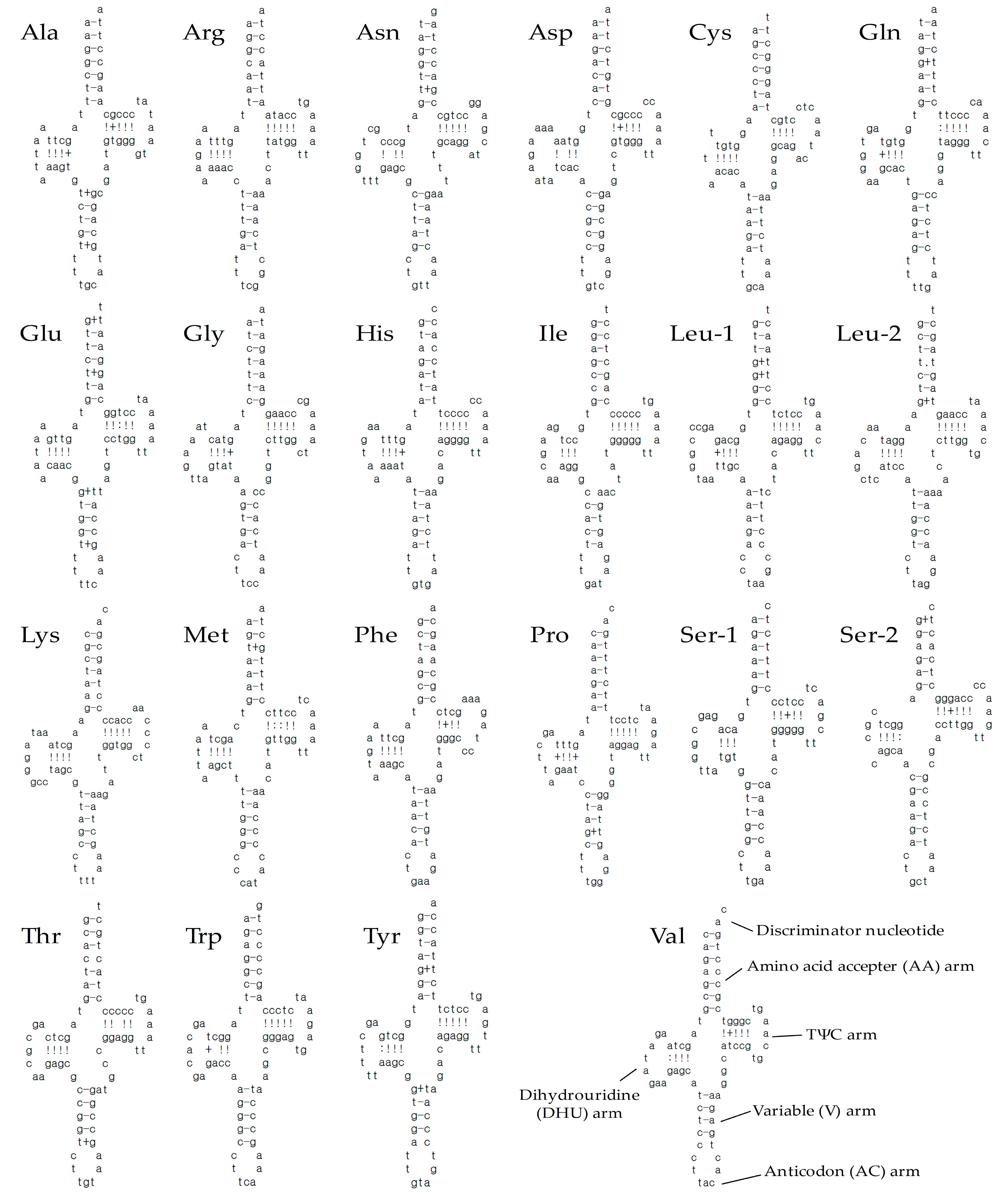
3.3. Ribosomal RNA, Transfer RNA Genes, and Control Region
3.4. Phylogenetic Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sun, C.-H.; Liu, H.-Y.; Xu, N.; Zhang, X.-L.; Zhang, Q.; Han, B.-P. Mitochondrial genome structures and phylogenetic analyses of two tropical characidae fishes. Front. Genet. 2021, 12, 627402. [Google Scholar] [CrossRef]
- Parhi, J.; Tripathy, P.S.; Priyadarshi, H.; Mandal, S.C.; Pandey, P.K. Diagnosis of mitogenome for robust phylogeny: A case of Cypriniformes fish group. Gene 2019, 713, 143967. [Google Scholar] [CrossRef]
- Avise, J.C.; Arnold, J.; Ball, R.M.; Bermingham, E.; Lamb, T.; Neigel, J.E.; Reeb, C.A.; Saunders, N.C. Intraspecific phylogeography: The mitochondrial DNA bridge between population genetics and systematics. Ann. Rev. Ecol. System. 1987, 18, 489–522. [Google Scholar] [CrossRef]
- Prosdocimi, F.; de Carvalho, D.C.; de Almeida, R.N.; Beheregaray, L.B. The complete mitochondrial genome of two recently derived species of the fish genus Nannoperca (Perciformes, Percichthyidae). Mol. Biol. Rep. 2012, 39, 2767–2772. [Google Scholar] [CrossRef]
- Castro Paz, F.P.; Batista, J.d.S.; Porto, J.I.R. DNA barcodes of rosy tetras and allied species (Characiformes: Characidae: Hyphessobrycon) from the Brazilian Amazon basin. PLoS ONE 2014, 9, e98603. [Google Scholar] [CrossRef] [PubMed]
- Anderson, S.; Bankier, A.T.; Barrell, B.G.; de Bruijn, M.H.; Coulson, A.R.; Drouin, J.; Eperon, I.C.; Nierlich, D.P.; Roe, B.A.; Sanger, F. Sequence and organization of the human mitochondrial genome. Nature 1981, 290, 457–465. [Google Scholar] [CrossRef]
- Tzeng, C.-S.; Hui, C.-F.; Shen, S.-C.; Huang, P. The complete nucleotide sequence of the Crossostoma lacustre mitochondrial genome: Conservation and variations among vertebrates. Nucleic Acids Res. 1992, 20, 4853–4858. [Google Scholar] [CrossRef]
- Billington, N.; Hebert, P.D. Mitochondrial DNA diversity in fishes and its implications for introductions. Can. J. Fish. Aquat. Sci. 1991, 48, 80–94. [Google Scholar] [CrossRef]
