
Spatial Patterns in Fish Assemblages across the National Ecological Observation Network (NEON): The First Six Years

Abstract
1. Introduction
2. Materials and Methods
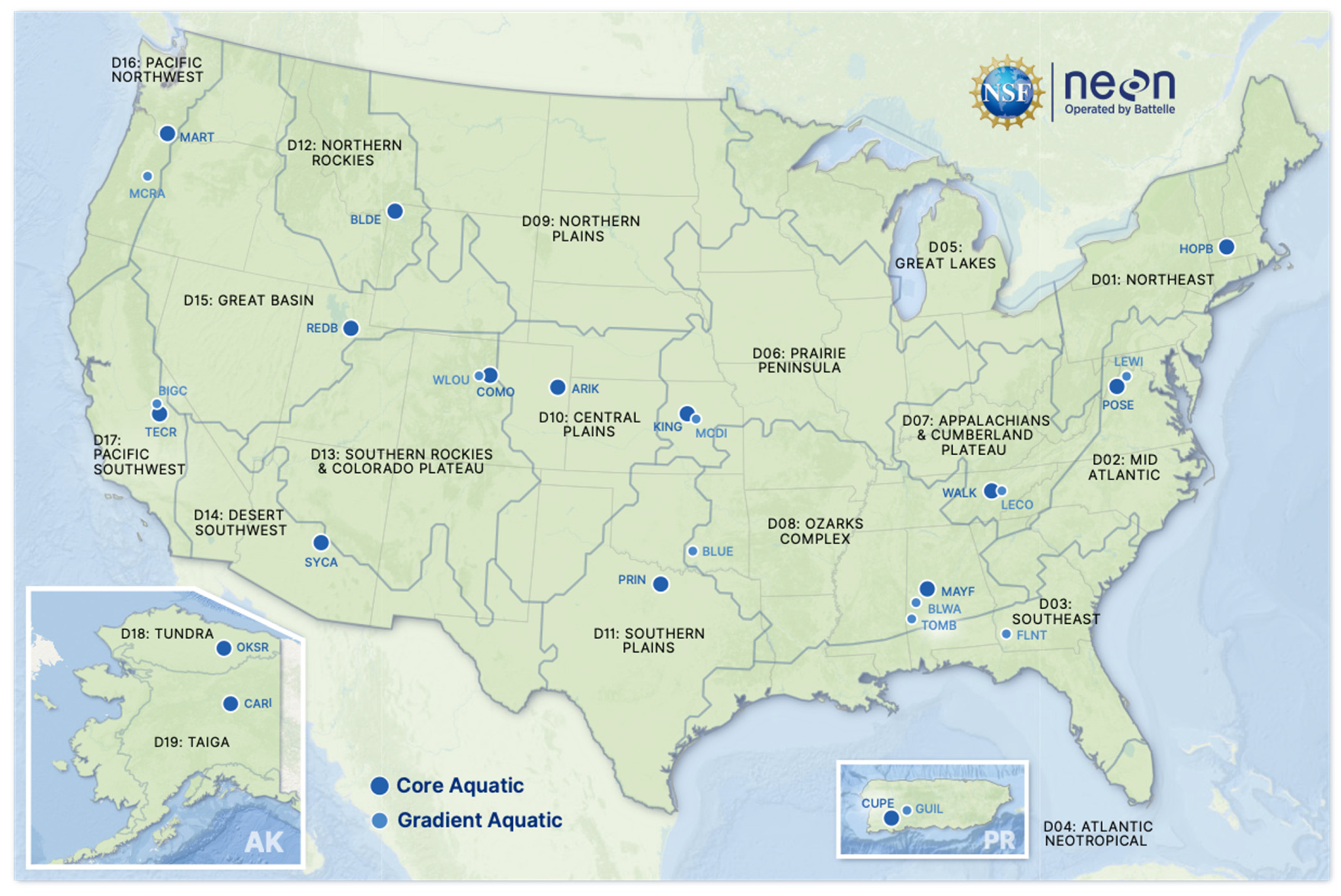
2.1. NEON Site Selection
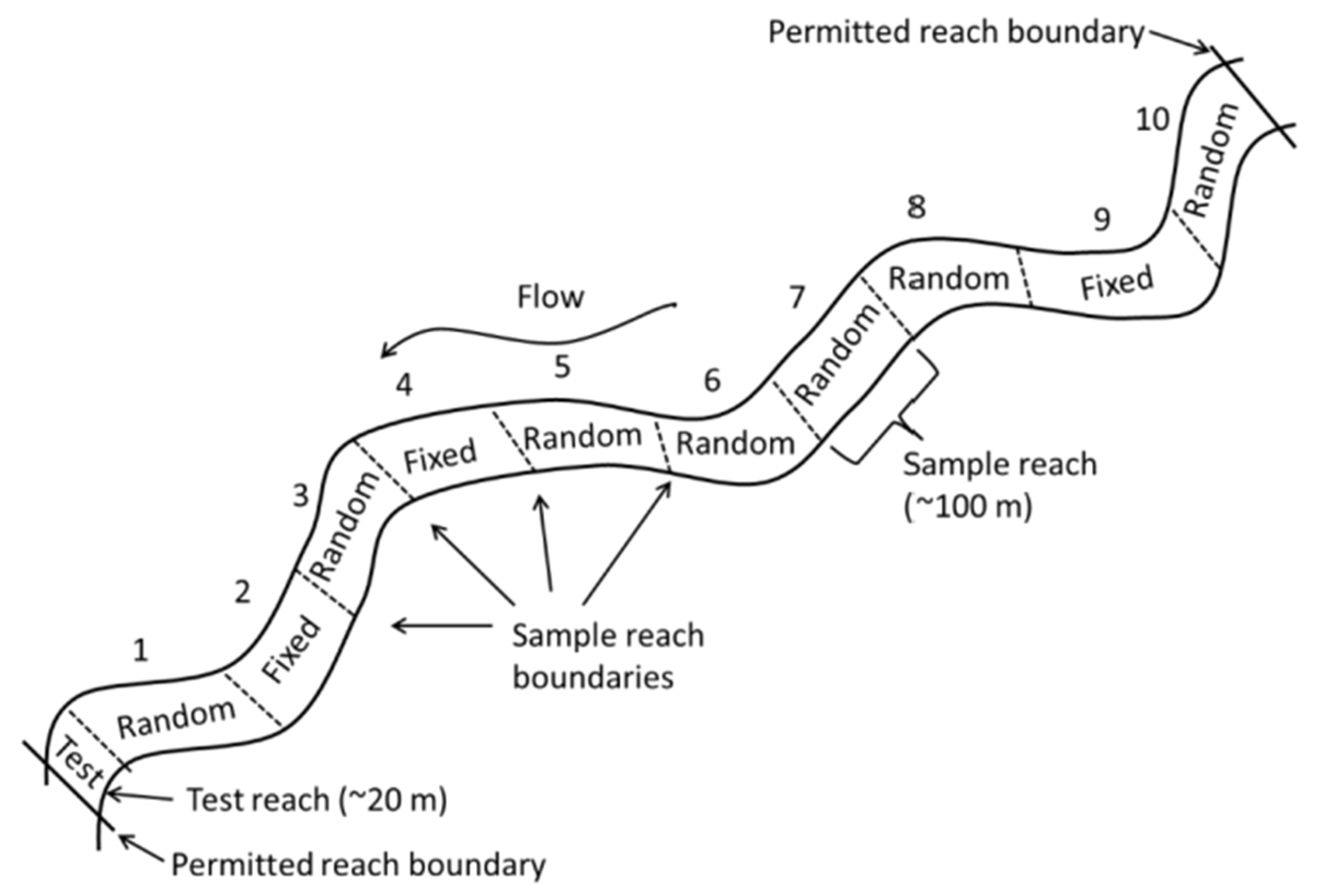
2.2. Biological Sampling Windows
2.3. NEON Fish Data
2.4. Downloading and Compiling NEON Fish Data
2.5. Alpha Diversity
2.6. Beta Diversity
2.7. Size Composition
3. Results
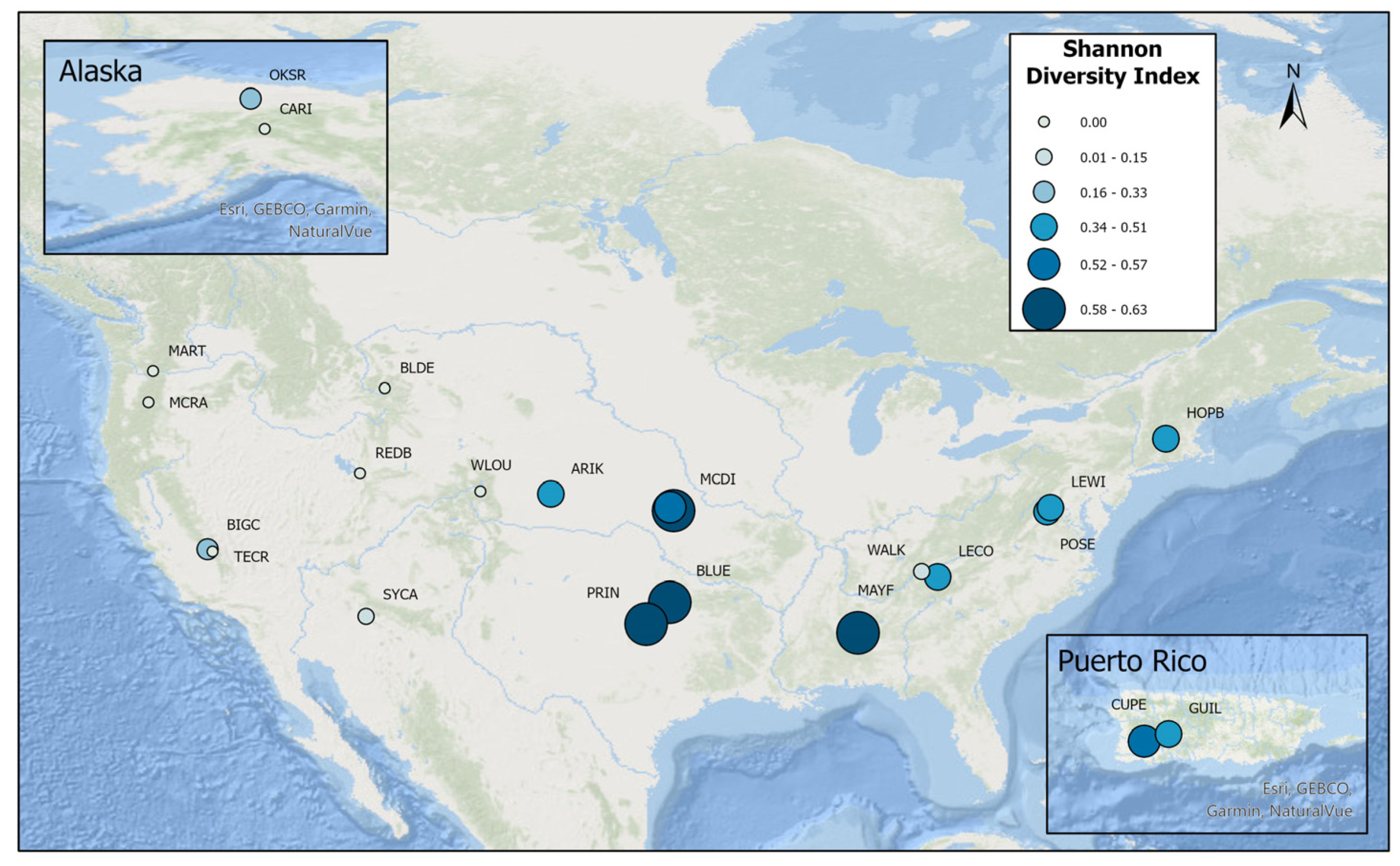
3.1. Alpha Diversity Metrics
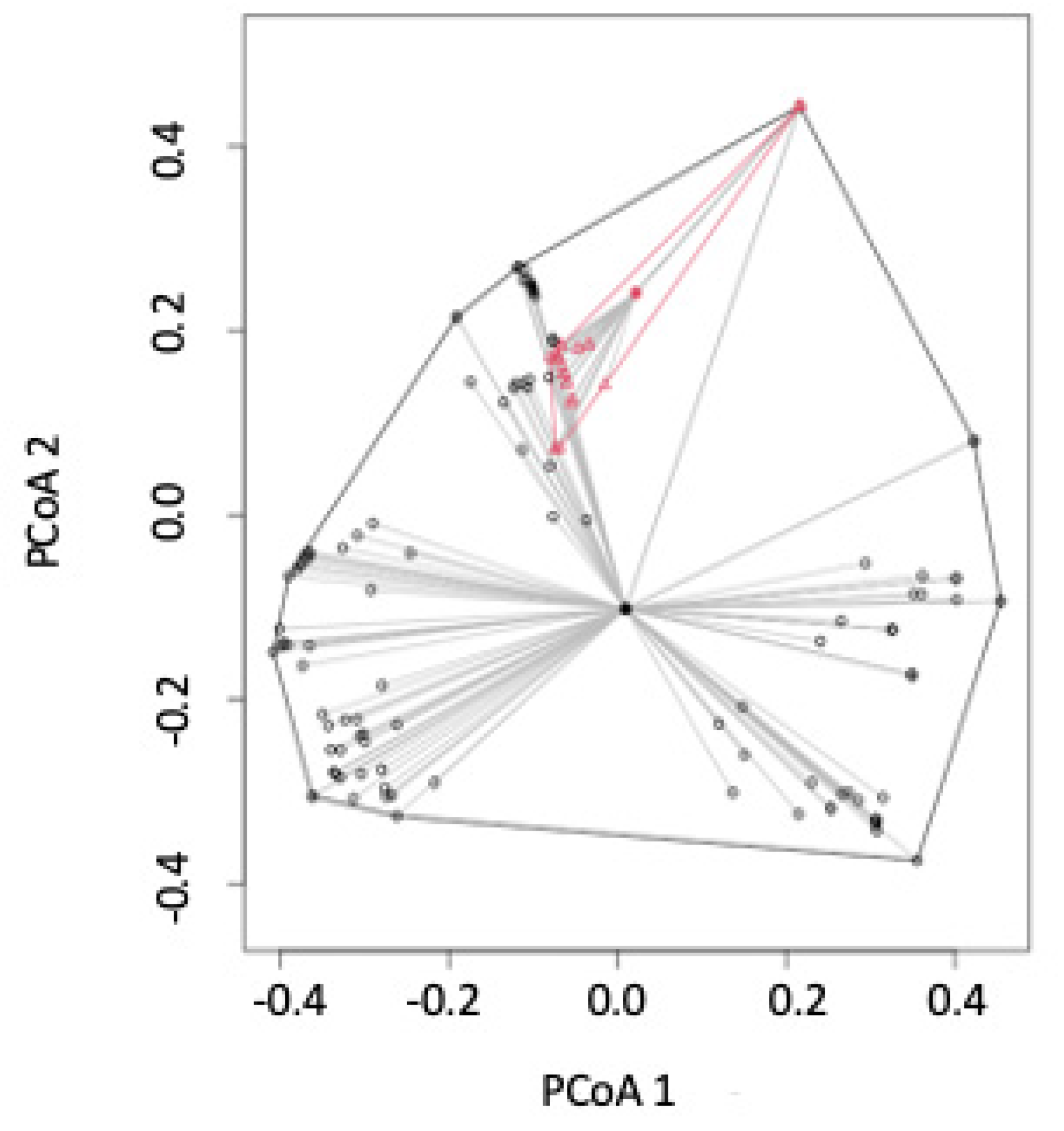
3.2. Beta Diversity Metrics
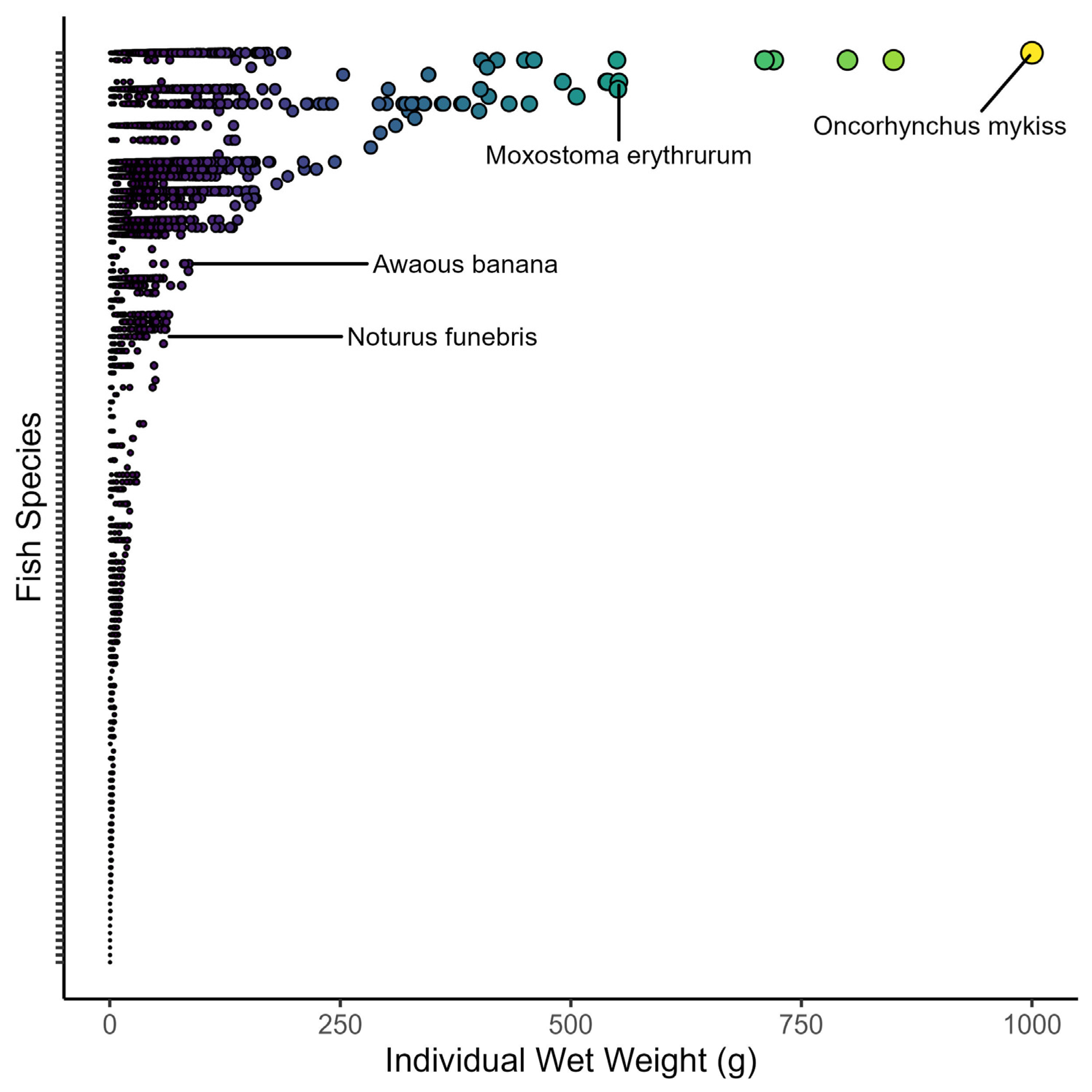
