
Evolutionary Conservation of Photophore Ultrastructure in Sharks: The Case of a Dalatiid Squalomorph

 ,
,  ,
, 
 and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Specimen Sampling
2.2. Microscopy Observation
2.3. Micrography Analyses
3. Results
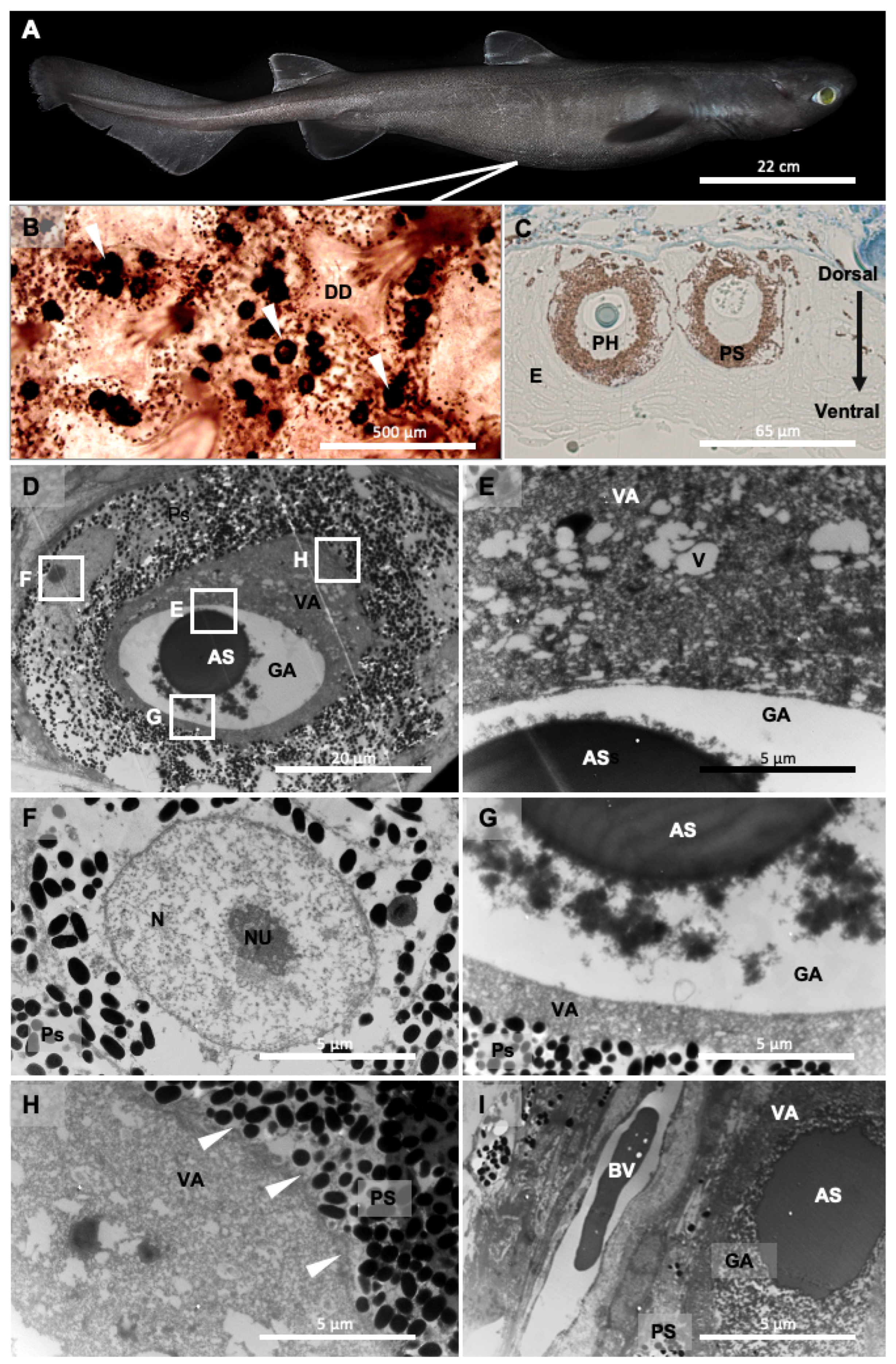
3.1. Photophore Ultrastructure
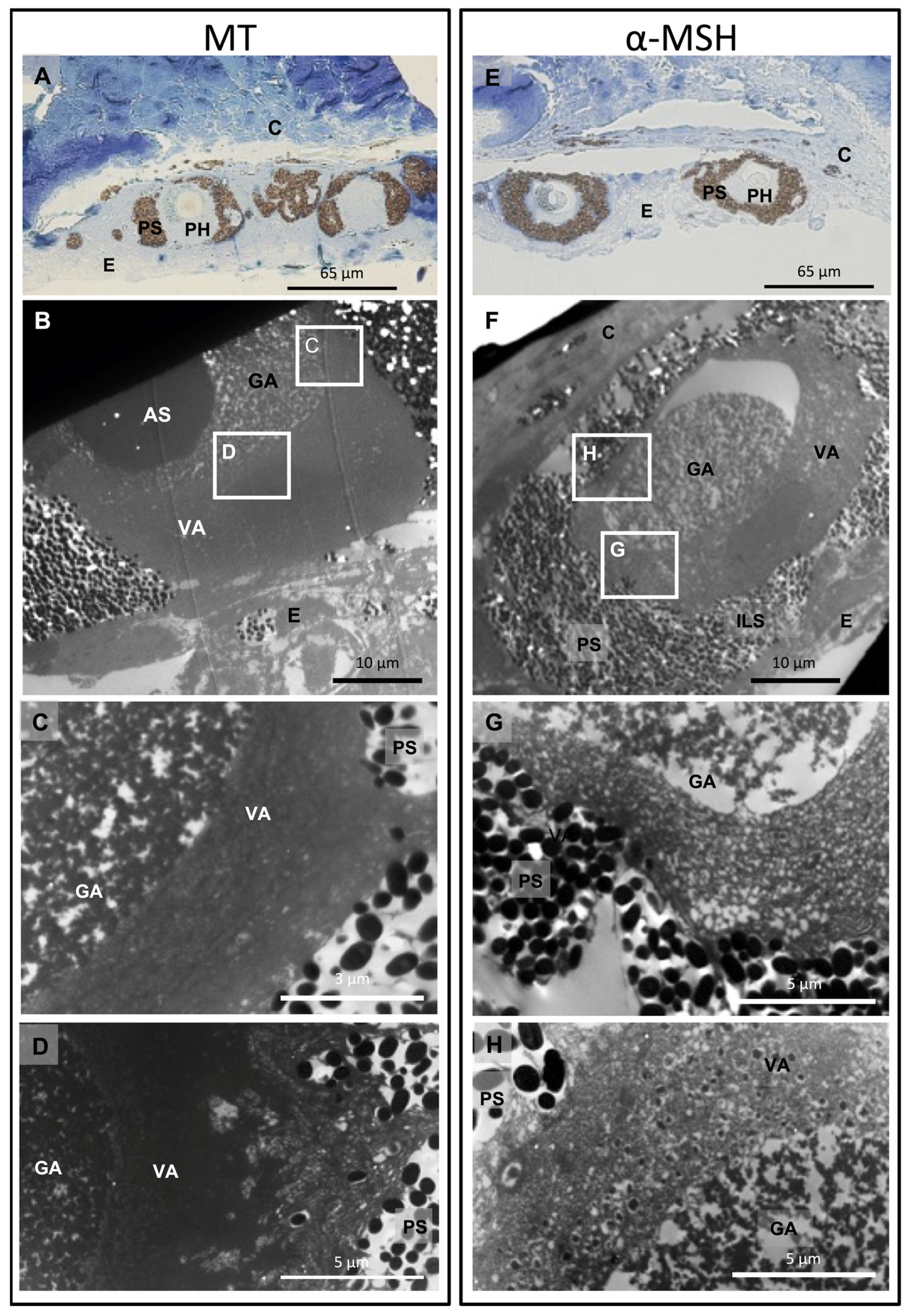
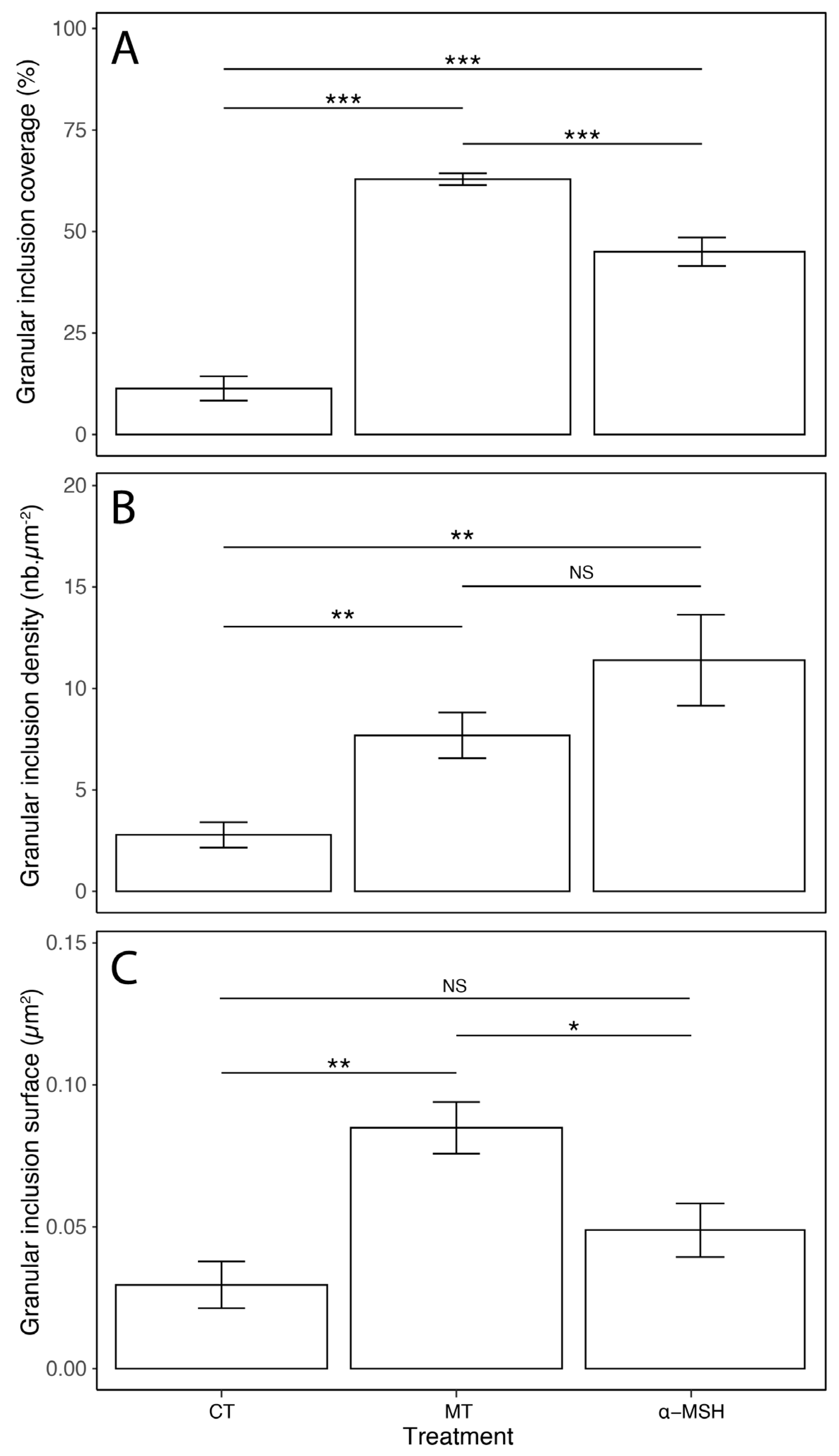
3.2. Cytological Changes upon Hormonal Treatments
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Haddock, S.H.D.; Moline, M.A.; Case, J.F. Bioluminescence in the sea. Annu. Rev. Mar. Sci. 2010, 2, 443–493. [Google Scholar] [CrossRef]
- Widder, E.A. Bioluminescence in the Ocean: Origins of biological, chemical, and ecological diversity. Science 2010, 328, 704–708. [Google Scholar] [CrossRef] [PubMed]
- Anctil, M. Physiological control of bioluminescence. Photochem. Photobiol. 1979, 30, 777–780. [Google Scholar] [CrossRef]
- Anctil, M. Neural Control Mechanisms in Bioluminescence. In Nervous Systems in Invertebrates; Ali, M.A., Ed.; Springer: Boston, MA, USA, 1987; pp. 573–602. [Google Scholar]
- Wilson, T.; Hastings, J.W. Bioluminescence. Annu. Rev. Cell Dev. Biol. 1998, 14, 197–230. [Google Scholar] [CrossRef]
- Herring, P.J. Aspects of the bioluminescence of fishes. In Oceanography and Marine Biology; Barnes, M., Ed.; Aberdeen University Press: Aberdeen, UK, 1982; pp. 415–470. [Google Scholar]
- Anctil, M. Ultrastructure of the luminescent system of the ctenophore Mnemiopsis leidyi. Cell Tissue Res. 1985, 242, 333–340. [Google Scholar] [CrossRef]