- Alam, M.T.; Petit, R.A., III; Read, T.D.; Dove, A.D. The complete mitochondrial genome sequence of the world’s largest fish, the whale shark (Rhincodon typus), and its comparison with those of related shark species. Gene 2014, 539, 44–49. [Google Scholar] [CrossRef]
- Deng, Y.-P.; Li, R.; Wang, H.-M.; Liu, G.-H.; Tu, Y. Complete Mitochondrial Genome of Contracaecum Sp. (Nematoda: Ascarididae) from Night Herons in China. J. Nematol. 2022, 54, 20220048. [Google Scholar] [CrossRef] [PubMed]
- Satoh, T.P.; Miya, M.; Mabuchi, K.; Nishida, M. Structure and variation of the mitochondrial genome of fishes. BMC Genom. 2016, 17, 719. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.I.; Baek, J.Y.; Kim, M.J.; Jeong, H.C.; Kim, K.-G.; Bae, C.H.; Han, Y.S.; Jin, B.R.; Kim, I. Complete nucleotide sequence and organization of the mitogenome of the red-spotted apollo butterfly, Parnassius bremeri (Lepidoptera: Papilionidae) and comparison with other lepidopteran insects. Mol. Cells 2009, 28, 347–363. [Google Scholar] [CrossRef] [PubMed]
- Gray, M.W. Origin and evolution of mitochondrial DNA. Annu. Rev. Cell Biol. 1989, 5, 25–50. [Google Scholar] [CrossRef]
- Wang, L.; Chen, Z.; Gao, J.; Zhao, Y.; Sun, P.; Lu, K. The complete mitochondrial genome of Indonesian tiger fish Datnioides microlepis (Bleeker 1854). Mitochondrial DNA B Resour. 2016, 1, 328–329. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Wang, X.; Liu, S.; Luo, H.; Lin, Q. Population genetic structure and phylogenetic analysis of gray’s pipefish, Halicampus grayi in the South China Sea. Genes Genom. 2020, 42, 155–164. [Google Scholar] [CrossRef]
- Mecklenburg, C. Family Stichaeidae Gill 1864-pricklebacks. California Academy of Sciences. Annot. Check List. Fishes 2004, 35, 1–36. [Google Scholar]
- Nelson, J.S.; Grande, T.C.; Wilson, M.V. Fishes of the World; John Wiley & Sons: Hoboken, NJ, USA, 2016. [Google Scholar]
- Gill, T. Note on the family of stichaeoids. Proc. Acad. Nat. Sci. Phila. 1864, 16, 208–211. [Google Scholar]
- Jordan, D.S.; Evermann, B.W. The Fishes of North and Middle America: A Descriptive Catalogue of the Species of Fish-Like Vertebrates Found in the Waters of North America, North of the Isthmus of Panama; US Government Printing Office: Washington, DC, USA, 1896.