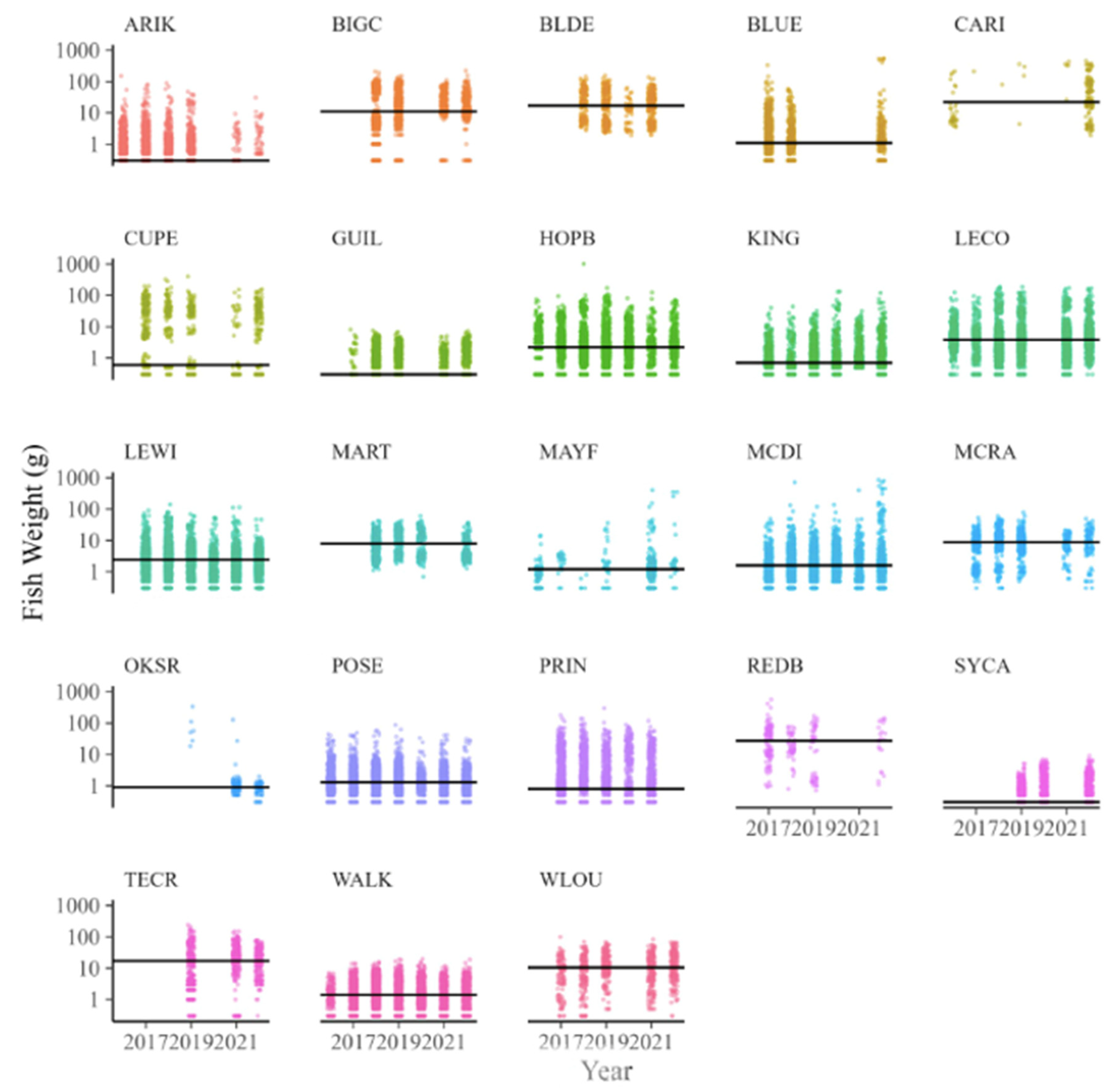
3.3. Size Composition
4. Discussion
4.1. Spatial Patterns of NEON Stream Fish Data
4.2. Occurence
4.3. Spatial Patterns in Size Composition
5. Summary and Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Mean | Spring Bout | Fall Bout | Highest | Lowest | Spring Bout Most Com. Spec. | Fall Bout Most Com Spec. | Domain |
|---|---|---|---|---|---|---|---|---|
| BLUE | 20.75 | 17.67 | 30 | 30 | 5 | ETHRAD (786) | ETHRAD (644) | Southern Plains |
| MAYF | 17 | 17 | 17 | 23 | 13 | NOTBAI (561) | NOTBAI (1053) | Ozarks Complex |
