
Effects of Stocking Density, Size, and External Stress on Growth and Welfare of African Catfish (Clarias gariepinus Burchell, 1822) in a Commercial RAS

 ,
,  , , and
, , and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Production System, Maintenance, and Water Quality
2.2. Fish Stocking, Feeding, and Growth
2.3. Sampling
2.4. External Stressor
2.5. RNA Isolation and Multiplex Quantitative PCR
2.6. Statistics
3. Results
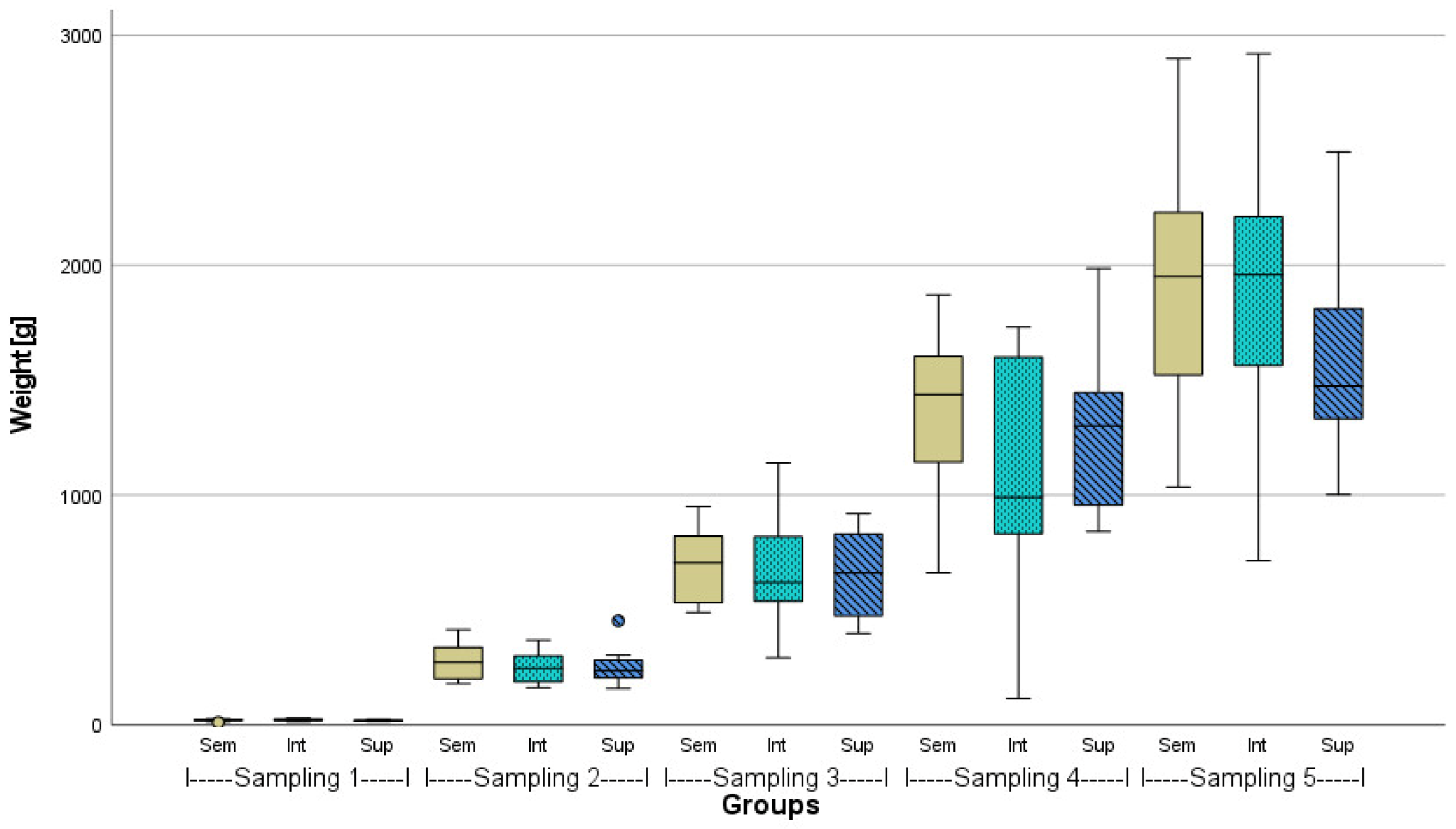
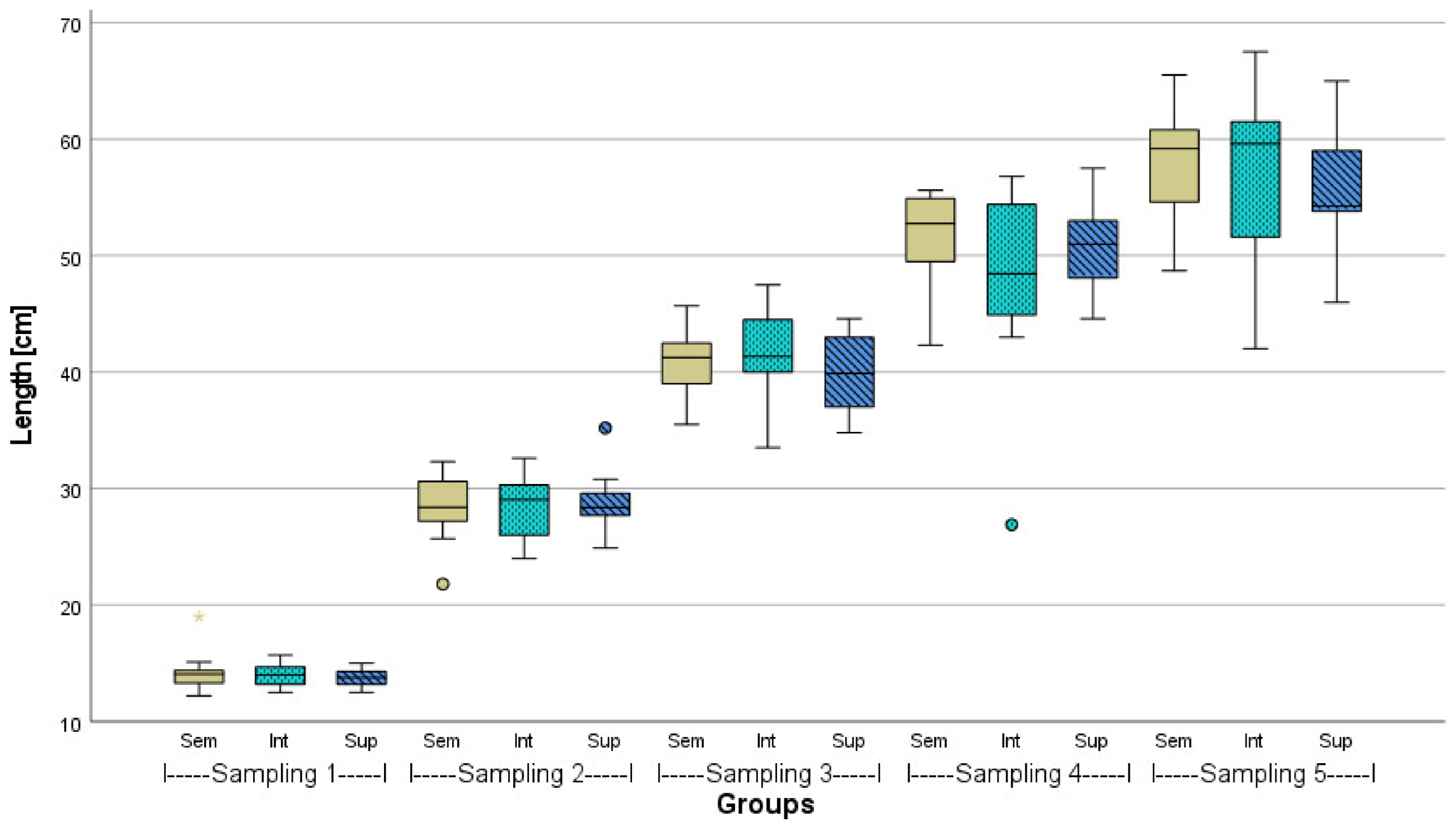
3.1. Fish Growth Performance
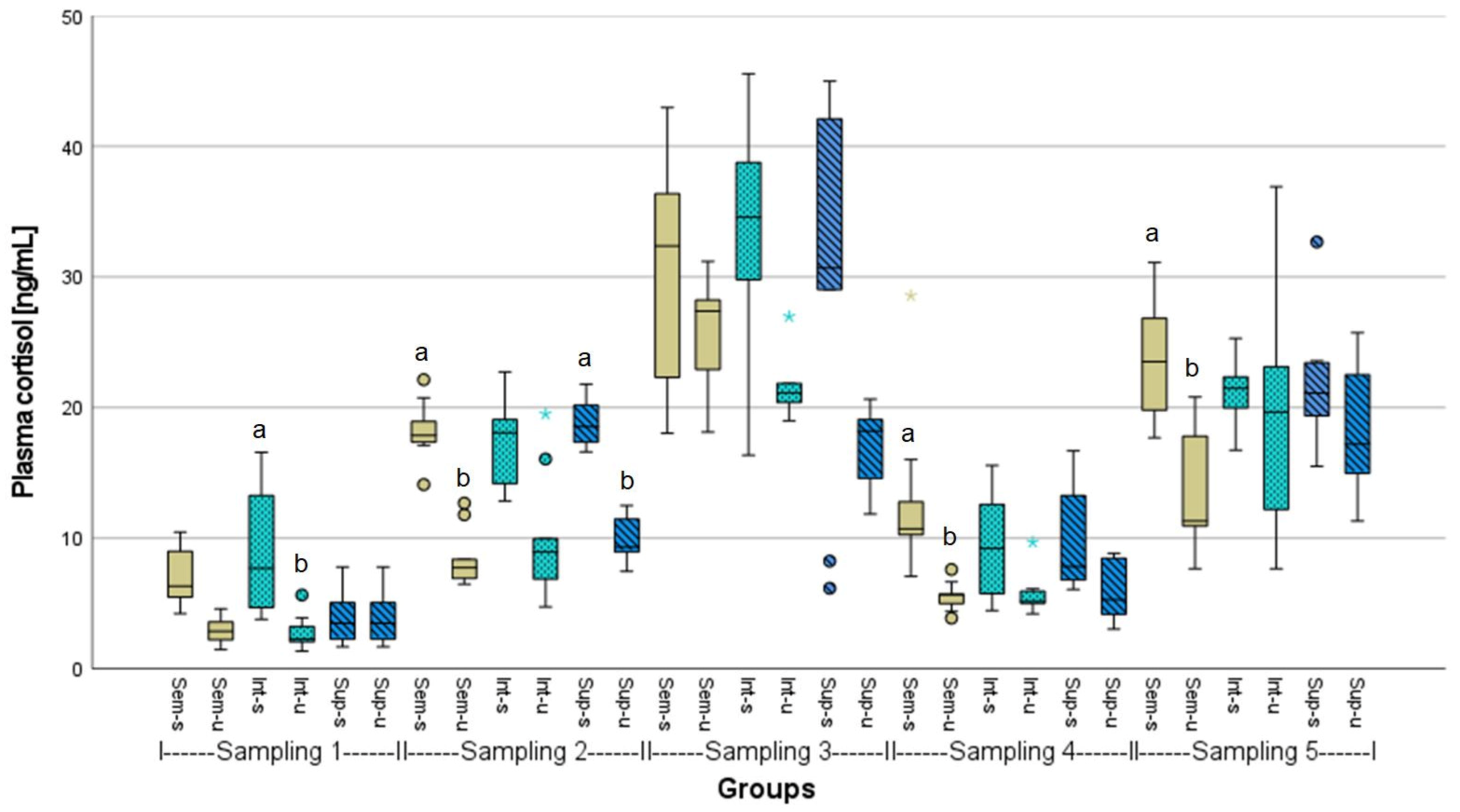
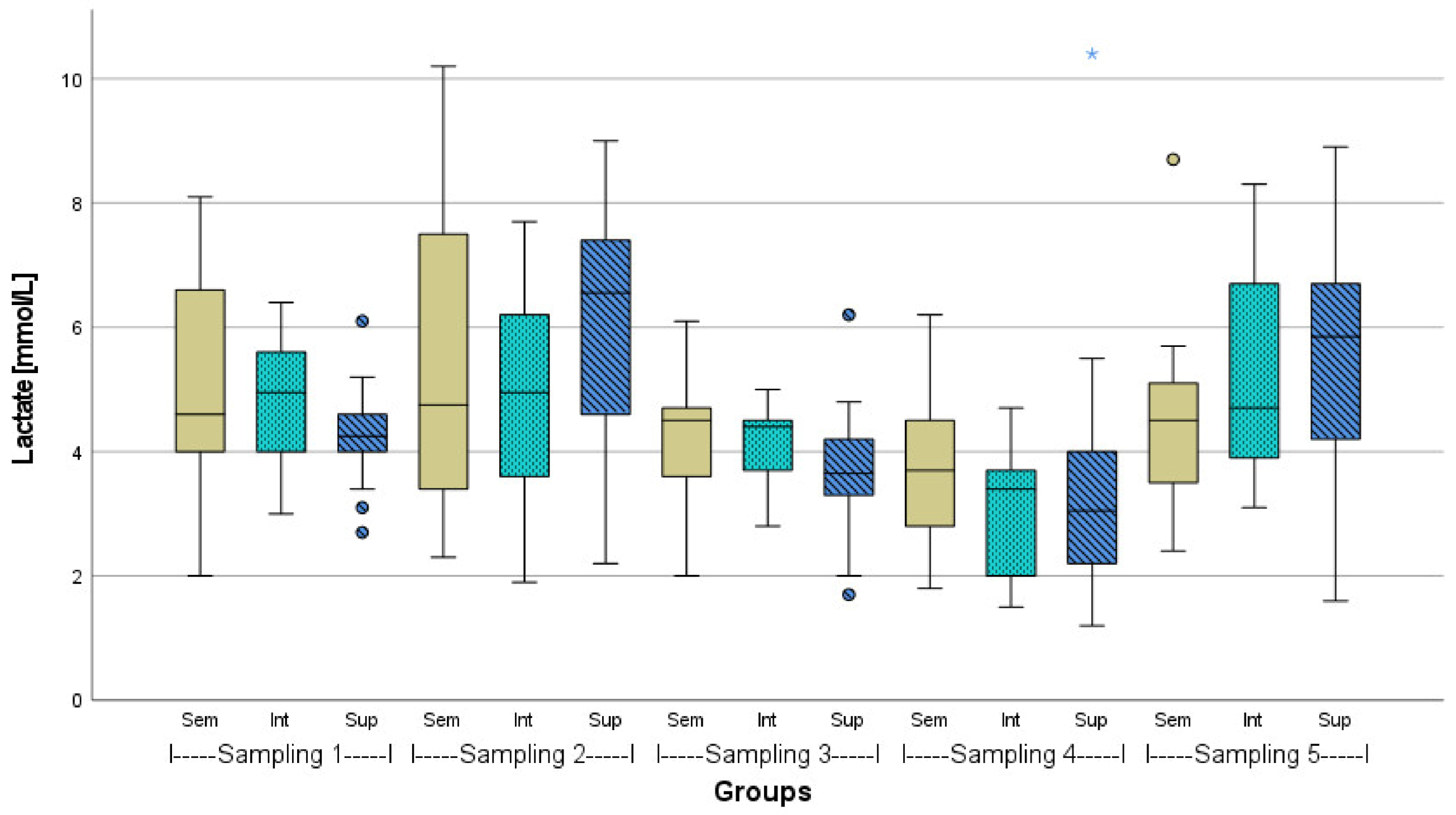
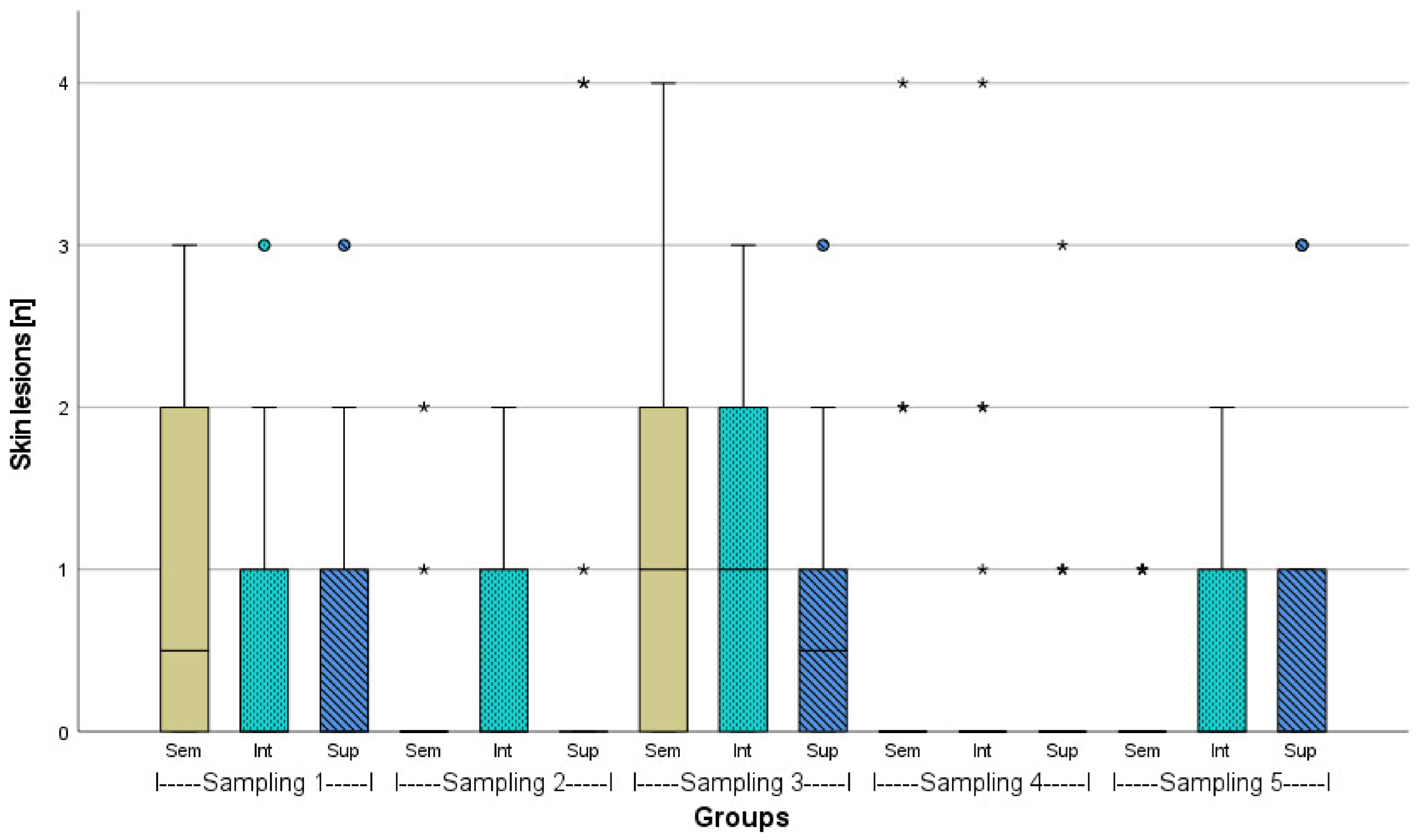
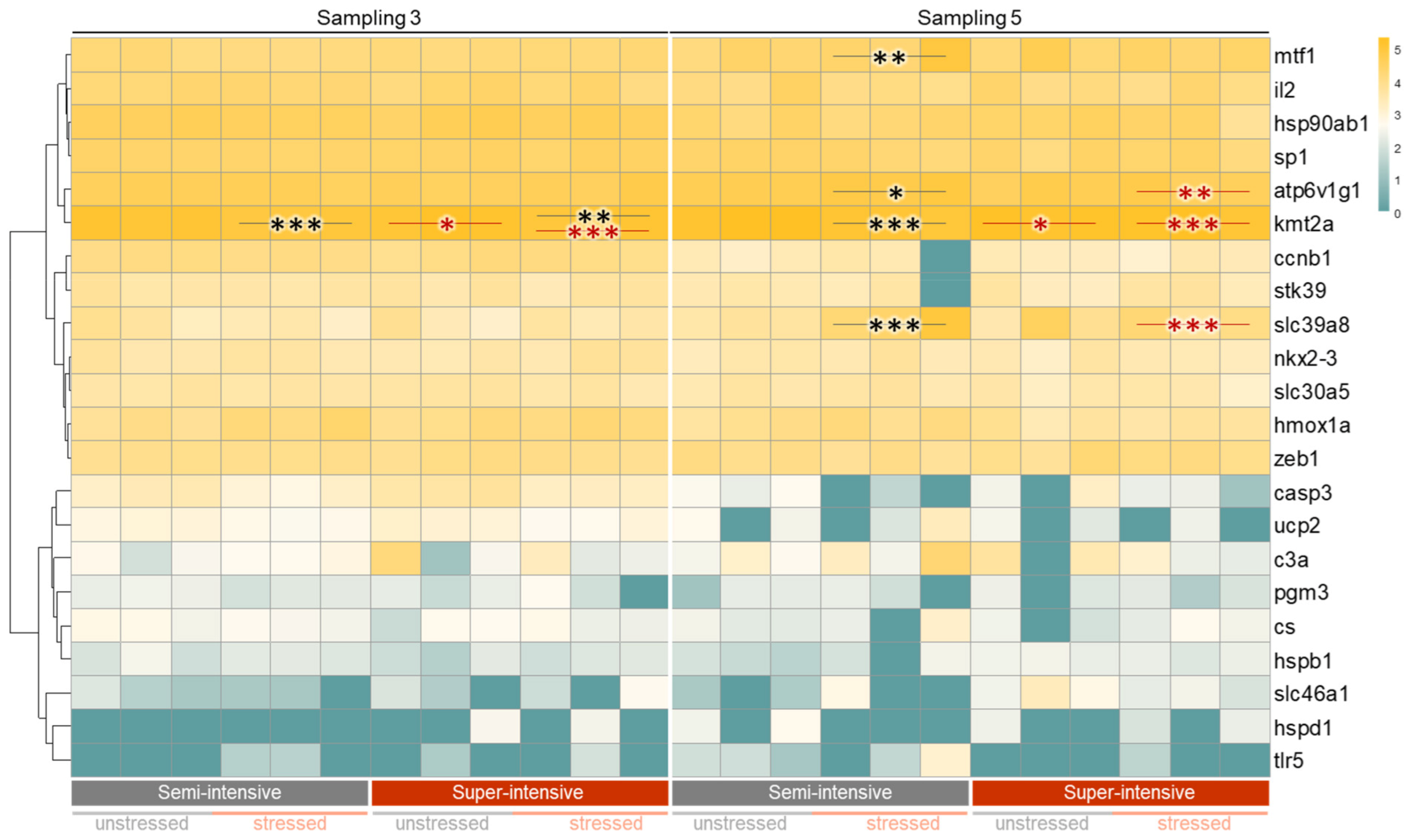
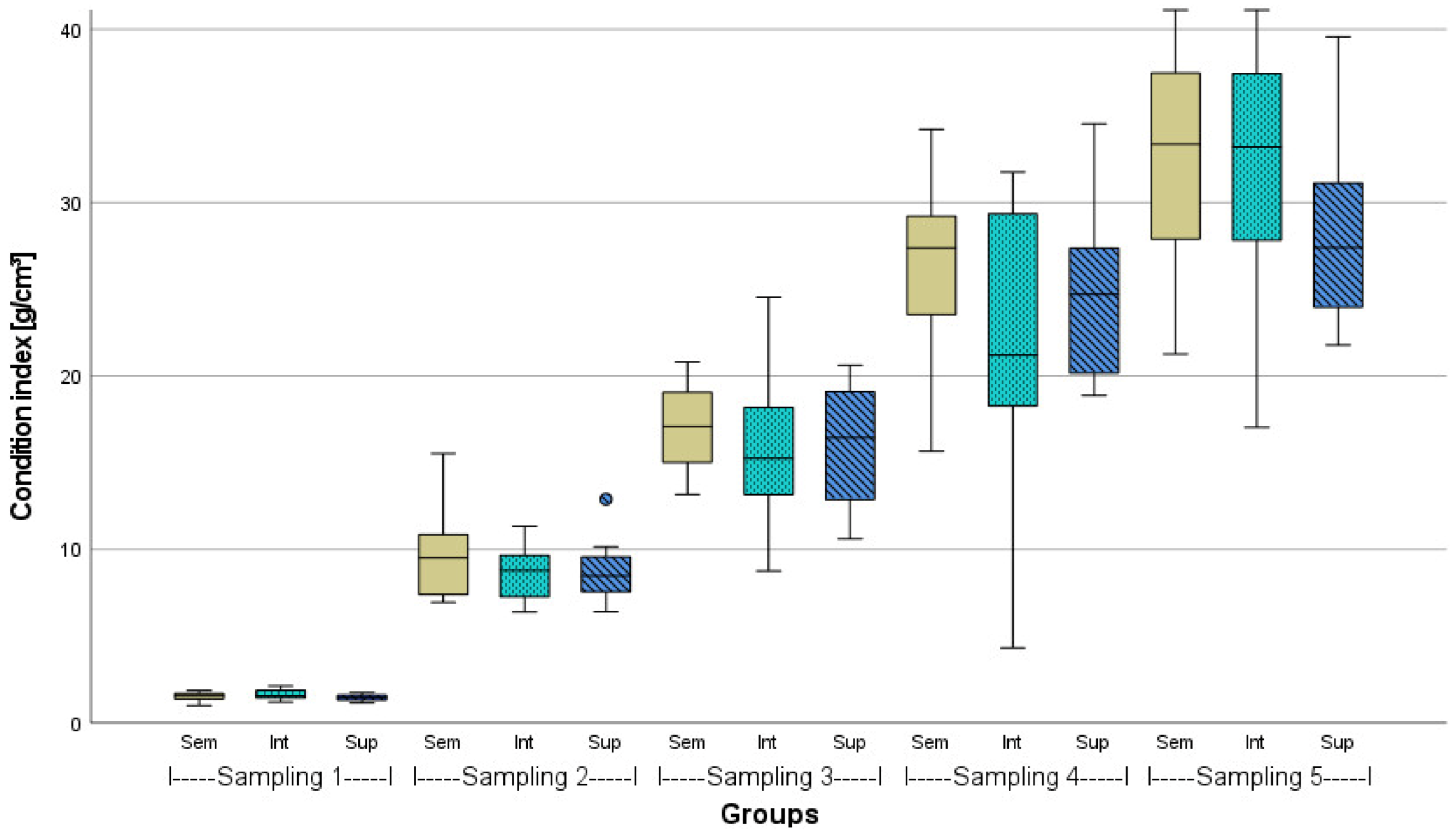
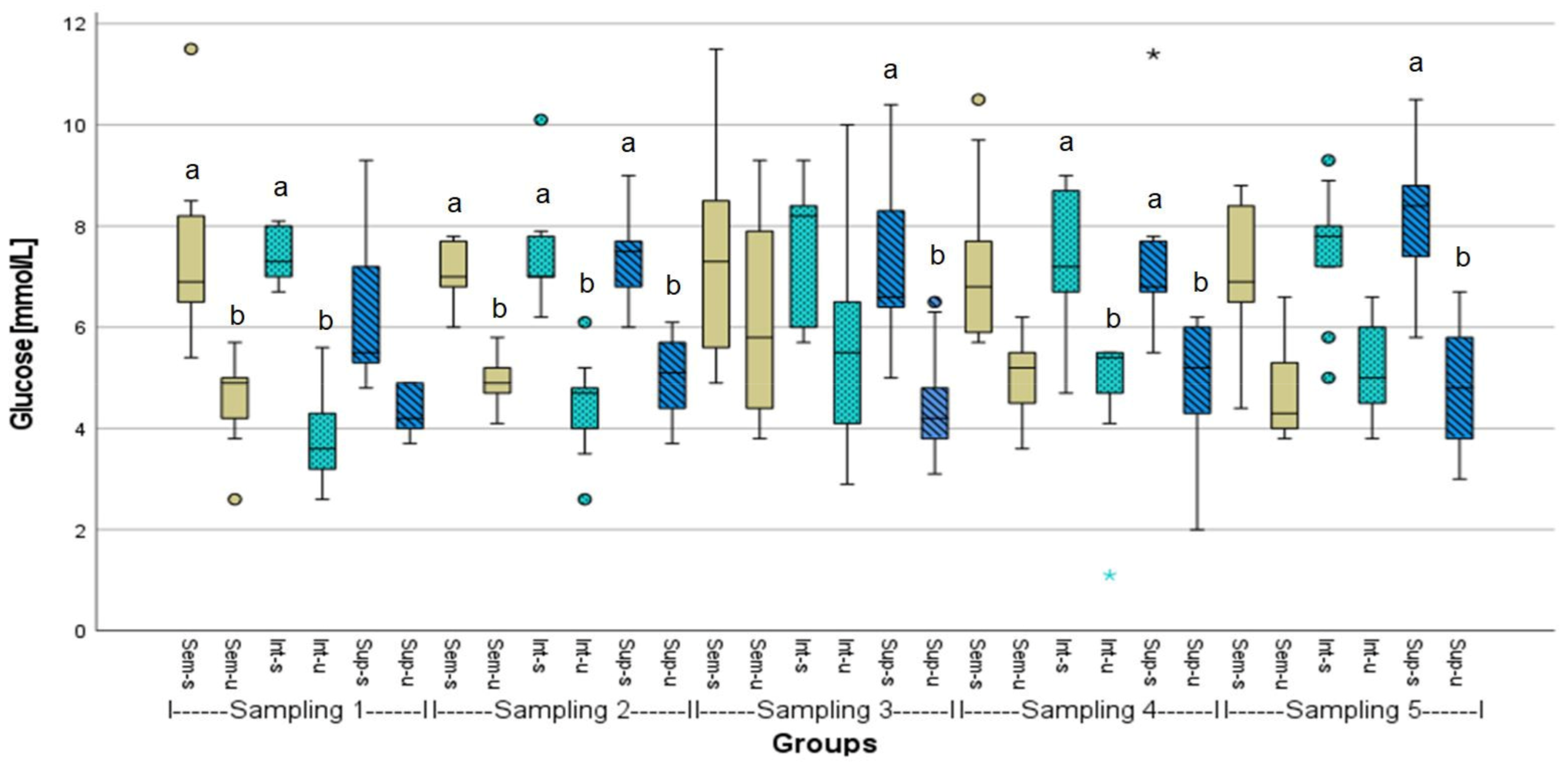
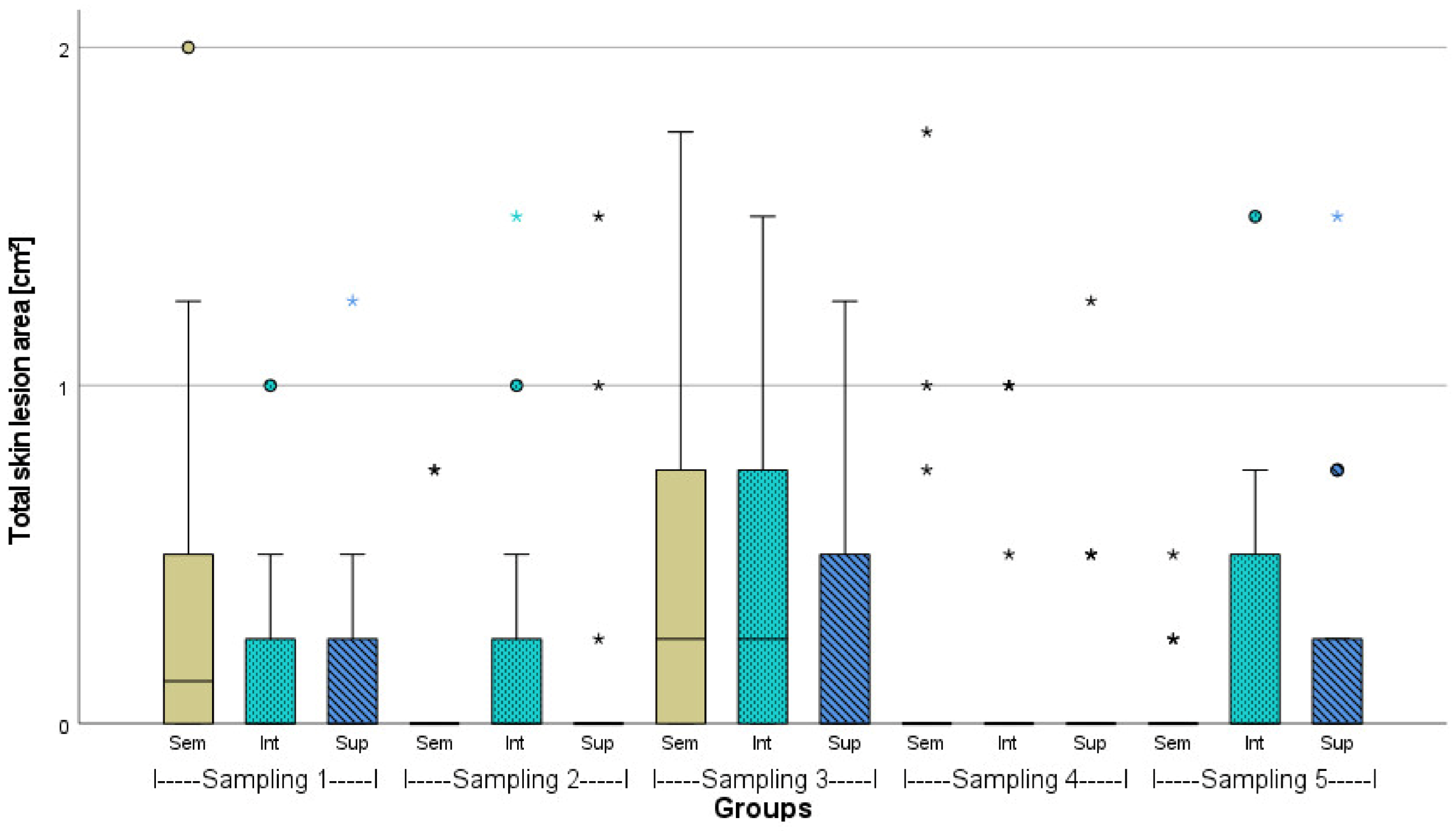
3.2. Evaluation of Fish Welfare
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Gene Product | Function | Sense Primer (5‘→3‘), Antisense Primer (5‘→3‘) | Source (Species; Accession Code) | Amplicon Length (bp) |
|---|---|---|---|---|---|
| Reference genes: | |||||
| rna18s | 18S ribosomal RNA | Structure of eukaryotic ribosomes | CTCTGCTGGACGATGGCTTAC, TCGATGAAGAACGCAGCCAGC | C. gariepinus; GQ465239 | 94 |
| actb | Actin-beta | Cell structure and motility, intercellular signalling | ACCACCACAGCCGAGAGAGAA, CTTCCAGCCATCTTTCCTTGGT | C. gariepinus; EU527191 | 204 |
| gapdh | Glyceraldehyde-3-phosphate dehydrogenase | Carbohydrate metabolism | TATGAAGCCCGCTGAGATCCC, GCCTCTTCTCACTTGCAGGGT | C. gariepinus; AF323693 | 106 |
| rpl | Ribosomal protein, large subunit | Structure of eukaryotic ribosomes | ACTAAATAGCAACTGATCCCTATC, GAATATCTGACCACTAAGATCCG | C. gariepinus; MW080924 | 134 |
| Target genes: | |||||