- Germain, G.; Anctil, M. Luminescent activity and ultrastructural characterization of photocytes dissociated from the coelenterate Renilla köllikeri. Tissue Cell 1988, 20, 701–720. [Google Scholar] [CrossRef] [PubMed]
- Hastings, J.W.; Morin, J.G. Bioluminescence. In Neural and Integrative Animal Physiology; Prosser, C.L., Ed.; Wiley-Liss: New-York, NY, USA, 1991; pp. 131–170. [Google Scholar]
- Herring, P.J.; Dilly, P.N.; Cope, C. The photophores of the squid family Cranchiidae (Cephalopoda: Oegopsida). J. Zool. 2002, 258, 73–90. [Google Scholar] [CrossRef]
- Renwart, M.; Delroisse, J.; Claes, J.M.; Mallefet, J. Ultrastructural organization of lantern shark (Etmopterus spinax Linnaeus, 1758) photophores. Zoomorphology 2014, 133, 405–416. [Google Scholar] [CrossRef]
- Haneda, Y.; Tsuji, F.I. Light production in the luminous fishes Photoblepharon and Anomalops from the Banda Islands. Science 1971, 173, 143–145. [Google Scholar] [CrossRef]
- Tebo, B.M.; Scott Linthicum, D.; Nealson, K.H. Luminous bacteria and light emitting fish: Ultrastructure of the symbiosis. Biosystems 1979, 11, 269–280. [Google Scholar] [CrossRef]
- Haygood, M.G. Light a organ symbioses in fishes. Crit. Rev. Microbiol. 1993, 19, 191–216. [Google Scholar] [CrossRef] [PubMed]
- Nowel, M.S.; Shelton, P.M.J.; Herring, P.J. Cuticular photophores of two decapod crustaceans, Oplophorus spinosus and Systellaspis debilis. Biol. Bull. 1998, 195, 290–307. [Google Scholar] [CrossRef] [PubMed]