- Liu, L.; Liu, Q.; Gao, T. Genome-wide survey reveals the phylogenomic relationships of Chirolophis japonicus Herzenstein, 1890 (Stichaeidae, Perciformes). ZooKeys 2022, 1129, 52–72. [Google Scholar] [CrossRef]
- Nakabo, T. Fishes of Japan with Pictorial Keys to the Species, English Edition; Tokai University Press: Tokyo, Japan, 2002. [Google Scholar]
- Kim, I.-S.; Choi, S.-H. Fishes of the Southwestern Coast of Korea. Anim. Syst. Evol. Divers. 1998, 14, 135–157. [Google Scholar]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data; Babraham Institute: Cambridge, UK, 2010. [Google Scholar]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed]
- Bernt, M.; Donath, A.; Jühling, F.; Externbrink, F.; Florentz, C.; Fritzsch, G.; Pütz, J.; Middendorf, M.; Stadler, P.F. MITOS: Improved de novo metazoan mitochondrial genome annotation. Mol. Phylogenet. Evol. 2013, 69, 313–319. [Google Scholar] [CrossRef]
- Stothard, P. The sequence manipulation suite: JavaScript programs for analyzing and formatting protein and DNA sequences. Biotechniques 2000, 28, 1102–1104. [Google Scholar] [CrossRef]
- Benson, G. Tandem repeats finder: A program to analyze DNA sequences. Nucleic Acids Res. 1999, 27, 573–580. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547. [Google Scholar] [CrossRef]
- Perna, N.T.; Kocher, T.D. Patterns of nucleotide composition at fourfold degenerate sites of animal mitochondrial genomes. J. Mol. Evol. 1995, 41, 353–358. [Google Scholar] [CrossRef]
- Laslett, D.; Canbäck, B. ARWEN: A program to detect tRNA genes in metazoan mitochondrial nucleotide sequences. Bioinformatics 2008, 24, 172–175. [Google Scholar] [CrossRef]
- Thompson, J.D.; Gibson, T.J.; Higgins, D.G. Multiple sequence alignment using ClustalW and ClustalX. Curr. Protoc. Bioinformatics 2003, 1, 2–3. [Google Scholar] [CrossRef]
- Felsenstein, J. Evolutionary trees from DNA sequences: A maximum likelihood approach. J. Molecular Evol. 1981, 17, 368–376. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Fan, M.; Wang, S.; Gu, Z.; Wang, A.; Liu, C.; Yang, Y.; Liu, S. The Complete Mitochondrial Genome of Hyotissa hyotis (Bivalvia: Gryphaeidae) Reveals a Unique Gene Order within Ostreoidea. Fishes 2022, 7, 317. [Google Scholar] [CrossRef]
- Yu, H.; Li, Q. Complete mitochondrial DNA sequence of Crassostrea nippona: Comparative and phylogenomic studies on seven commercial Crassostrea species. Mol. Biol. Rep. 2012, 39, 999–1009. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Xu, X.; Yu, Z.; Wei, Z.; Xia, J. Comparison of seven Crassostrea mitogenomes and phylogenetic analyses. Mol. Phylogenet. Evol. 2010, 57, 448–454. [Google Scholar] [CrossRef] [PubMed]