| atp6v1g1 | ATPase H+ transporting v1 subunit g1 | Intercellular Fe homeostasis | CGGAAAAACCGCCGCTTGAAG, GACCAAGGAAGCCGCGGCAC | P. hypophthalmus; XM_026922532 | 106 |
| c3a | Complement component 3, variant a | Bacteria opsonisation and destruction | ATGTCTTTCGATGTCACGGTTTAT, TCGAACCAAGAGTAACGGCATG | I. punctatus; XM_017457024 | 114 |
| casp3 | Caspase 3 | Apoptosis | CTCTTTATCATTCAGGCTTGTCG, GTACTCTACTGCTCCAGGTTATT | I. punctatus; XM_017473312 | 139 |
| ccnb1 | Cyclin b1 | Control of the G2/M transition phase of the cell cycle | TCAAAAATCGGAGAGGTTACAGC, TGCACTTTGCTCCCTCTCTGG | I. punctatus; NC_030443 | 103 |
| cs | Citrate synthase | Aerobic metabolism | GGTGGTGAAGTGTCCGATGAAA, GCTATGGGCATGCTGTCCTGA | I. punctatus; XM_017487510 | 94 |
| hmox1a | Heme oxygenase 1 | Cellular response to xenobiotic stimulus | GATTCTTCTGTGTTCCCTGTATG, CCATCTACTTCCCTCAGGAGC | I. punctatus; XM_017491622 | 104 |
| hsp90ab1 | Heat-shock protein 90 alpha family class b member 1 | Chaperone function, stress response | GAACATCAAGCTGGGCATCCAT, TTACTACATCACTGGTGAGAGCA | I. punctatus; XM_017456214 | 167 |
| hspb1 | Heat-shock protein family b (small) member 1 | Differentiation of cell types, stress response | ACAGGACAACTGGAAGGTGAAC, GATTATCGGAAACCATGAGGAGA | Clarias batrachus; KT359728 | 107 |
| hspd1 | Heat shock protein family d (hsp60) member 1 | Chaperone function, stress response | GCACGCTTGTCCTCAACAGGTT, AGACATGGCGATTGCTACTGGA | I. punctatus; XM_017469365 | 113 |
| il2 | Interleukin-2 | Activation and proliferation of lymphocytes | GTCGGCCTGGGAAAAAGCCAAT, TTATGTGTTTGCACCAGACAACG | I. punctatus; XM_017474923 | 162 |
| kmt2a | Lysine-specific methyltransferase 2a | Regulation of early development and hematopoiesis | ATTGGGTCGAAATCGTGCTGTAT, ATGATAAGTCTTCAGTGGCAGGT | I. punctatus; XM_017490460 | 121 |
| mtf1 | Metal regulatory transcription factor 1 | Catabolic regulation of cartilages | GTAGGAGGGCATTCAGGGAAC, AGTCAGAACGCTGCCCCCTC | I. punctatus; XM_017475296 | 146 |
| nkx2-3 | NK2 homeobox 3 | Cell differentiation | TACAGGACAACCTGGTGGAAAG, ACAACTCTTGGTTTCCTGCTCTT | I. punctatus; XM_017464595 | 119 |
| pgm3 | Phosphoglucomutase 3 | Carbohydrate metabolism | GACACAGGCAGGGCTGAATCT, CTTCGTACAGCACACTGTAACC | I. punctatus; XM_017494096 | 112 |
| slc30a5 | Solute carrier family 30, member 5 | Zinc transportation | AATAGTCACCAAAAGACAGTGGAT, CATCGTTGTGCTCGAACAACAG | I. punctatus; XM_017459891 | 134 |
| slc39a8 | Solute carrier family 39, member 8 alias ZIP8 | Cellular zinc uptake, protection from inflammation-related injury and death | TTTAACCTGATCTCAGCCATGTC, TATGTTCCCTGAGATGAATGCCA | I. punctatus; XM_017489708 | 151 |
| slc46a1 | Solute carrier family 46, member 1 | Folate transportation | AATGGCGACATGCACAAGGGTAT, AGAACAGCCTTGCCCCAGGG | I. punctatus; XM_017491375 | 129 |
| sp1 | SP1 transcription factor | Cell growth, apoptosis, differentiation and immune responses | AGCACAGCAGGTGATCAGGGA, GAGAAGCGTGCACATGTCCATA | I. punctatus; XM_017450095 | 119 |
| stk39 | Serine/threonine kinase 39 | Stress response | TGTAGTTGTTGCTGCTAACCTTC, AGATCCCTGACGAGGTGAAGC | I. punctatus; XM_017469076 | 116 |
| tlr5 | Toll-like receptor 5 | Detection of bacteria | GGCAGCATGGGAAAGGGAGTT, GTTAAGGCTCTGGATCTGTCCA | I. punctatus; NM_001200229 | 103 |
| ucp2 | Uncoupling protein 2 | Regulation of production of reactive oxygen species, function of mitochondria | GGCTCCAGATCCAAGGGGAGA, CCACGTAGTCTCTACAACGGG | I. punctatus; XM_017489367 | 131 |
| zeb1 | Zinc finger e-box binding homeobox 1 | Repression of interleukin-2 function | GCAGAGACCAGCGGCATGTAA, ATACGAGTGCCCCAACTGTAAAA | I. punctatus; XM_017483097 | 156 |



| Group Comparison | p-Value | Group Comparison | p-Value | Group Comparison | p-Value |
|---|---|---|---|---|---|
| Within a sampling | Within a sampling | Within a sampling | |||
| 2Sem-u-2Sem-s | 0.022 | 1Int-u-1Int-s | 0.047 | 2Sup-u-2Sup-s | 0.038 |
| 4Sem-u-4Sem-s | 0.043 | Between samplings | Between samplings | ||
| 5Sem-u-5Sem-s | 0.032 | 1Int-u-2Int-u | 0.021 | 1Sup-u-2Sup-u | 0.040 |
| Between samplings | 1Int-s-2Int-s | 0.026 | 1Sup-s-2Sup-s | 0.000 | |
| 1Sem-u-2Sem-u | 0.042 | 1Int-u-3Int-u | 0.000 | 1Sup-u-3Sup-u | 0.000 |
| 1Sem-s-2Sem-s | 0.006 | 1Int-s-3Int-s | 0.000 | 1Sup-s-3Sup-s | 0.000 |
| 1Sem-u-3Sem-u | 0.000 | 1Inti-u-5Int-u | 0.000 | 1Sup-s-4Sup-s | 0.049 |
| 1Sem-s-3Sem-s | 0.000 | 2Int-u-3Int-u | 0.003 | 1Sup-u-5Sup-u | 0.000 |
| 1Sem-u-5Sem-u | 0.001 | 2Int-u-5Int-u | 0.043 | 1Sup-s-5Sup-s | 0.000 |
| 1Sem-s-5Sem-s | 0.000 | 4Int-s-2Int-s | 0.048 | 4Sup-s-2Sup-s | 0.031 |
| 2Sem-u-3Sem-u | 0.000 | 4Int-u-3Int-u | 0.000 | 4Sup-u-3Sup-u | 0.002 |
| 4Sem-u-3Sem-u | 0.000 | 4Int-s-3Int-s | 0.000 | 4Sup-s-3Sup-s | 0.002 |
| 4Sem-s-3Sem-s | 0.004 | 4Int-u-5Int-u | 0.001 | 4Sup-u-5Sup-u | 0.001 |
| 4Sem-u-5Sem-u | 0.018 | 4Int-s-5Int-s | 0.004 | 4Sup-s-5Sup-s | 0.008 |
| 4Sem-s-5Sem-s | 0.013 | ||||
| 5Sem-u-3Sem-u | 0.021 |


References
- Montero, D.; Izquierdo, M.S.; Tort, L.; Robaina, L.; Vergara, J.M. High stocking density produces crowding stress altering some physiological and biochemical parameters in gilthead seabream, Sparus aurata, juveniles. Fish Physiol. Biochem. 1999, 20, 53–60. [Google Scholar] [CrossRef]
- Ellis, T.; North, B.; Scott, A.P.; Bromage, N.R.; Porter, M.; Gadd, D. The relationships between density and welfare in farmed rainbow trout. J. Fish Biol. 2002, 61, 493–531. [Google Scholar] [CrossRef]
- Carbonara, P.; Alfonso, S.; Gai, F.; Gasco, L.; Palmegiano, G.; Spedicato, M.T.; Zupa, W.; Lembo, G. Moderate stocking density does not influence the behavioural and physiological responses of rainbow trout (Oncorhynchus mykiss) in organic aquaculture. Aquac. Res. 2020, 51, 3007–3016. [Google Scholar] [CrossRef]
- Conte, F.S. Stress and the welfare of cultured fish. Appl. Anim. Behav. Sci. 2004, 86, 205–223. [Google Scholar] [CrossRef]
- Ashley, P.J. Fish welfare: Current issues in aquaculture. Appl. Anim. Behav. Sci. 2007, 104, 199–235. [Google Scholar] [CrossRef]
- Baldwin, L. The effects of stocking density on fish welfare. Plym. Stud. Sci. 2011, 4, 372–383. [Google Scholar]
- Oké, V.; Goosen, N.J. The effect of stocking density on profitability of African catfish (Clarias gariepinus) culture in extensive pond systems. Aquaculture 2019, 507, 385–392. [Google Scholar] [CrossRef]
- Ferreira, H.; Ramalho Ribeiro, A.; Dias, J.; Yúfera, M.; Arias, A.M.; Falcão, M.; Serpa, D.; Aires, T.; Pousão-Ferreira, P.; Cunha, M.E.; et al. Sustainable semi-intensive polyculture of seabream and sole in earthen ponds. Aquac. Eur. 2010, 35, 17–21. [Google Scholar]
- Lymbery, P. In too deep The welfare of intensively farmed fish. A report for Compassion in World Farming Trust. 2002. Available online: https://citeseerx.ist.psu.edu/document?repid=rep1&type=pdf&doi=0adba50d39fb67867286eed5f52c7c2ccb586d62 (accessed on 23 January 2023).
- Naylor, R.L.; Hardy, R.W.; Buschmann, A.H.; Bush, S.R.; Cao, L.; Klinger, D.H.; Little, D.C.; Lubchenco, J.; Shumway, S.E.; Troell, M. A 20-year retrospective review of global aquaculture. Nature 2021, 591, 551–563. [Google Scholar] [CrossRef]
- Ortuno, J.; Esteban, M.A.; Meseguer, J. Effects of short-term crowding stress on the gilthead seabream (Sparus aurata L.) innate immune response. Fish Shelf. Immunol. 2001, 11, 187–197. [Google Scholar] [CrossRef]
- North, B.P.; Turnbull, J.F.; Ellis, T.; Porter, M.J.; Migaud, H.; Bron, J.; Bromage, N.R. The impact of stocking density on the welfare of rainbow trout (Oncorhynchus mykiss). Aquaculture 2006, 255, 466–479. [Google Scholar] [CrossRef]
- Yarahmadi, P.; Miandare, H.K.; Fayaz, S.; Caipang, C.M.A. Increased stocking density causes changes in expression of selected stress-and immune-related genes, humoral innate immune parameters and stress responses of rainbow trout (Oncorhynchus mykiss). Fish Shelf. Immunol. 2016, 48, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Farhaduzzaman, A.M.; Hanif, A.; Khan, S.; Osman, M.H.; Shovon, N.H.; Rahman, M.K. Perfect Stocking Density Ensures Best Production and Economic Returns in Floating Cage Aquaculture System. J. Aquac. Res. Dev. 2020, 11, 1–7. [Google Scholar] [CrossRef]
- Debnath, D.; Das, B.K.; Yengkokpam, S.; Das, P.; Yadav, A.K.; Sharma, N.; Borah, S.; Ray, B.C.; Kakati, A.; Sarkar, U.K.; et al. Evaluating growth, production and economics of a new candidate species Labeo bata in cages: A regional model for table fish production in floodplain wetlands of North East India. Aquaculture 2022, 546, 737344. [Google Scholar] [CrossRef]
- Destatis, Statistisches Bundesamt. Land und Forstwirtschaft, Fischerei. Erzeugung in Aquakulturbetrieben 2012. Available online: https://www.statistischebibliothek.de/mir/servlets/MCRFileNodeServlet/DEHeft_derivate_00009985/2030460127004.pdf (accessed on 16 November 2022).
- Destatis, Statistisches Bundesamt. Land und Forstwirtschaft, Fischerei. Erzeugung in Aquakulturbetrieben 2020. Available online: https://www.statistischebibliothek.de/mir/servlets/MCRFileNodeServlet/DEHeft_derivate_00061752/2030460207004.pdf (accessed on 16 November 2022).
- Dauda, A.B.; Natrah, I.; Karim, M.; Kamarudin, M.S.; Bichi, A. African catfish aquaculture in Malaysia and Nigeria: Status, trends and prospects. Fish. Aquac. J. 2018, 9, 1–5. [Google Scholar] [CrossRef]
- Roques, J.A.; Schram, E.; Spanings, T.; van Schaik, T.; Abbink, W.; Boerrigter, J.; de Vries, P.; van de Vis, H.; Flik, G. The impact of elevated water nitrite concentration on physiology, growth and feed intake of African catfish Clarias gariepinus (Burchell 1822). Aquac. Res. 2015, 46, 1384–1395. [Google Scholar] [CrossRef]
- Păpuc, T.; Petrescu-Mag, I.V.; Gavriloaie, C.; Botha, M.; Kovacs, E.; Coroian, C.O. Swimming in the mud-a short review of environmental parameter ranges tolerated by Clarias gariepinus. Extreme Life Biospeol. Astrobiol. 2019, 11, 9–17. [Google Scholar]
- Belão, T.C.; Leite, C.A.C.; Florindo, L.H.; Kalinin, A.L.; Rantin, F.T. Cardiorespiratory responses to hypoxia in the African catfish, Clarias gariepinus (Burchell 1822), an air-breathing fish. J. Comp. Physiol. B 2011, 181, 905–916. [Google Scholar] [CrossRef] [PubMed]
- Van de Nieuwegiessen, P.G.; Boerlage, A.S.; Verreth, J.A.; Schrama, J.W. Assessing the effects of a chronic stressor, stocking density, on welfare indicators of juvenile African catfish, Clarias gariepinus Burchell. Appl. Anim. Behav. Sci. 2008, 115, 233–243. [Google Scholar] [CrossRef]
- Schram, E.; van der Heul, J.W.; Kamstra, A.; Verdegem, M.C.J. Stocking density-dependent growth of Dover sole (Solea solea). Aquaculture 2006, 252, 339–347. [Google Scholar] [CrossRef]
- Liu, B.; Liu, Y.; Sun, G. Effects of stocking density on growth performance and welfare-related physiological parameters of Atlantic salmon Salmo salar L. in recirculating aquaculture system. Aquac. Res. 2017, 48, 2133–2144. [Google Scholar] [CrossRef]
- Refaey, M.M.; Li, D.; Tian, X.; Zhang, Z.; Zhang, X.; Li, L.; Tang, R. High stocking density alters growth performance, blood biochemistry, intestinal histology, and muscle quality of channel catfish Ictalurus punctatus. Aquaculture 2018, 492, 73–81. [Google Scholar] [CrossRef]
- Long, L.; Zhang, H.; Ni, Q.; Liu, H.; Wu, F.; Wang, X. Effects of stocking density on growth, stress, and immune responses of juvenile Chinese sturgeon (Acipenser sinensis) in a recirculating aquaculture system. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2019, 219, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Oyarzún, R.; Paredes, R.; Saravia, J.; Morera, F.J.; Muñoz, J.L.P.; Ruiz-Jarabo, I.; Mancera, J.M.; Vargas-Chacoff, L. Stocking density affects the growth performance, intermediary metabolism, osmoregulation, and response to stress in Patagonian blennie Eleginops maclovinus. Aquaculture 2020, 515, 734565. [Google Scholar] [CrossRef]
- Van de Nieuwegiessen, P.G.; Olwo, J.; Khong, S.; Verreth, J.A.; Schrama, J.W. Effects of age and stocking density on the welfare of African catfish, Clarias gariepinus Burchell. Aquaculture 2009, 288, 69–75. [Google Scholar] [CrossRef]
- Wendelaar Bonga, S.E. The stress response in fish. Physiol. Rev. 1997, 77, 591–625. [Google Scholar] [CrossRef]
- Wenzel, L.C.; Berchtold, E.; Palm, H.W. Effects of stocking density and grading on behaviour, cannibalism and performance of African catfish (Clarias gariepinus) fry. Aquac. Rep. 2022, 27, 101400. [Google Scholar] [CrossRef]
- Palm, H.W.; Knaus, U.; Wasenitz, B.; Bischoff, A.A.; Strauch, S.M. Proportional up scaling of African catfish (Clarias gariepinus Burchell, 1822) commercial recirculating aquaculture systems disproportionally affects nutrient dynamics. Aquaculture 2018, 491, 155–168. [Google Scholar] [CrossRef]
- Martins, C.I.; Schrama, J.W.; Verreth, J.A. The effect of group composition on the welfare of African catfish (Clarias gariepinus). Appl. Anim. Behav. Sc. 2006, 97, 323–334. [Google Scholar] [CrossRef]
- Haufe, Handwerker auf Baustellen sind unterschiedlicher Lärmbelastung Ausgesetzt. Available online: https://www.haufe.de/arbeitsschutz/sicherheit/laermbelastung-auf-baustellen_96_156758.html (accessed on 15 February 2022).
- Hildebrand, M.-C.; Rebl, A.; Nguinkal, J.A.; Palm, H.W.; Baßmann, B. Effects of Fe-DTPA on Health and Welfare of the African Catfish Clarias gariepinus (Burchell, 1822). Water 2023, 15, 299. [Google Scholar] [CrossRef]
- Hecht, T. A review of on-farm feed management practices for North African catfish (Clarias gariepinus) in sub-Saharan Africa. In On-Farm Feeding and Feed Management in Aquaculture; FAO Fisheries and Aquaculture Technical Paper No., 583; Hasan, M.R., New, M.B., Eds.; FAO: Rome, Italy, 2013; pp. 463–479. [Google Scholar]
- Toko, I.; Fiogbe, E.D.; Koukpode, B.; Kestemont, P. Rearing of African catfish (Clarias gariepinus) and vundu catfish (Heterobranchus longifilis) in traditional fish ponds (whedos): Effect of stocking density on growth, production and body composition. Aquaculture 2007, 262, 65–72. [Google Scholar] [CrossRef]
- Dai, W.; Wang, X.; Guo, Y.; Wang, Q.; Ma, J. Growth performance, hematological and biochemical responses of African catfish (Clarias gariepinus) reared at different stocking densities. Afr. J. Agric. Res. 2011, 6, 6177–6182. [Google Scholar] [CrossRef]
- Wang, X.; Dai, W.; Xu, M.; Pan, B.; Li, X.; Chen, Y. Effects of stocking density on growth, nonspecific immune response, and antioxidant status in African catfish (Clarias gariepinus). Isr. J. Aquac. Bamidgeh 2013, 65, 1–6. [Google Scholar]
- Wang, X.M.; Gao, J.W.; Xu, M.; Mo, B.L.; Dai, W.; Chen, C.H. Responses of growth rates and growth hormone levels of African catfish (Clarias gariepinus) to stocking density. Iran. J. Fish. Sci. 2017, 16, 698–710. [Google Scholar]
- Hengsawat, K.; Ward, F.J.; Jaruratjamorn, P. The effect of stocking density on yield, growth and mortality of African catfish (Clarias gariepinus Burchell 1822) cultured in cages. Aquaculture 1997, 152, 67–76. [Google Scholar] [CrossRef]
- Hecht, T.; Appelbaum, S. Notes on the growth of Israeli sharptooth catfish (Clarias gariepinus) during the primary nursing phase. Aquaculture 1987, 63, 195–204. [Google Scholar] [CrossRef]
- Li, L.; Shen, Y.; Yang, W.; Xu, X.; Li, J. Effect of different stocking densities on fish growth performance: A meta-analysis. Aquaculture 2021, 544, 737152. [Google Scholar] [CrossRef]
- Akinwole, A.O.; Faturoti, E.O. Biological performance of African Catfish (Clarias gariepinus) cultured in recirculating system in Ibadan. Aquac. Eng. 2007, 36, 18–23. [Google Scholar] [CrossRef]
- Boerrigter, J.G.; van den Bos, R.; van de Vis, H.; Spanings, T.; Flik, G. Effects of density, PVC-tubes and feeding time on growth, stress and aggression in African catfish (Clarias gariepinus). Aquac. Res. 2016, 47, 2553–2568. [Google Scholar] [CrossRef]
- Martins, C.I.; Schrama, J.W.; Verreth, J.A. The relationship between individual differences in feed efficiency and stress response in African catfish Clarias gariepinus. Aquaculture 2006, 256, 588–595. [Google Scholar] [CrossRef]
- Manuel, R.; Boerrigter, J.; Roques, J.; van der Heul, J.; van den Bos, R.; Flik, G.; van de Vis, H. Stress in African catfish (Clarias gariepinus) following overland transportation. Fish Physiol. Biochem. 2014, 40, 33–44. [Google Scholar] [CrossRef]
- Polakof, S.; Panserat, S.; Soengas, J.L.; Moon, T.W. Glucose metabolism in fish: A review. J. Comp. Physiol. B 2012, 182, 1015–1045. [Google Scholar] [CrossRef] [PubMed]
- Baßmann, B.; Brenner, M.; Palm, H.W. Stress and welfare of African catfish (Clarias gariepinus Burchell, 1822) in a coupled aquaponic system. Water 2017, 9, 504. [Google Scholar] [CrossRef]
- Baßmann, B.; Harbach, H.; Weißbach, S.; Palm, H.W. Effect of plant density in coupled aquaponics on the welfare status of African catfish, Clarias gariepinus. J. World Aquac. Soc. 2020, 51, 183–199. [Google Scholar] [CrossRef]
- Palm, H.W.; Berchtold, E.; Gille, B.; Knaus, U.; Wenzel, L.C.; Baßmann, B. Growth and Welfare of African Catfish (Clarias gariepinus Burchell, 1822) under Dietary Supplementation with Mixed-Layer Clay Mineral Montmorillonite-Illite/Muscovite in Commercial Aquaculture. Aquac. J. 2022, 2, 227–245. [Google Scholar] [CrossRef]
- Huntingford, F.A.; Adams, C.; Braithwaite, V.A.; Kadri, S.; Pottinger, T.G.; Sandøe, P.; Turnbull, J.F. Current issues in fish welfare. J. Fish Biol. 2006, 68, 332–372. [Google Scholar] [CrossRef]
- Kaiser, H.; Weyl, O.; Hecht, T. Observations on agonistic behaviour of Clarias gariepinus larvae and juveniles under different densities and feeding frequencies in a controlled environment. J. Appl. Ichthyol. 1995, 11, 25–36. [Google Scholar] [CrossRef]
- Günther, V.; Lindert, U.; Schaffner, W. The taste of heavy metals: Gene regulation by MTF-1. Biochim. Biophys. Acta (BBA) Mol. Cell Res. 2012, 1823, 1416–1425. [Google Scholar] [CrossRef]
- Jeong, J.; Eide, D.J. The SLC39 family of zinc transporters. Mol. Asp. Med. 2013, 34, 612–619. [Google Scholar] [CrossRef]
- Wu, D.M.; Liu, T.; Deng, S.H.; Han, R.; Xu, Y. SLC39A4 expression is associated with enhanced cell migration, cisplatin resistance, and poor survival in non-small cell lung cancer. Sci. Rep. 2017, 7, 7211. [Google Scholar] [CrossRef]
- Swaleh, S.B.; Banday, U.Z.; Asadi, M.A.; Usmani, N. Biochemical profile and gene expression of Clarias gariepinus as a signature of heavy metal stress. Environ. Pollut. 2020, 264, 114693. [Google Scholar] [CrossRef] [PubMed]
- Kari, Z.A.; Kabir, M.A.; Dawood, M.A.; Razab, M.K.A.A.; Ariff, N.S.N.A.; Sarkar, T.; Pati, S.; Edinur, H.A.; Mat, K.; Ismail, T.A.; et al. Effect of fish meal substitution with fermented soy pulp on growth performance, digestive enzyme, amino acid profile, and immune-related gene expression of African catfish (Clarias gariepinus). Aquaculture 2022, 546, 737418. [Google Scholar] [CrossRef]
- Coppola, C.L.; Enns, R.M.; Grandin, T. Noise in the animal shelter environment: Building design and the effects of daily noise exposure. J. Appl. Anim. Welf. Sci. 2006, 9, 1–7. [Google Scholar] [CrossRef]
- Kunc, H.P.; Schmidt, R. The effects of anthropogenic noise on animals: A meta-analysis. Biol. Lett. 2019, 15, 20190649. [Google Scholar] [CrossRef] [PubMed]
- Bart, A.N.; Clark, J.; Young, J.; Zohar, Y. Underwater ambient noise measurements in aquaculture systems: A survey. Aquac. Eng. 2001, 25, 99–110. [Google Scholar] [CrossRef]
- Wysocki, L.E.; Dittami, J.P.; Ladich, F. Ship noise and cortisol secretion in European freshwater fishes. Biol. Conserv. 2006, 128, 501–508. [Google Scholar] [CrossRef]
- Hang, S.; Zhao, J.; Ji, B.; Li, H.; Zhang, Y.; Peng, Z.; Zhou, F.; Ding, X.; Ye, Z. Impact of underwater noise on the growth, physiology and behavior of Micropterus salmoides in industrial recirculating aquaculture systems. Environ. Pollut. 2021, 291, 118152. [Google Scholar] [CrossRef] [PubMed]





| Mean ± SD | Min/Max (T0–T1) | Min/Max (T1–T2) | Min/Max (T2–T3) | Min/Max (T3–T4) | Min/Max (T4–T5) | |
|---|---|---|---|---|---|---|
| Temperature [°C] | 26.1 ± 1.6 | 24.2/26.1 | 23.8/26.7 | 19.0/27.5 | 23.6/28.0 | 25.7/28.6 |
| O2 [mg L−1] | 6.0 ± 0.9 | 7.1/8.1 | 6.1/8.3 | 4.1/8.7 | 4.7/6.8 | 4.4/6.2 |
| O2 [%] | 74.0 ± 10.9 | 90.4/96.5 | 74.4/103.5 | 50.8/97.1 | 60.1/82.6 | 53.1/80.2 |
| pH | 6.7 ± 0.9 | 7.7/8.2 | 3.7/8.1 | 6.1/7.7 | 6.1/7.8 | 5.0/7.2 |
| EC [µS cm−1] | 1382 ± 451 | 762/883 | 906/1610 | 672/1520 | 814/2230 | 1371/2480 |
| Salinity [‰] | 0.7 ± 0.2 | 0.4/0.4 | 0.4/0.8 | 0.4/0.7 | 0.4/1.1 | 0.7/1.2 |
| RedOx [mV] | 160.1 ± 31.7 | 173.6/194.5 | 158.8/276.2 | 120.2/192.0 | 105.7/191.2 | 60.8/174.3 |
| NH4 * [mg L−1] | 13.60 ± 3.88 | 9.62/18.99 | 9.03/14.73 | 8.80/13.41 | 8.48/16.50 | 8.78/13.21 |
| NO2 * [mg L−1] | 0.19 ± 0.17 | 0.03/0.03 | 0.09/0.49 | 0.06/0.36 | 0.05/0.35 | 0.07/0.37 |
| NO3 * [mg L−1] | 475 ± 154 | 591/606 | 511/518 | 339/346 | 325/750 | 344/349 |
| Intended Stocking Density | n Fish Tank−1 | Mean Weight Fish−1 [g] ± SD | Mean Lengths Fish−1 [cm] ± SD | de facto Density Tank−1 [kg m−3] | |
|---|---|---|---|---|---|
| Before stocking | Semi-intensive (0.84 kg m−3) | 88 88 88 | 12.5 ± 1.9 | 12.1 ± 0.7a | 0.87 0.87 0.87 |
| Intensive (1.68 kg m−3) | 176 176 176 | 12.4 ± 1.9 | 11.6 ± 0.8b | 1.72 1.72 1.72 | |
| Super-intensive (3.35 kg m−3) | 352 352 352 | 12.2 ± 1.6 | 12.0 ± 0.6a | 3.41 3.41 3.41 | |
| After 23 weeks | Semi-intensive (100 kg m−3) | 55 73 73 | 1830.5 ± 596.7 a | 57.3 ± 6.3 | 89.15 * 106.40 106.85 |
| Intensive (200 kg m−3) | 161 161 163 | 1664.8 ± 588.5 a,b | 55.6 ± 7.8 | 230.62 212.34 214.96 | |
| Super-Intensive (400 kg m−3) | 284 313 309 | 1615.4 ± 451.7 b | 56.1 ± 5.1 | 364.52 * 401.13 396.60 * |
| Stocking Density | T0–T1 | T1–T2 | T2–T3 | T3–T4 | T4–T5 | T0–T5 (in Total) | |
|---|---|---|---|---|---|---|---|
| Semi-intensive (100 kg m−3) | FCR SGR [% d−1] | 0.7 7.0 | 0.7 6.0 | 0.8 2.2 | 0.7 1.7 | 1.1 0.9 | 0.77 3.0 |
| Intensive (200 kg m−3) | FCR SGR [% d−1] | 0.6 7.8 | 0.7 5.8 | 0.7 2.4 | 0.9 1.4 | 0.8 1.2 | 0.75 3.0 |
| Super-intensive (400 kg m−3) | FCR SGR [% d−1] | 0.7 6.8 | 0.7 6.0 | 0.8 2.3 | 0.8 1.6 | 1.2 0.7 | 0.83 2.9 |
| Stocking Density | Mortality Tank−1 [%] | Mean [%] ± SD |
|---|---|---|
| Semi-intensive (100 kg m−3) | 2.59 | 7.12 ± 3.92 |
| 9.56 | ||
| 9.20 | ||
| Intensive (200 kg m−3) | 3.89 | 3.79 ± 0.59 |
| 4.32 | ||
| 3.16 | ||
| Super-Intensive (400 kg m−3) | 14.62 | 9.24 ± 4.80 |
| 7.71 | ||
| 5.40 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baßmann, B.; Hahn, L.; Rebl, A.; Wenzel, L.C.; Hildebrand, M.-C.; Verleih, M.; Palm, H.W. Effects of Stocking Density, Size, and External Stress on Growth and Welfare of African Catfish (Clarias gariepinus Burchell, 1822) in a Commercial RAS. Fishes 2023, 8, 74. https://doi.org/10.3390/fishes8020074
Baßmann B, Hahn L, Rebl A, Wenzel LC, Hildebrand M-C, Verleih M, Palm HW. Effects of Stocking Density, Size, and External Stress on Growth and Welfare of African Catfish (Clarias gariepinus Burchell, 1822) in a Commercial RAS. Fishes. 2023; 8(2):74. https://doi.org/10.3390/fishes8020074
Chicago/Turabian StyleBaßmann, Björn, Lisa Hahn, Alexander Rebl, Lisa Carolina Wenzel, Marc-Christopher Hildebrand, Marieke Verleih, and Harry Wilhelm Palm. 2023. "Effects of Stocking Density, Size, and External Stress on Growth and Welfare of African Catfish (Clarias gariepinus Burchell, 1822) in a Commercial RAS" Fishes 8, no. 2: 74. https://doi.org/10.3390/fishes8020074
APA StyleBaßmann, B., Hahn, L., Rebl, A., Wenzel, L. C., Hildebrand, M.-C., Verleih, M., & Palm, H. W. (2023). Effects of Stocking Density, Size, and External Stress on Growth and Welfare of African Catfish (Clarias gariepinus Burchell, 1822) in a Commercial RAS. Fishes, 8(2), 74. https://doi.org/10.3390/fishes8020074