- Bracken-Grissom, H.D.; DeLeo, D.M.; Porter, M.L.; Iwanicki, T.; Sickles, J.; Frank, T.M. Light organ photosensitivity in deep-sea shrimp may suggest a novel role in counterillumination. Sci. Rep. 2020, 10, 4485. [Google Scholar] [CrossRef]
- Anctil, M. Development of bioluminescence and photophores in the midshipman fish, Porichthys notatus. J. Morphol. 1977, 151, 363–395. [Google Scholar] [CrossRef]
- Cavallero, M.; Mammola, C.L.; Verdiglione, R. Structural and ultrastructural comparison of photophores of two species of deep-sea fishes: Argyropelecus hemigymnus and Maurolicus muelleri. J. Fish Biol. 2004, 64, 1552–1567. [Google Scholar] [CrossRef]
- Herring, P.J.; Cope, C. Red bioluminescence in fishes: On the suborbital photophores of Malacosteus, Pachystomias and Aristostomias. Mar. Biol. 2005, 148, 383–394. [Google Scholar] [CrossRef]
- Mallefet, J.; Duchatelet, L.; Hermans, C.; Baguet, F. Luminescence control of Stomiidae photophores. Acta Histochem. 2019, 121, 7–15. [Google Scholar] [CrossRef]
- Duchatelet, L.; Claes, J.M.; Delroisse, J.; Flammang, P.; Mallefet, J. Glow on sharks: State of the art on bioluminescence research. Oceans 2021, 2, 822–842. [Google Scholar] [CrossRef]
- Duchatelet, L.; Marion, R.; Mallefet, J. A third luminous shark family: Confirmation of luminescence ability for Zameus Squamulosus (Squaliformes; Somniosidae). Photochem. Photobiol. 2021, 97, 739–744. [Google Scholar] [CrossRef]
- Claes, J.M.; Mallefet, J. Hormonal control of luminescence from lantern shark (Etmopterus spinax) photophores. J. Exp. Biol. 2009, 212, 3684–3692. [Google Scholar] [CrossRef]
- Duchatelet, L.; Delroisse, J.; Pinte, N.; Sato, K.; Ho, H.-C.; Mallefet, J. Adrenocorticotropic hormone and cyclic adenosine monophosphate are involved in the control of shark bioluminescence. Photochem. Photobiol. 2020, 96, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Claes, J.M.; Ho, H.-C.; Mallefet, J. Control of luminescence from pygmy shark (Squaliolus aliae) photophores. J. Exp. Biol. 2012, 215, 1691–1699. [Google Scholar] [CrossRef] [PubMed]
- Mallefet, J.; Stevens, D.W.; Duchatelet, L. Bioluminescence of the largest luminous vertebrate, the kitefin shark, Dalatias licha: First insights and comparative aspects. Front. Mar. Sci. 2021, 8, 153. [Google Scholar] [CrossRef]
- Ohshima, H. Some observations on the luminous organs of fishes. J. Coll. Sci. Imp. Univ. Tokyo 1911, 27, 1–25. [Google Scholar]
- Claes, J.M.; Mallefet, J. Early development of bioluminescence suggests camouflage by counter-illumination in the velvet belly lantern shark Etmopterus spinax (Squaloidea: Etmopteridae). J. Fish Biol. 2008, 73, 1337–1350. [Google Scholar] [CrossRef]
- Renwart, M.; Delroisse, J.; Flammang, P.; Claes, J.M.; Mallefet, J. Cytological changes during luminescence production in lanternshark (Etmopterus spinax Linnaeus, 1758) photophores. Zoomorphology 2015, 134, 107–116. [Google Scholar] [CrossRef]
- Duchatelet, L.; Sugihara, T.; Delroisse, J.; Koyanagi, M.; Rezsohazy, R.; Terakita, A.; Mallefet, J. From extraocular photoreception to pigment movement regulation: A new control mechanism of the lanternshark luminescence. Sci. Rep. 2020, 10, 10195. [Google Scholar] [CrossRef]