- Ojala, D.; Montoya, J.; Attardi, G. tRNA punctuation model of RNA processing in human mitochondria. Nature 1981, 290, 470–474. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Li, X.; Li, L.; Xu, X.; Xia, J.; Yu, Z. New features of Asian Crassostrea oyster mitochondrial genomes: A novel alloacceptor tRNA gene recruitment and two novel ORFs. Gene 2012, 507, 112–118. [Google Scholar] [CrossRef]
- Boore, J.L. Animal mitochondrial genomes. Nucleic Acids Res. 1999, 27, 1767–1780. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.-H.; Sun, P.-Y.; Lao, Y.-L.; Wu, T.; Zhang, Y.-N.; Huang, Q.; Zhang, Q. Mitogenome of a monotypic genus, Oliotius Kottelat, 2013 (Cypriniformes: Cyprinidae): Genomic characterization and phylogenetic position. Gene 2023, 851, 147035. [Google Scholar] [CrossRef] [PubMed]
- Su, Z.; Wilson, B.; Kumar, P.; Dutta, A. Noncanonical roles of tRNAs: tRNA fragments and beyond. Annu. Rev. Genet. 2020, 54, 47–69. [Google Scholar] [CrossRef]
- Kundu, S.; Binarao, J.D.; De Alwis, P.S.; Kim, A.R.; Lee, S.-R.; Andriyono, S.; Gietbong, F.Z.; Kim, H.-W. First Mitogenome of Endangered Enteromius thysi (Actinopterygii: Cypriniformes: Cyprinidae) from Africa: Characterization and Phylogeny. Fishes 2022, 8, 25. [Google Scholar] [CrossRef]
- Zhang, D.-X.; Hewitt, G.M. Insect mitochondrial control region: A review of its structure, evolution and usefulness in evolutionary studies. Biochem. Syst. Ecol. 1997, 25, 99–120. [Google Scholar] [CrossRef]
- Balakirev, E.S.; Saveliev, P.A.; Ayala, F.J. Complete mitochondrial genomes of the Cherskii’s sculpin Cottus czerskii and Siberian taimen Hucho taimen reveal GenBank entry errors: Incorrect species identification and recombinant mitochondrial genome. Evol. Bioinform. 2017, 13, 1176934317726783. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subfamily | Species | Length (bp) | (A + T)% | (G + C)% | AT-Skew | GC-Skew | Accession No. |
|---|---|---|---|---|---|---|---|
| Chirolophinae | Chirolophis wui | 16,522 | 54.0 | 46.0 | −0.0556 | −0.2043 | OP388414 |
| Chirolophis japonicus | 16,521 | 54.1 | 45.9 | −0.0573 | −0.2026 | KT266879 | |
| Chirolophis ascanii | 16,520 | 53.8 | 46.2 | −0.0558 | −0.2078 | MT410928 | |
| Stichaeinae | Stichaeus fuscus | 16,529 | 55.4 | 44.6 | −0.0325 | −0.2108 | NC063112 |
| Stichaeus grigorjewi | 16,532 | 54.5 | 45.5 | −0.0385 | −0.2044 | NC045382 | |
| Stichaeus nozawae | 16,530 | 54.6 | 45.3 | −0.0366 | −0.2053 | NC046850 | |
| Opisthocentrinae | Opisthocentrus zonope | 16,518 | 53.8 | 46.2 | −0.0186 | −0.2208 | NC062676 |
| Opisthocentrus ocellatus | 16,517 | 54.1 | 45.9 | −0.0166 | −0.2244 | NC045921 | |
| Opisthocentrus tenuis | 16,515 | 54.1 | 45.9 | −0.0166 | −0.2244 | MT006232 |
| Gene | Strand | Location | Size (bp) | Start Codon | Stop Codon | Anticodon | Intergenic Nucleotides |
|---|---|---|---|---|---|---|---|
| tRNA-Phe | H | 1–68 | 68 | GAA | 0 | ||