| PRIN | 9.5 | 10 | 9 | 11 | 7 | GAMAFF (654) | GAMAFF (1954) | Southern Plains |
| MCDI | 9.17 | 7.5 | 10.83 | 13 | 6 | CAMANO (1652) | CAMANO (566) | Prairie Peninsula |
| ARIK | 7.13 | 6.4 | 8.33 | 10 | 5 | ETHSPE (552) | * GAMAFF (1914) | Central Plains |
| KING | 6.42 | 5.33 | 7.5 | 9 | 3 | ETHSPE (790) | CHRERY (4911) | Prairie Peninsula |
| LEWI | 5.81 | 5.2 | 5.17 | 7 | 4 | COTGIR (1955) | COTGIR (2930) | Mid-Atlantic |
| CUPE | 4.4 | 4.8 | 4 | 7 | 2 | POERET (265) | POERET (111) | Atlantic NeoTropical |
| HOPB | 4.19 | 4.6 | 3 | 7 | 2 | RHIATR (674) | RHIATR (1382) | Northeast |
| POSE | 4.19 | 4.2 | 4.17 | 5 | 4 | RHIATR (1351) | RHIATR (2181) | Mid-Atlantic |
| LECO | 3.44 | 3.5 | 3.4 | 4 | 3 | RHIATR (755) | RHIATR (1965) | Appalachian and Cumberland Plateau |
| WALK | 2.46 | 2.4 | 2.5 | 4 | 2 | RHIATR (1221) | RHIATR (2811) | Appalachian and Cumberland Plateau |
| GUIL | 2.3 | 2.4 | 2.2 | 3 | 2 | POERET (3698) | POERET (4199) | Atlantic NeoTropical |
| BIGC | 2.29 | 2.75 | 1.667 | 3 | 1 | * SALTRU (645) | * SALTRU (695) | Pacific Southwest |
| SYCA | 2.25 | 2.33 | 2 | 3 | 1 | AGOCHR (2410) | AGOCHR (699) | Desert Southwest |
| OKSR | 1.33 | 2 | 1 | 2 | 1 | THYARC (1) | THYARC (64) | Tundra |
| CARI | 1.14 | 0.75 | 1.67 | 3 | 1 | THYARC (8) | THYARC (87) | Taiga |