- Hubbs, C.L.; Iwai, T.; Matsubara, K. External and internal characters, horizontal and vertical distributions, luminescence, and food of the dwarf pelagic shark, Euprotomicrus bispinatus. Bull. Scripps Inst. Oceanogr. Univ. Calif. 1967, 10, 1–64. [Google Scholar]
- Seigel, J.A. Revision of the dalatiid shark genus Squaliolus: Anatomy, systematics, ecology. Copeia 1978, 1978, 602–614. [Google Scholar] [CrossRef]
- Delroisse, J.; Duchatelet, L.; Flammang, P.; Mallefet, J. Photophore distribution and enzymatic diversity within the photogenic integument of the cookie-cutter shark Isistius brasiliensis (Chondrichthyes: Dalatiidae). Front. Mar. Sci. 2021, 8, 627045. [Google Scholar] [CrossRef]
- Duchatelet, L.; Ho, H.-C.; Mallefet, J. Photophore morphogenesis and extraocular encephalopsin expression during the embryogenesis of smalleye pygmy shark (Squaliolus aliae). Diversity 2022, 14, 1100. [Google Scholar] [CrossRef]
- Claes, J.M.; Delroisse, J.; Grace, M.A.; Doosey, M.H.; Duchatelet, L.; Mallefet, J. Histological evidence for secretory bioluminescence from the pectoral pockets of the American pocket shark (Mollisquama Mississippiensis). Sci. Rep. 2020, 10, 18762. [Google Scholar] [CrossRef] [PubMed]
- Bernal, D.; Donley, J.M.; Shadwick, R.E.; Syme, D.A. Mammal-like muscles power swimming in a cold-water shark. Nature 2005, 437, 1349–1352. [Google Scholar] [CrossRef] [PubMed]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef]
- RStudio Team. RStudio: Integrated Development for R; RStudio: Boston, MA, USA, 2020; Available online: http://www.rstudio.com (accessed on 23 November 2022).
- Anctil, M. Ultrastructural correlates of luminescence in Porichthys photophores. I. Effects of spinal cord stimulation and exogenous noradrenaline. Rev. Can. Biol. 1979, 38, 67–80. [Google Scholar]
- Deheyn, D.; Mallefet, J.; Jangoux, M. Cytological changes during bioluminescence production in dissociated photocytes from the ophiuroid Amphipholis squamata (Echinodermata). Cell Tissue Res. 2000, 299, 115–128. [Google Scholar] [CrossRef]
- Kaskova, Z.M.; Tsarkova, A.S.; Yampolsky, I.V. 1001 Lights: Luciferins, Luciferases, Their Mechanisms of Action and Applications in Chemical Analysis, Biology and Medicine. Chem. Soc. Rev. 2016, 45, 6048–6077. [Google Scholar] [CrossRef]
- Delroisse, J.; Duchatelet, L.; Flammang, P.; Mallefet, J. Leaving the dark side? Insights into the evolution of luciferases. Front. Mar. Sci. 2021, 8, 673620. [Google Scholar] [CrossRef]
- Duchatelet, L.; Delroisse, J.; Mallefet, J. Bioluminescence in lanternsharks: Insights from hormone receptor localization. Gen. Comp. Endocrinol. 2020, 294, 113488. [Google Scholar] [CrossRef]
- Cavallaro, M.; Battaglia, P.; Guerrera, M.C.; Abbate, F.; Levanti, M.B.; Andaloro, F.; Germanà, A.; Laurà, R. New data on morphology and ultrastructure of skin photophores in the deep-sea squid Histioteuthis bonnellii (Férussac, 1834), Cephalopoda: Histioteuthidae. Acta Zool. 2017, 98, 271–277. [Google Scholar] [CrossRef]
- Claes, J.M.; Sato, K.; Mallefet, J. Morphology and control of photogenic structures in a rare dwarf pelagic lantern shark (Etmopterus splendidus). J. Exp. Mar. Biol. Ecol. 2011, 406, 1–5. [Google Scholar] [CrossRef]
- Claes, J.M.; Mallefet, J. Comparative Control of Luminescence in Sharks: New Insights from the Slendertail Lanternshark (Etmopterus molleri). J. Exp. Mar. Biol. Ecol. 2015, 467, 87–94. [Google Scholar] [CrossRef]
- Claes, J.M.; Mallefet, J. Bioluminescence of sharks: First synthesis. In Bioluminescence in Focus—A Collection of Illuminating Essays; Meyer-Rochow, V.B., Ed.; Research signpost: Kerala, India, 2009; pp. 51–65. [Google Scholar]
- Straube, N.; Iglésias, S.P.; Sellos, D.Y.; Kriwet, J.; Schliewen, U.K. Molecular phylogeny and node time estimation of bioluminescent lantern sharks (Elasmobranchii: Etmopteridae). Mol. Phylogenet. Evol. 2010, 56, 905–917. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duchatelet, L.; Nuyt, C.; Puozzo, N.; Mallefet, J.; Delroisse, J. Evolutionary Conservation of Photophore Ultrastructure in Sharks: The Case of a Dalatiid Squalomorph. Fishes 2023, 8, 87. https://doi.org/10.3390/fishes8020087
Duchatelet L, Nuyt C, Puozzo N, Mallefet J, Delroisse J. Evolutionary Conservation of Photophore Ultrastructure in Sharks: The Case of a Dalatiid Squalomorph. Fishes. 2023; 8(2):87. https://doi.org/10.3390/fishes8020087
Chicago/Turabian StyleDuchatelet, Laurent, Charlotte Nuyt, Nathan Puozzo, Jérôme Mallefet, and Jérôme Delroisse. 2023. "Evolutionary Conservation of Photophore Ultrastructure in Sharks: The Case of a Dalatiid Squalomorph" Fishes 8, no. 2: 87. https://doi.org/10.3390/fishes8020087
APA StyleDuchatelet, L., Nuyt, C., Puozzo, N., Mallefet, J., & Delroisse, J. (2023). Evolutionary Conservation of Photophore Ultrastructure in Sharks: The Case of a Dalatiid Squalomorph. Fishes, 8(2), 87. https://doi.org/10.3390/fishes8020087