| 12S rRNA | H | 69–1016 | 948 | 0 | |||
| tRNA-Val | H | 1017–1088 | 72 | TAC | 0 | ||
| 16S rRNA | H | 1089–2781 | 1693 | 0 | |||
| tRNA-Leu | H | 2782–2855 | 74 | TAA | 0 | ||
| ND1 | H | 2586–3830 | 975 | ATG | TAG | 3 | |
| tRNA-Ile | H | 3834–3903 | 69 | GAT | 0 | ||
| tRNA-Gln | L | 3903–3973 | 71 | TTG | −1 | ||
| tRNA-Met | H | 3973–4041 | 69 | CAT | −1 | ||
| ND2 | H | 4042–5087 | 1046 | ATG | TA- | 0 | |
| tRNA-Trp | H | 5088–5158 | 71 | TCA | 1 | ||
| tRNA-Ala | L | 5160–5228 | 69 | TGC | 1 | ||
| tRNA-Asn | L | 5230–5302 | 73 | GTT | 38 | ||
| tRNA-Cys | L | 5341–5406 | 66 | GCA | 0 | ||
| tRNA-Tyr | L | 5407–5477 | 71 | GTA | 1 | ||
| COI | H | 5479–7029 | 1551 | GTG | TAA | 0 | |
| tRNA-Ser | L | 7030–7100 | 71 | TGA | 3 | ||
| tRNA-Asp | H | 7104–7176 | 73 | GTC | 14 | ||
| COII | H | 7191–7881 | 691 | ATG | T-- | 0 | |
| tRNA-Lys | H | 7882–7955 | 74 | TTT | 1 | ||
| ATPase 8 | H | 7957–8124 | 168 | ATG | TAA | 0 | |
| ATPase 6 | H | 8115–8797 | 683 | ATG | TA- | −10 | |
| COIII | H | 8798–9582 | 785 | ATG | TA- | 0 | |
| tRNA-Gly | H | 9583–9654 | 72 | TCC | 0 | ||
| ND3 | H | 9655–10,003 | 349 | ATG | T-- | 0 | |
| tRNA-Arg | H | 10,004–10,072 | 69 | TCG | 0 | ||
| ND4L | H | 10,073–10,369 | 297 | ATG | TAA | 0 | |
| ND4 | H | 10,363–11,743 | 381 | ATG | T-- | −8 | |
| tRNA-His | H | 11,744–11,813 | 70 | GTG | 0 | ||
| tRNA-Ser | H | 11,814–11,881 | 68 | GCT | 0 | ||
| tRNA-Leu | H | 11,886–11,958 | 73 | TAG | 4 | ||
| ND5 | H | 11,959–13,797 | 1839 | ATG | TAA | 0 | |
| ND6 | L | 13,794–14,315 | 522 | ATG | TAA | −4 | |
| tRNA-Glu | L | 14,316–14,384 | 69 | TTC | 0 | ||
| Cytb | H | 14,390–15,530 | 1141 | ATG | T-- | 5 | |
| tRNA-Thr | H | 15,531–15,602 | 72 | TGT | 0 | ||
| tRNA-Pro | L | 15,602–15,671 | 70 | TGG | −1 | ||
| D-loop | H | 15,672–16,522 | 851 | 0 |
| C. wui | Size | T% | C% | A% | G% | AT% | AT-Skew | GC-Skew |
|---|---|---|---|---|---|---|---|---|
| mitogenome | 16,522 | 28.5 | 27.7 | 25.5 | 18.3 | 54.0 | −0.0556 | −0.2043 |
| PCGs | 11,400 | 31.3 | 28.5 | 22.5 | 17.6 | 53.8 | −0.1636 | −0.2364 |
| 1st codon | - | 21.9 | 26.7 | 24.6 | 26.8 | 46.5 | 0.0581 | 0.0019 |
| 2nd codon | - | 40.3 | 27.9 | 17.8 | 14.0 | 58.1 | −0.3873 | −0.3317 |
| 3rd codon | - | 31.7 | 31.0 | 25.2 | 12.1 | 56.9 | −0.1142 | −0.4385 |
| tRNAs | 1555 | 23.7 | 25.2 | 29.4 | 21.7 | 53.1 | 0.1073 | −0.0746 |
| rRNAs | 2641 | 22.6 | 24.3 | 31.0 | 22.0 | 53.6 | 0.1567 | −0.0497 |
| D-loop | 851 | 28.6 | 21.7 | 30.6 | 19.2 | 59.2 | 0.0338 | −0.0611 |
| AA | Codon | N | /1000 | Freq. | AA | Codon | N | /1000 | Freq. |
|---|---|---|---|---|---|---|---|---|---|
| Ala | GCG | 32 | 8.42 | 0.09 | Pro | CCG | 18 | 4.74 | 0.08 |
| GCA | 84 | 22.11 | 0.24 | CCA | 30 | 7.89 | 0.14 | ||
| GCT | 94 | 24.74 | 0.26 | CCT | 78 | 20.53 | 0.35 | ||
| GCC | 145 | 38.16 | 0.41 | CCC | 96 | 25.26 | 0.43 | ||
| Cys | TGT | 8 | 2.11 | 0.31 | Gln | CAG | 38 | 10.00 | 0.37 |
| TGC | 18 | 4.74 | 0.69 | CAA | 64 | 16.84 | 0.63 | ||
| Asp | GAT | 31 | 8.16 | 0.40 | Arg | CGG | 11 | 2.89 | 0.14 |
| GAC | 46 | 12.11 | 0.60 | CGA | 30 | 7.89 | 0.39 | ||
| Glu | GAG | 35 | 9.21 | 0.36 | CGT | 17 | 4.47 | 0.22 | |
| GAA | 62 | 16.32 | 0.64 | CGC | 18 | 4.74 | 0.24 | ||
| Phe | TTT | 139 | 36.58 | 0.60 | Ser | AGG | 0 | 0.00 | 0.00 |
| TTC | 94 | 24.74 | 0.40 | AGA | 0 | 0.00 | 0.00 | ||
| Gly | GGG | 49 | 12.89 | 0.19 | AGT | 25 | 6.58 | 0.10 | |
| GGA | 64 | 16.84 | 0.25 | AGC | 33 | 8.68 | 0.13 | ||
| GGT | 47 | 12.37 | 0.19 | TCG | 23 | 6.05 | 0.09 | ||
| GGC | 93 | 24.47 | 0.37 | TCA | 38 | 10.00 | 0.15 | ||
| His | CAT | 48 | 12.63 | 0.45 | TCT | 55 | 14.47 | 0.22 | |
| CAC | 59 | 15.53 | 0.55 | TCC | 75 | 19.74 | 0.30 | ||
| Ile | ATT | 169 | 44.47 | 0.67 | Thr | ACG | 29 | 7.63 | 0.10 |
| ATC | 83 | 21.84 | 0.33 | ACA | 78 | 20.53 | 0.27 | ||
| Lys | AAG | 16 | 4.21 | 0.23 | ACT | 68 | 17.89 | 0.23 | |
| AAA | 55 | 14.47 | 0.77 | ACC | 117 | 30.79 | 0.40 | ||
| Leu | TTG | 42 | 11.05 | 0.06 | Val | GTG | 19 | 5.00 | 0.08 |
| TTA | 113 | 29.74 | 0.17 | GTA | 70 | 18.42 | 0.30 | ||
| CTG | 41 | 10.79 | 0.06 | GTT | 103 | 27.11 | 0.44 | ||
| CTA | 108 | 28.42 | 0.16 | GTC | 44 | 11.58 | 0.19 | ||
| CTT | 227 | 59.74 | 0.34 | Trp | TTG | 33 | 8.68 | 0.28 | |
| CTC | 131 | 34.47 | 0.20 | TGA | 86 | 22.63 | 0.72 | ||
| Met | ATG | 73 | 19.21 | 0.49 | Tyr | TAT | 51 | 13.42 | 0.46 |
| ATA | 76 | 20.00 | 0.51 | TAC | 59 | 15.53 | 0.54 | ||
| Asn | AAT | 45 | 11.84 | 0.40 | |||||
| AAC | 67 | 17.63 | 0.60 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, Y.-S.; Patil, M.P.; Kim, J.-O.; Lee, Y.-J.; Seo, Y.B.; Kim, J.-K.; Mahale, K.R.; Kim, G.-D. Complete Mitochondrial Genome and Phylogenetic Position of Chirolophis wui (Perciformes: Stichaeidae). Fishes 2023, 8, 165. https://doi.org/10.3390/fishes8030165
Lee Y-S, Patil MP, Kim J-O, Lee Y-J, Seo YB, Kim J-K, Mahale KR, Kim G-D. Complete Mitochondrial Genome and Phylogenetic Position of Chirolophis wui (Perciformes: Stichaeidae). Fishes. 2023; 8(3):165. https://doi.org/10.3390/fishes8030165
Chicago/Turabian StyleLee, Yong-Suk, Maheshkumar Prakash Patil, Jong-Oh Kim, Yu-Jin Lee, Yong Bae Seo, Jin-Koo Kim, Kiran R. Mahale, and Gun-Do Kim. 2023. "Complete Mitochondrial Genome and Phylogenetic Position of Chirolophis wui (Perciformes: Stichaeidae)" Fishes 8, no. 3: 165. https://doi.org/10.3390/fishes8030165
APA StyleLee, Y.-S., Patil, M. P., Kim, J.-O., Lee, Y.-J., Seo, Y. B., Kim, J.-K., Mahale, K. R., & Kim, G.-D. (2023). Complete Mitochondrial Genome and Phylogenetic Position of Chirolophis wui (Perciformes: Stichaeidae). Fishes, 8(3), 165. https://doi.org/10.3390/fishes8030165