| BLDE | 1 | NA | 1 | 1 | 1 | NA | * SALFON (587) | Northern Rockies |
| MART | 1 | NA | 1 | 1 | 1 | NA | SALSP (785) | Pacific Northwest |
| MCRA | 1 | NA | 1 | 1 | 1 | NA | ONCCLA (720) | Pacific Northwest |
| REDB | 1 | 1 | 1 | 1 | 1 | ONCCLA (24) | ONCCLA (154) | Great Basin |
| TECR | 1 | 1 | 1 | 1 | 1 | * SALFON (529) | * SALFON (387) | Pacific Southwest |
| WLOU | 1 | 1 | 1 | 1 | 1 | * SALFON (74) | * SALFON (494) | Southern Rockies and Colorado Plateau |
| Site | Spring/Fall | Species | Count |
|---|---|---|---|
| ARIK | Spring | Etheostoma exile | 206 |
| BLUE | Fall | Ameiurus melas | 1 |
| BLUE | Spring | Cyprinella camura | 1 |
| BLUE | Fall | Lythrurus umbratilis | 6 |
| BLUE | Spring | Micropterus punctulatus | 1 |
| BLUE | Spring | Micropterus salmoides | 1 |
| BLUE | Spring | Notropis boops | 13 |
| BLUE | Spring | Notropis buchanani | 6 |
| BLUE | Fall | Pylodictis olivaris | 1 |
| CUPE | Spring | Anguilla rostrata | 3 |
| GUIL | Fall | Tilapia rendalli | 1 |
| HOPB | Spring | Ameiurus nebulosus | 1 |
| HOPB | Spring | Notemigonus crysoleucas | 1 |
| HOPB | Spring | Noturus gyrinus | 1 |
| HOPB | Fall | Notemigonus crysoleucas | 6 |
| KING | Spring | Cyprinella lutrensis | 2 |
| KING | Spring | Etheostoma pseudovulatum | 4 |
| KING | Spring | Etheostoma tennesseense | 1 |
| KING | Fall | Lepomis macrochirus | 1 |
| KING | Fall | Luxilus cornutus | 1 |
| KING | Fall | Moxostoma pisolabrum | 1 |
| KING | Fall | Phoxinus erythrogaster | 78 |
| LECO | Spring | Campostoma anomalum | 1 |
| LEWI | Fall | Cyprinella spiloptera | 1 |
| LEWI | Spring | Etheostoma flabellare | 1 |
| LEWI | Spring | Lepomis cyanellus | 1 |
| LEWI | Spring | Lepomis macrochirus | 4 |
| MAYF | Fall | Elassoma zonatum | 1 |
| MAYF | Fall | Erimyzon oblongus | 1 |
| MAYF | Spring | Lepomis auritus | 2 |
| MAYF | Spring | