
Identification and Characterization of the Stimulator of Interferon Genes (STING) in Chinese Giant Salamander Andrias davidianus


Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals, Cell, and Virus
2.2. Cloning and Sequence Analysis of AdSTING
2.3. Expression Profiles Analyses of AdSTING
2.4. Plasmid Construction
2.5. Prokaryotic Expression, Protein Purification and Antibody Preparation
2.6. Western Blot Analysis
2.7. Fluorescence Microscopy Observation
2.8. Luciferase Reporter Assay
2.9. Effects of AdSTING Overexpression on ADRV Infection
3. Results
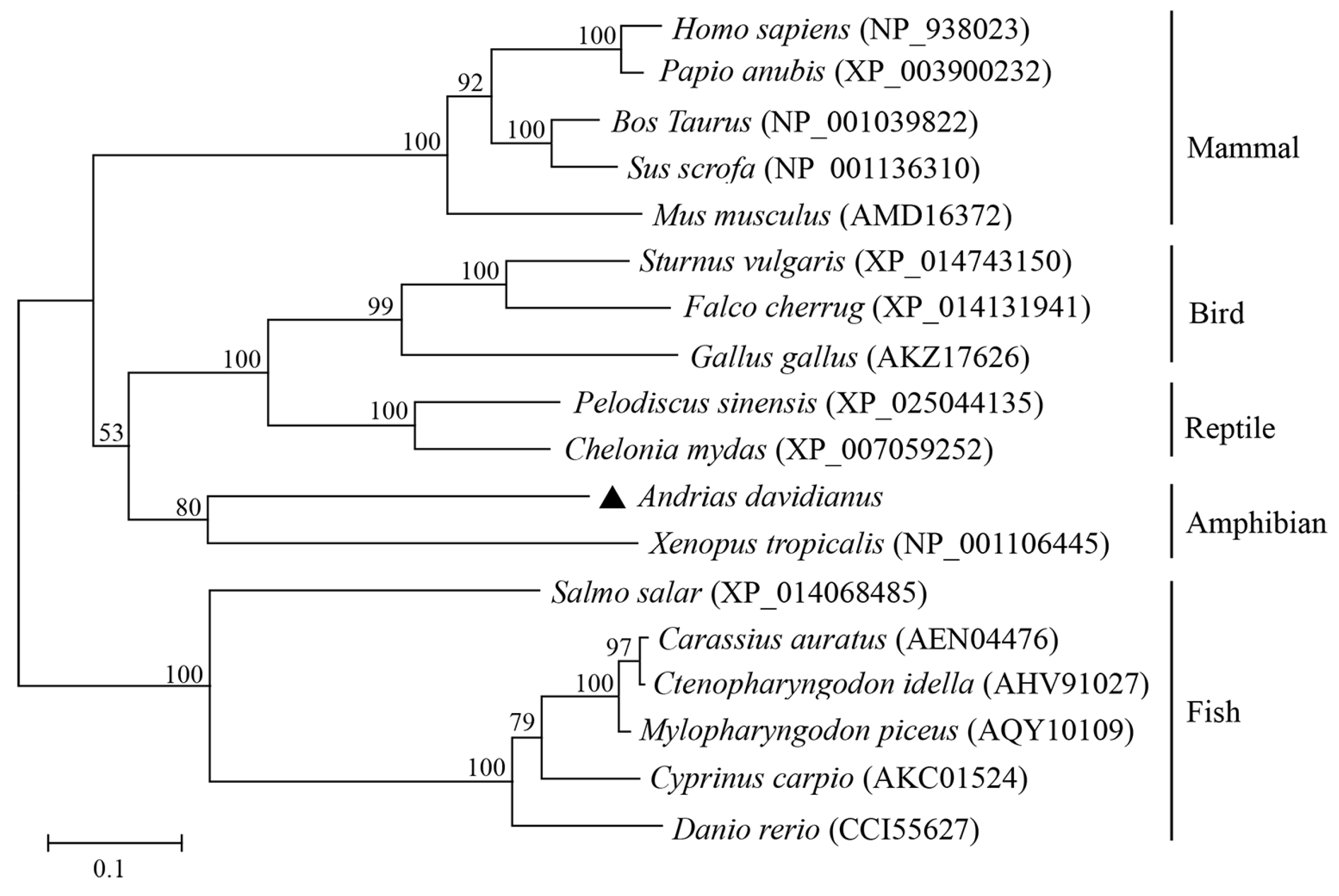
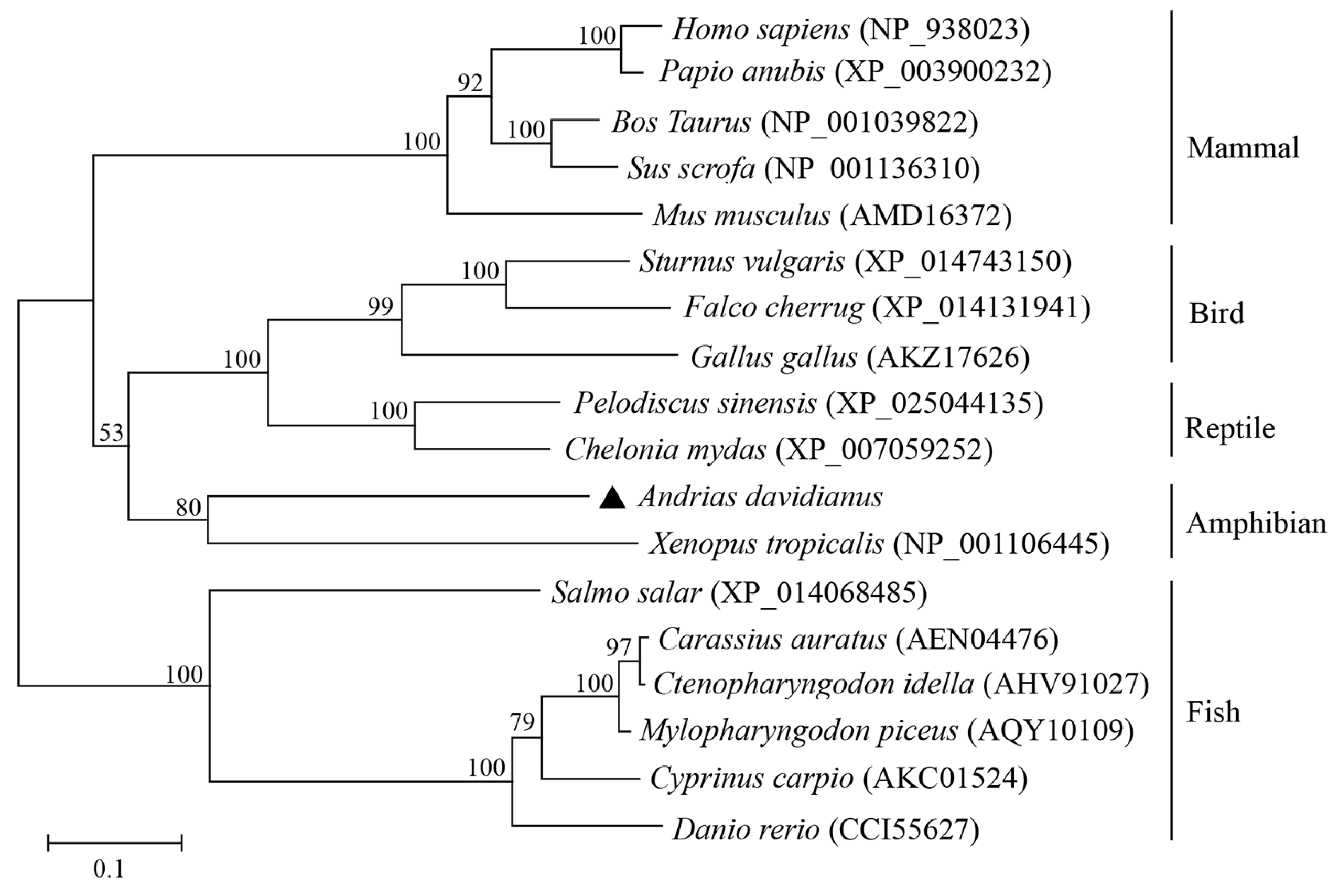
3.1. Identification and Sequence Characterization of AdSTING
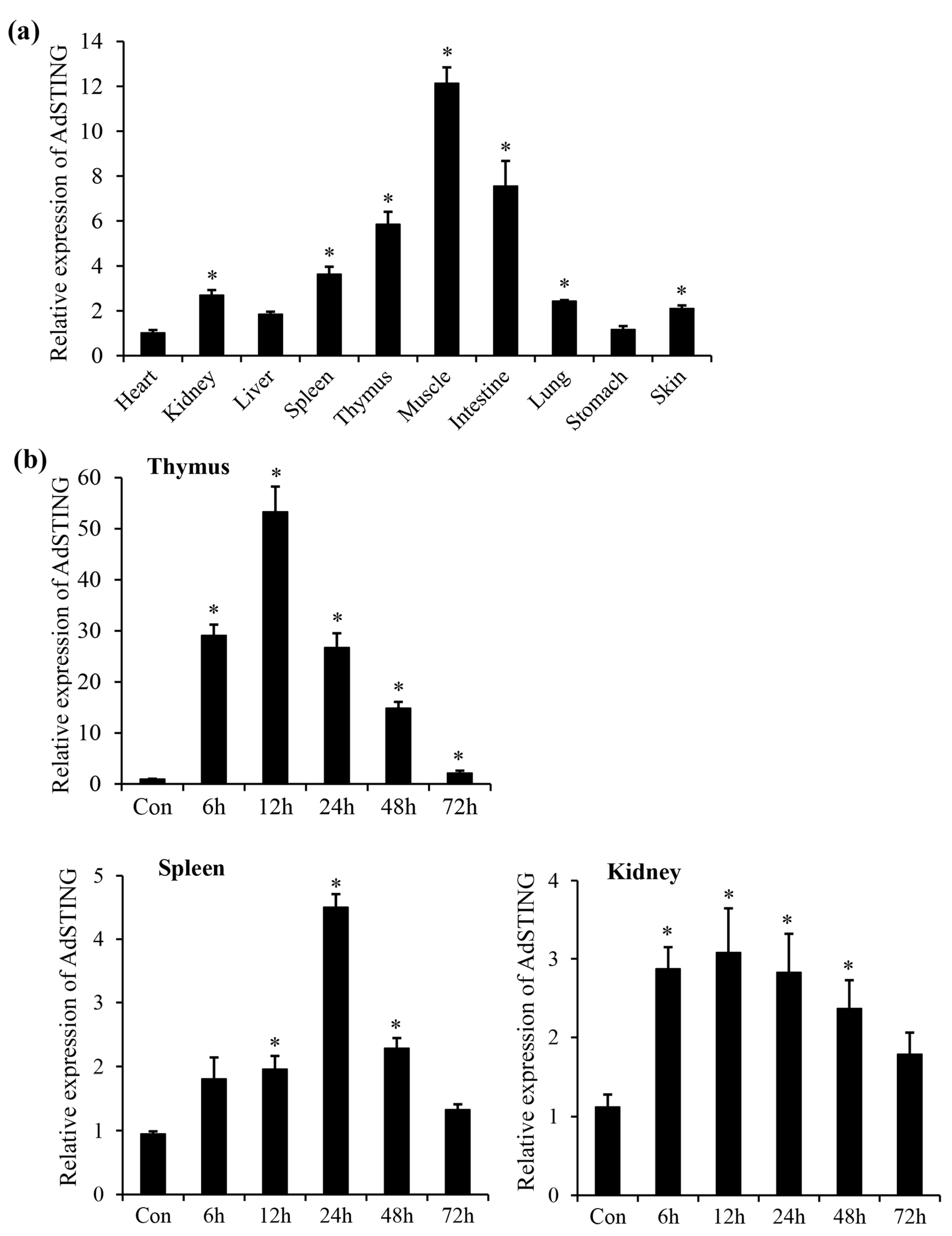
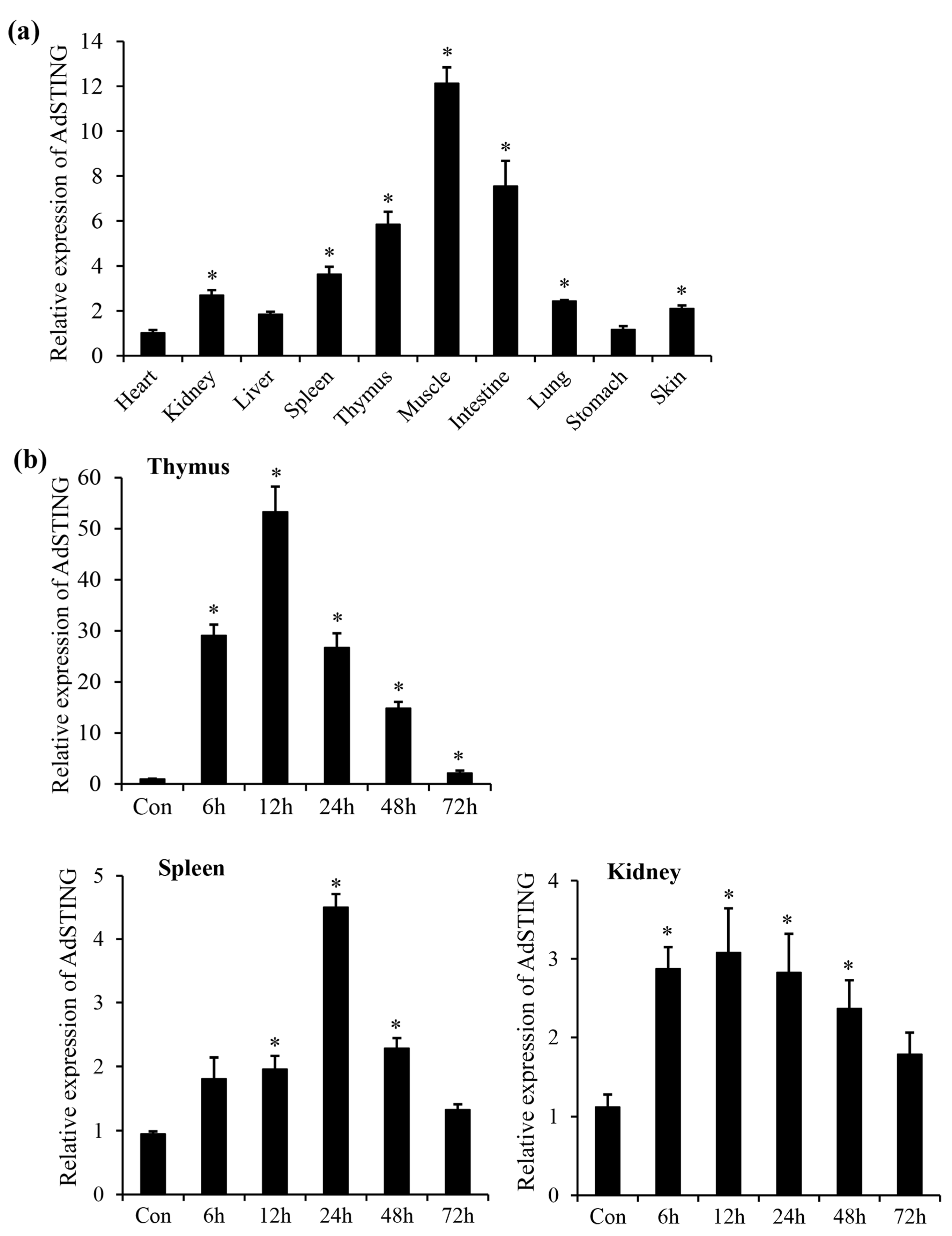
3.2. Tissue Expression Profile of AdSTING
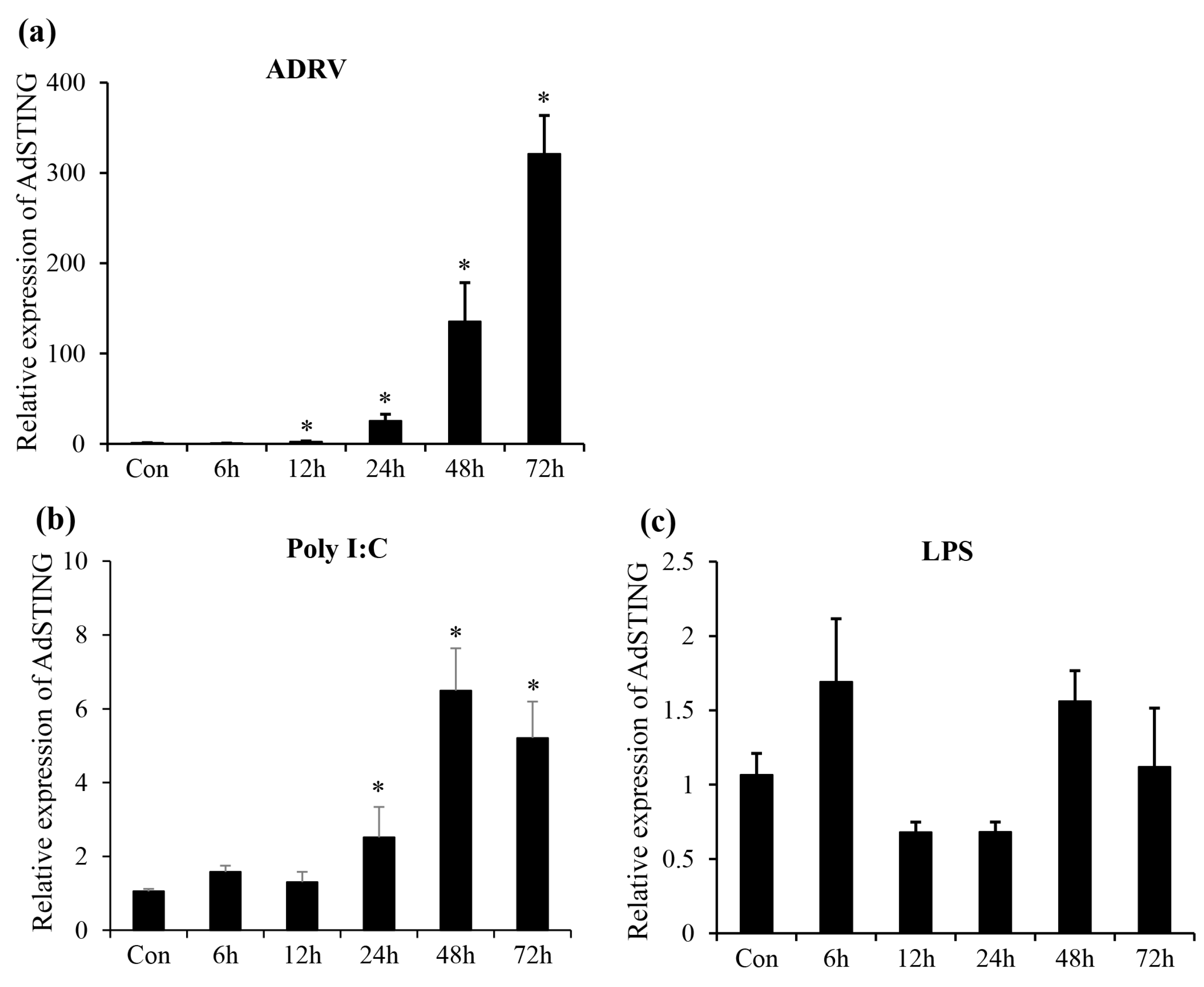
3.3. Expression Profile of AdSTING after In Vitro Stimulation
3.4. Expression and Confirmation of AdSTING
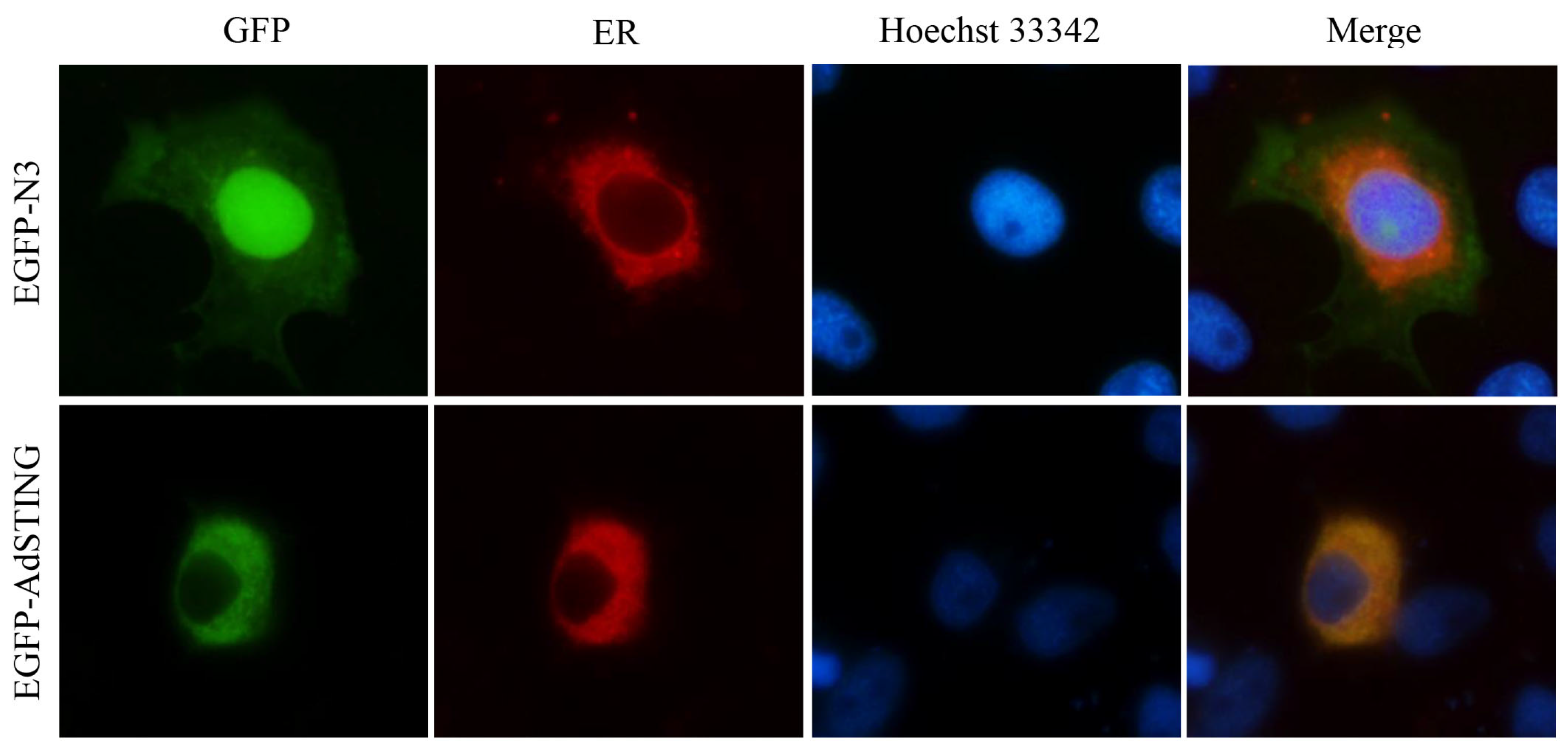
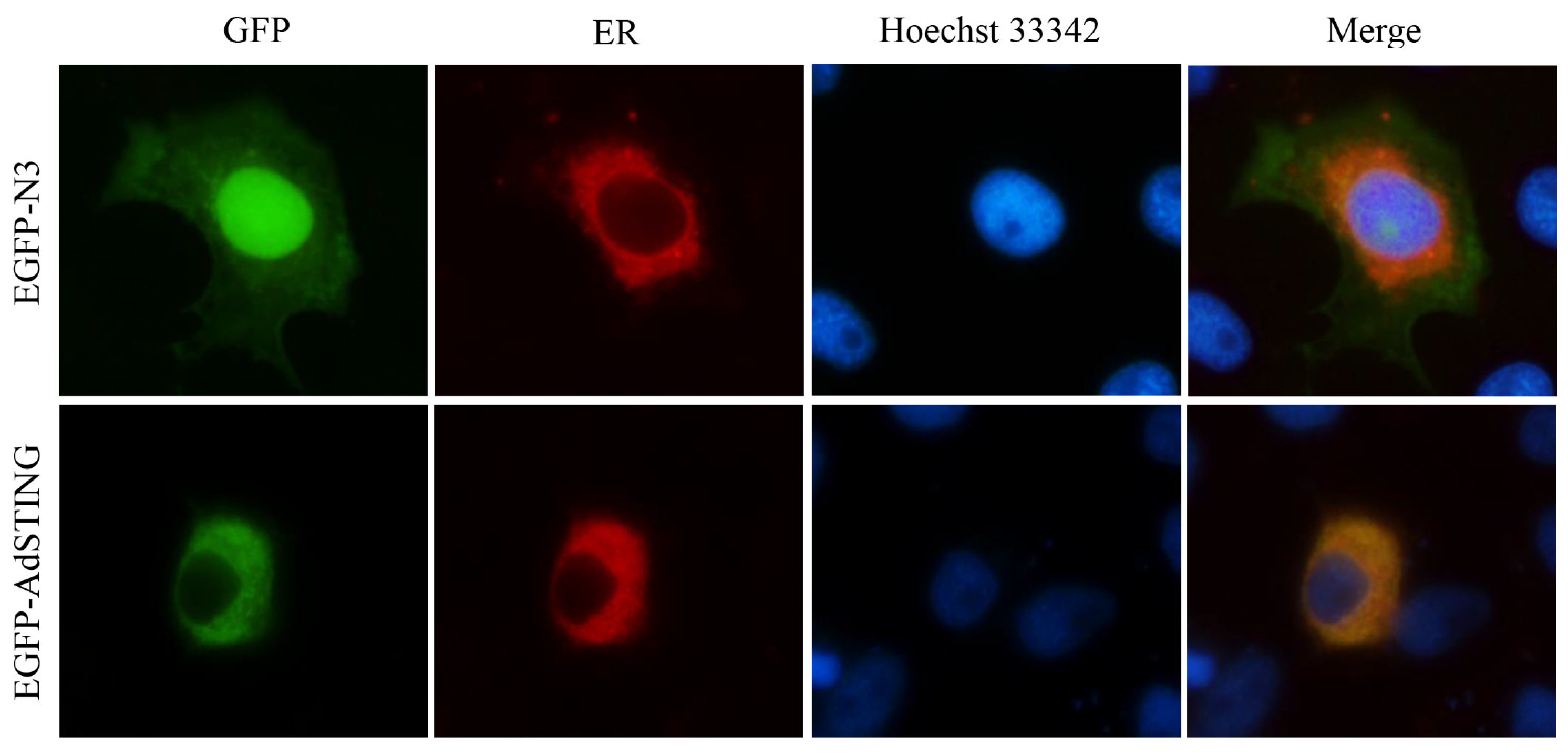
3.5. Subcellular Localization of AdSTING
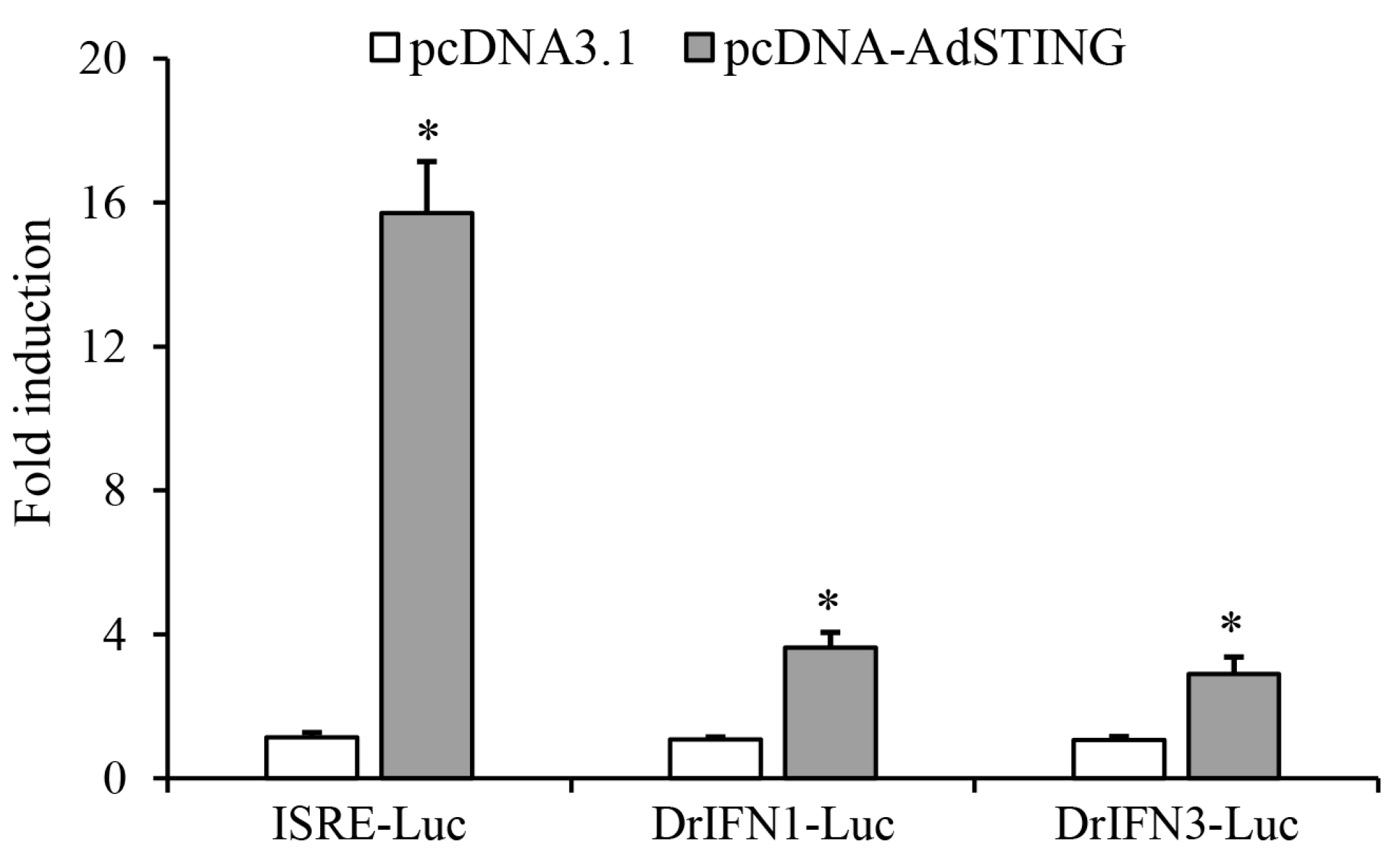
3.6. AdSTING Induced ISRE and IFN Promoter Activity
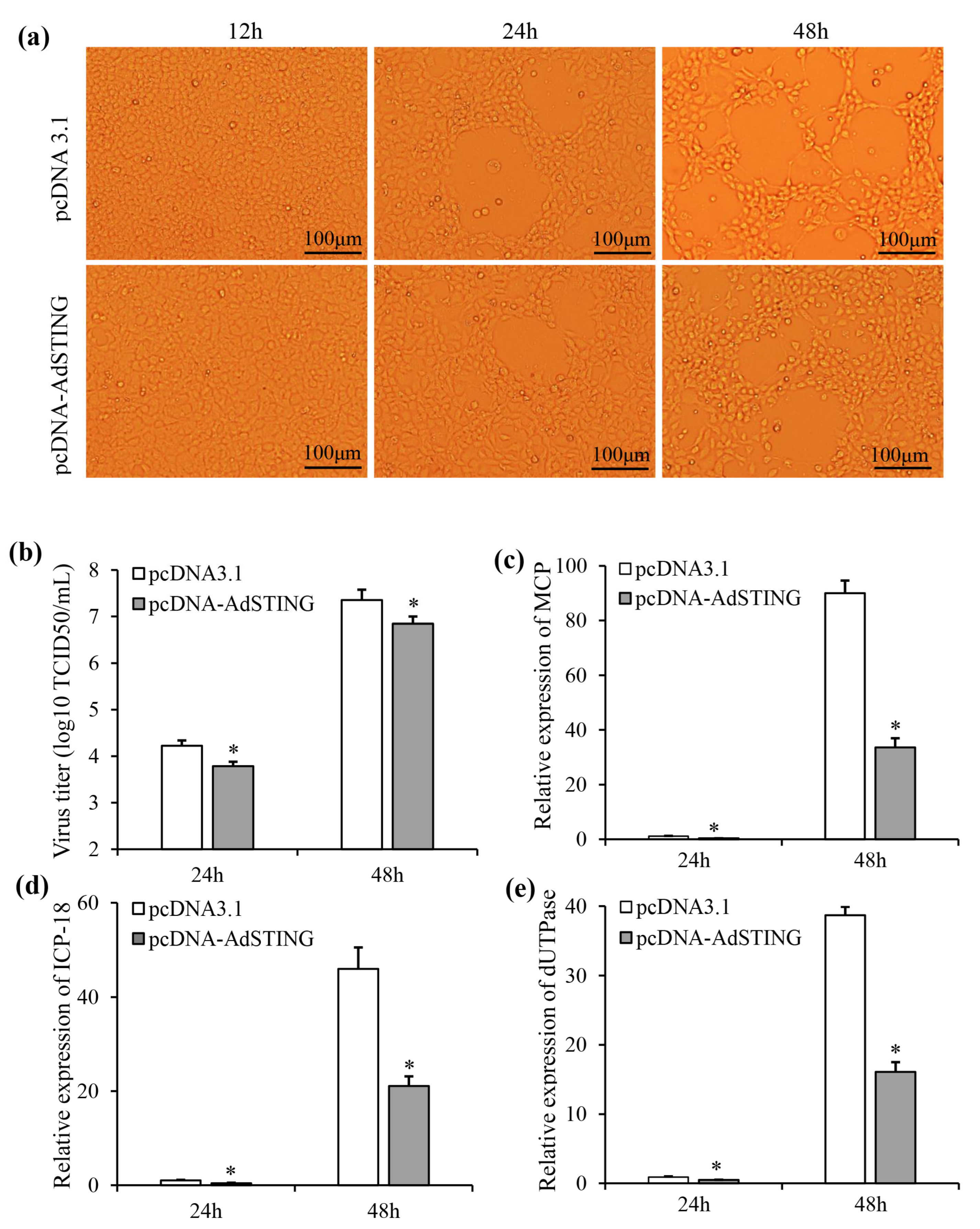
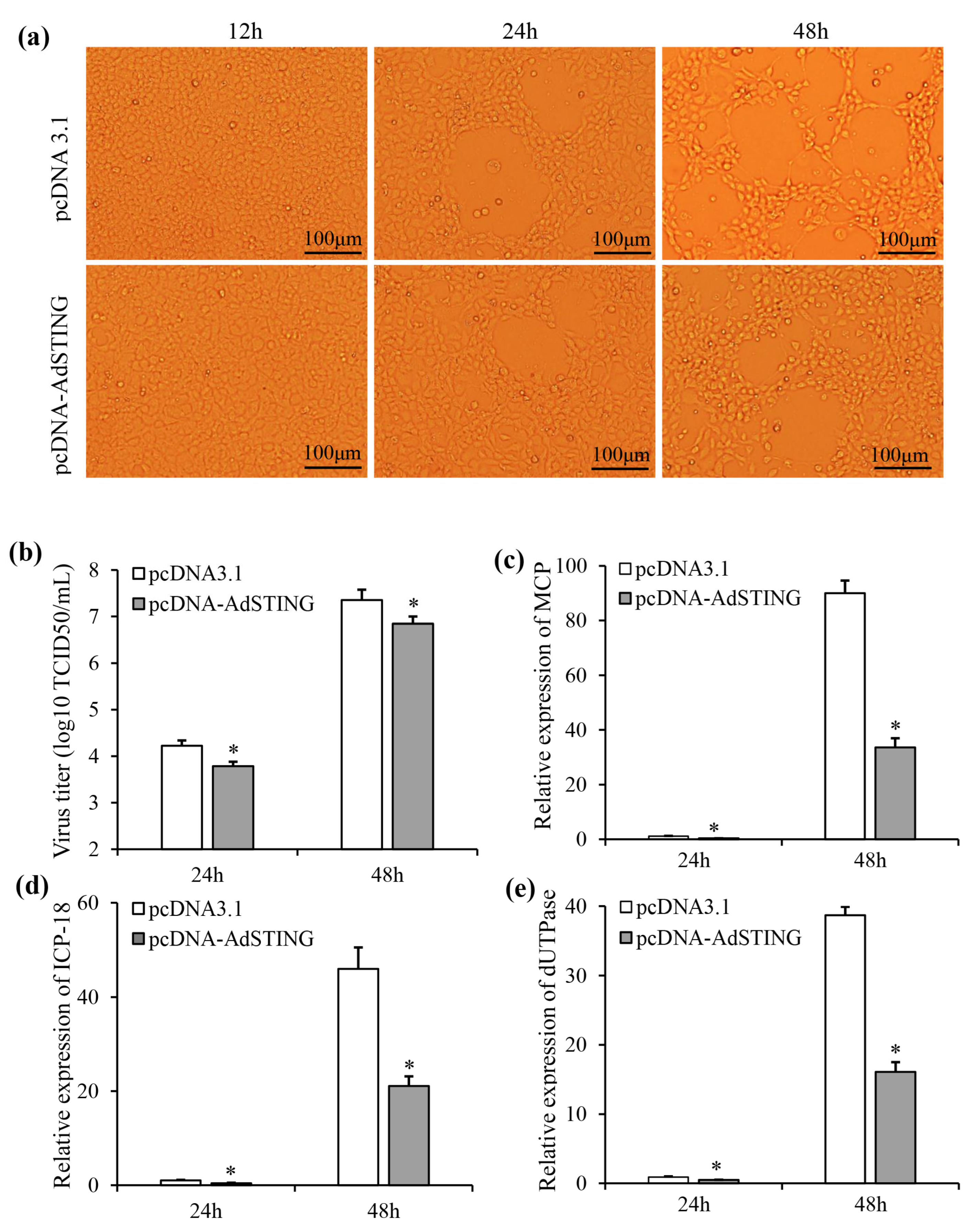
3.7. Overexpression of AdSTING-Reduced ADRV Replication
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chan, Y.K.; Gack, M.U. Viral evasion of intracellular DNA and RNA sensing. Nat. Rev. Microbiol. 2016, 14, 360–373. [Google Scholar] [CrossRef]
- Zhong, B.; Yang, Y.; Li, S.; Wang, Y.Y.; Li, Y.; Diao, F.; Lei, C.; He, X.; Zhang, L.; Tien, P.; et al. The adaptor protein MITA links virus-sensing receptors to IRF3 transcription factor activation. Immunity 2008, 29, 538–550. [Google Scholar] [CrossRef]
- Ishikawa, H.; Ma, Z.; Barber, G.N. STING regulates intracellular DNA-mediated, type I interferon-dependent innate immunity. Nature 2009, 461, 788–792. [Google Scholar] [CrossRef]
- Barber, G.N. STING-dependent cytosolic DNA sensing pathways. Trends Immunol. 2014, 35, 88–93. [Google Scholar] [CrossRef]
- Chauvin, S.; Stinson, W.A.; Platt, D.J.; Poddar, S.; Miner, J.J. Regulation of cGAS and STING signaling during inflammation and infection. J. Biol. Chem. 2023, 299, 104866. [Google Scholar] [CrossRef] [PubMed]
- Zevini, A.; Olagnier, D.; Hiscott, J. Crosstalk between cytoplasmic RIG-I and STING sensing pathways. Trends Immunol. 2017, 38, 194–205. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Hu, Z.Q.; Luo, Y.; Wang, H.Y.; Yu, X.; Wang, R.F. Function and regulation of cGAS-STING signaling in infectious diseases. Front. Immunol. 2023, 14, 1130423. [Google Scholar] [CrossRef] [PubMed]
- Abe, T.; Harashima, A.; Xia, T.; Konno, H.; Konno, K.; Morales, A.; Ahn, J.; Gutman, D.; Barber, G.N. STING recognition of cytoplasmic DNA instigates cellular defense. Mol. Cell 2013, 50, 5–15. [Google Scholar] [CrossRef]
- Hopfner, K.P.; Hornung, V. Molecular mechanisms and cellular functions of cGAS-STING signalling. Nat. Rev. Mol. Cell Biol. 2020, 21, 501–521. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.W.; Bai, X.C.; Chen, Z.J. Structures and mechanisms in the cGAS-STING innate immunity pathway. Immunity 2020, 53, 43–53. [Google Scholar] [CrossRef]
- Sun, F.; Zhang, Y.B.; Liu, T.K.; Shi, J.; Wang, B.; Gui, J.F. Fish MITA serves as a mediator for distinct fish IFN gene activation dependent on IRF3 or IRF7. J. Immunol. 2011, 187, 2531–2539. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.L.; Yang, C.R.; Zhang, Y.X.; Peng, L.M.; Chen, X.H.; Rao, Y.L.; Gu, T.L.; Su, J.G. Identification, characterization and immunological response analysis of stimulator of interferon gene (STING) from grass carp Ctenopharyngodon idella. Dev. Comp. Immunol. 2014, 45, 163–176. [Google Scholar] [CrossRef]
- Huang, Y.H.; Ouyang, Z.L.; Wang, W.; Yu, Y.P.; Li, P.F.; Zhou, S.; Wei, S.N.; Wei, J.G.; Huang, X.H.; Qin, Q.W. Antiviral role of grouper STING against iridovirus infection. Fish Shellfish Immunol. 2015, 47, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Wang, X.; Wu, S.Z.; Song, X.J.; Zou, Z.Q.; Xie, X.C.; Xiao, J.; Chen, S.; Feng, H. Black carp STING functions importantly in innate immune defense against RNA virus. Fish Shellfish Immunol. 2017, 70, 13–24. [Google Scholar] [CrossRef]
- Qin, X.W.; He, J.; Yu, Y.; Liu, C.; Luo, Z.Y.; Li, Z.M.; Weng, S.P.; Guo, C.J.; He, J.G. The roles of mandarin fish STING in innate immune defense against infectious spleen and kidney necrosis virus infections. Fish Shellfish Immunol. 2020, 100, 80–89. [Google Scholar] [CrossRef]
- Gao, F.Y.; Lu, M.X.; Wang, M.; Liu, Z.G.; Ke, X.L.; Zhang, D.F.; Cao, J.M. Nile tilapia DNA sensor STING is involved in the IFN-β and AP-1 signaling pathways in the immune response dependent on DDX41. Int. J. Biol. Macromol. 2023, 225, 27–39. [Google Scholar]
- Zhang, P.; Chen, Y.Q.; Liu, Y.F.; Zhou, H.; Qu, L.H. The complete mitochondrial genome of the Chinese giant salamander, Andrias davidianus (Amphibia: Caudata). Gene 2003, 311, 93–98. [Google Scholar] [CrossRef]
- Turvey, S.T.; Chen, S.; Tapley, B.; Wei, G.; Xie, F.; Yan, F.; Yang, J.; Liang, Z.; Tian, H.; Wu, M.; et al. Imminent extinction in the wild of the world’s largest amphibian. Curr. Biol. 2018, 28, R592–R594. [Google Scholar] [CrossRef]
- Chen, Z.Y.; Gui, J.F.; Gao, X.C.; Pei, C.; Hong, Y.J.; Zhang, Q.Y. Genome architecture changes and major gene variations of Andrias davidianus ranavirus (ADRV). Vet. Res. 2013, 44, 101. [Google Scholar] [CrossRef]
- Zhou, Z.Y.; Geng, Y.; Liu, X.X.; Ren, S.Y.; Zhou, Y.; Wang, K.Y.; Huang, X.L.; Chen, D.F.; Peng, X.; Lai, W.M. Characterization of a ranavirus isolated from the Chinese giant salamander (Andrias davidianus, Blanchard, 1871) in China. Aquaculture 2013, 384, 66–73. [Google Scholar] [CrossRef]
- Herath, J.; Ellepola, G.; Meegaskumbura, M. Patterns of infection, origins, and transmission of ranaviruses among the ectothermic vertebrates of Asia. Ecol. Evol. 2021, 11, 15498–15519. [Google Scholar] [CrossRef]
- Li, W.; Zhang, X.; Weng, S.P.; Zhao, G.X.; He, J.G.; Dong, C.F. Virion-associated viral proteins of a Chinese giant salamander (Andrias davidianus) iridovirus (genus Ranavirus) and functional study of the major capsid protein (MCP). Vet. Microbiol. 2014, 172, 129–319. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.D.; Chen, Z.Y.; Huang, X.; Gao, X.C.; Zhang, Q.Y. Establishment of three cell lines from Chinese giant salamander and their sensitivities to the wild-type and recombinant ranavirus. Vet. Res. 2015, 46, 58. [Google Scholar] [CrossRef] [PubMed]
- Jiang, N.; Fan, Y.D.; Zhou, Y.; Liu, W.Z.; Ma, J.; Meng, Y.; Xie, C.X.; Zeng, L.B. Characterization of Chinese giant salamander iridovirus tissue tropism and inflammatory response after infection. Dis. Aquat. Organ. 2015, 114, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Ke, F.; Gui, J.F.; Chen, Z.Y.; Li, T.; Lei, C.K.; Wang, Z.H.; Zhang, Q.Y. Divergent transcriptomic responses underlying the ranaviruses-amphibian interaction processes on interspecies infection of Chinese giant salamander. BMC Genomics 2018, 19, 211. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.Y.; Li, T.; Gao, X.C.; Wang, C.F.; Zhang, Q.Y. Protective immunity induced by DNA vaccination against ranavirus infection in Chinese giant salamander Andrias davidianus. Viruses 2018, 10, 52. [Google Scholar] [CrossRef] [PubMed]
- Ke, F.; Yu, X.D.; Wang, Z.H.; Gui, J.F.; Zhang, Q.Y. Replication and transcription machinery for ranaviruses: Components, correlation, and functional architecture. Cell Biosci. 2022, 12, 6. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Ma, J.; Fan, Y.D.; Meng, Y.; Xu, J.; Zhou, Y.; Liu, W.Z.; Zeng, X.H.; Zeng, L.B. Identification of type I IFN in Chinese giant salamander (Andrias davidianus) and the response to an iridovirus infection. Mol. Immunol. 2015, 65, 350–359. [Google Scholar] [CrossRef]
- Huang, L.; Fan, Y.D.; Zhou, Y.; Jiang, N.; Liu, W.Z.; Meng, Y.; Zeng, L.B. Cloning, sequence analysis and expression profiles of Toll-like receptor 7 from Chinese giantsalamander Andrias davidianus. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2015, 184, 52–57. [Google Scholar] [CrossRef]
- Yang, H.; Lan, Q.J.; Liu, R.R.; Cui, D.; Liu, H.X.; Xiong, D.M.; Li, F.G.; Liu, X.L.; Wang, L.X. Characterization of galectin-1 from Chinese giant salamanders Andrias davidianus and its involvements during immune response. Dev. Comp. Immunol. 2017, 70, 59–68. [Google Scholar] [CrossRef]
- Jiang, N.; Fan, Y.D.; Zhou, Y.; Meng, Y.; Liu, W.Z.; Li, Y.Q.; Xue, M.Y.; Robert, J.; Zeng, L.B. The immune system and the antiviral responses in Chinese giant salamander, Andrias davidianus. Front Immunol. 2021, 12, 718627. [Google Scholar] [CrossRef]
- Meng, Y.; Fan, Y.D.; Jiang, N.; Xue, M.Y.; Li, Y.Q.; Liu, W.Z.; Zeng, L.B.; Zhou, Y. Four Mx genes identified in Andrias davidianus and characterization of their response to Chinese giant salamander iridovirus infection. Animals 2022, 12, 2147. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Xia, H.; Yang, P.H.; Chen, Z.Y.; Zhang, Y.S.; Liu, L.G.; Luo, Y.S.; Meng, S.Y.; Fang, X.; Liu, Y.J.; Liu, W.; et al. Polymeric immunoglobulin receptor in dongtingking crucian carp (Carassius auratus indigentiaus): Molecular characterization and expression analysis in response to Aeromonas hydrophila challenge. Aquac. Res. 2022, 53, 3818–3827. [Google Scholar] [CrossRef]
- He, L.B.; Ke, F.; Wang, J.; Gao, X.C.; Zhang, Q.Y. Rana grylio virus (RGV) envelope protein 2L: Subcellular localization and essential roles in virus infectivity revealed by conditional lethal mutant. J. Gen. Virol. 2014, 95, 679–690. [Google Scholar] [CrossRef]
- Earl, J.E.; Chaney, J.C.; Sutton, W.B.; Lillard, C.E.; Kouba, A.J.; Langhorne, C.; Krebs, J.; Wilkes, R.P.; Hill, R.D.; Miller, D.L.; et al. Ranavirus could facilitate local extinction of rare amphibian species. Oecologia 2016, 182, 611–623. [Google Scholar] [CrossRef] [PubMed]
- Snyder, P.W.; Ramsay, C.T.; Harjoe, C.C.; Khazan, E.S.; Briggs, C.J.; Hoverman, J.T.; Johnson, P.T.J.; Preston, D.; Rohr, J.R.; Blaustein, A.R. Experimental evidence that host species composition alters host-pathogen dynamics in a ranavirus-amphibian assemblage. Ecology 2023, 104, e3885. [Google Scholar] [CrossRef]
- Gui, L.; Chinchar, V.G.; Zhang, Q.Y. Molecular basis of pathogenesis of emerging viruses infecting aquatic animals. Aquac. Fish. 2018, 3, 1–5. [Google Scholar] [CrossRef]
- Ouyang, S.Y.; Song, X.Q.; Wan, Y.Y.; Ru, H.; Shaw, N.; Jiang, Y.; Niu, F.F.; Zhu, Y.P.; Qiu, W.C.; Parvatiyar, K.; et al. Structural analysis of the STING adaptor protein reveals a hydrophobic dimer interface and mode of cyclic di-GMP binding. Immunity 2012, 36, 1073–1086. [Google Scholar] [CrossRef]
- Yan, J.; Qiao, G.X.; Chang, H.Y.; Wu, H.; Liu, M.L.; Tu, J.G.; Zhang, Y.G.; Feng, H. Role of the dimerization domain of black carp STING during the antiviral innate immunity. Reprod. Breed. 2023, 3, 59–65. [Google Scholar] [CrossRef]
- Tanaka, Y.; Chen, Z.J. STING specifies IRF3 phosphorylation by TBK1 in the cytosolic DNA signaling pathway. Sci. Signal. 2012, 5, ra20. [Google Scholar] [CrossRef]
- Zhao, B.Y.; Du, F.L.; Xu, P.B.; Shu, C.; Sankaran, B.; Bell, S.L.; Liu, M.M.; Lei, Y.J.; Gao, X.S.; Fu, X.F.; et al. A conserved PLPLRT/SD motif of STING mediates the recruitment and activation of TBK1. Nature 2019, 569, 718–722. [Google Scholar] [CrossRef]
- Amparyup, P.; Charoensapsri, W.; Soponpong, S.; Jearaphunt, M.; Wongpanya, R.; Tassanakajon, A. Stimulator of interferon gene (STING) and interferon regulatory factor (IRF) are crucial for shrimp antiviral defense against WSSV infection. Fish Shellfish Immunol. 2021, 117, 240–247. [Google Scholar] [CrossRef]
- Cheng, Y.Q.; Sun, Y.J.; Wang, H.G.; Yan, Y.X.; Ding, C.; Sun, J.H. Chicken STING mediates activation of the IFN gene independently of the RIG-I gene. J. Immunol. 2015, 195, 3922–3936. [Google Scholar] [CrossRef] [PubMed]
- Colombo, B.M.; Scalvenzi, T.; Benlamara, S.; Pollet, N. Microbiota and mucosal immunity in amphibians. Front. Immunol. 2015, 6, 111. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, V.L.; Robert, J. The amphibian immune system. Phil. Trans. R. Soc. B 2023, 378, 20220123. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Cai, X.; Wu, J.; Cong, Q.; Chen, X.; Li, T.; Du, F.; Ren, J.; Wu, Y.T.; Grishin, N.V.; et al. Phosphorylation of innate immune adaptor proteins MAVS, STING, and TRIF induces IRF3 activation. Science 2015, 347, aaa2630. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequence (5′-3′) | Usage |
|---|---|---|
| AdSTING-F1 | GAAATCAAGCACGTATAACTGCTGG | Gene cloning |
| AdSTING-R1 | CTTTCTTCTTTGTCATTCACCTGGC | |
| NUP | AAGCAGTGGTATCAACGCAGAGT | RACE-PCR |
| UPM | CTAATACGACTCACTATAGGGCAAGCAGTGGTATCAACGCAGAGT | |
| 5′GSP1 | CATTAACACATGCCTTTACAGCCTCAG | |
| 5′GSP2 | TTTCTTCTGTTAGGTCGCACACTCC | |
| 3′GSP1 | GTAGCGAAGATCGCATTCAACAAGC | |
| 3′GSP2 | CGTTCTCTATTTACATGGCAAATTCC | |
| AdSTING-32-F | GGGATCCATGAAAGAGTCTGTACGCCAAT | Plasmid construction |
| AdSTING-32-R | CCCTCGAGTTTCTCCATAGAATATTCC | |
| AdSTING-3.1-F | CGGGATCCAAGATGAATCGTGCATC | |
| AdSTING-3.1-R | GGAATTCTTATTTCTCCATAGAATATTCC | |
| AdSTING-N3-F | GGAATTCATGAATCGTGCATCAGA | |
| AdSTING-N3-R | CGGGATCCTTTCTCCATAGAATATTCC | |
| AdSTING-RT-F | TGCCATTGAGCATGACCATAAC | Real-time PCR |
| AdSTING-RT-R | TGTCTTCACATCTCAGGAGGTTC | |
| MCP-RT-F | CAGTCAGGGACATGGTCGTG | |
| MCP-RT-R | GGGAGTGACGCAGGTGTAAT | |
| ICP18-RT-F | CAGGTCAGGATCACCATGACTC | |
| ICP18-RT-F | TCCAGGTTGTCGTACAGGCAT | |
| DUT-RT-F | ACGGCAGGGTGGCTCCCAGG | |
| DUT-RT-R | CGGTAGAGTCCAGGCTGTCCA | |
| β-Actin-F | CCACTGCTGCCTCCTCTT | |
| β-Actin-R | GCAATGCCTGGGTACATG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Z.; Wang, R.; Wang, R.; Yu, N.; Yang, P. Identification and Characterization of the Stimulator of Interferon Genes (STING) in Chinese Giant Salamander Andrias davidianus. Fishes 2023, 8, 597. https://doi.org/10.3390/fishes8120597
Chen Z, Wang R, Wang R, Yu N, Yang P. Identification and Characterization of the Stimulator of Interferon Genes (STING) in Chinese Giant Salamander Andrias davidianus. Fishes. 2023; 8(12):597. https://doi.org/10.3390/fishes8120597
Chicago/Turabian StyleChen, Zhongyuan, Ronghua Wang, Ruisong Wang, Naitong Yu, and Pinhong Yang. 2023. "Identification and Characterization of the Stimulator of Interferon Genes (STING) in Chinese Giant Salamander Andrias davidianus" Fishes 8, no. 12: 597. https://doi.org/10.3390/fishes8120597
APA StyleChen, Z., Wang, R., Wang, R., Yu, N., & Yang, P. (2023). Identification and Characterization of the Stimulator of Interferon Genes (STING) in Chinese Giant Salamander Andrias davidianus. Fishes, 8(12), 597. https://doi.org/10.3390/fishes8120597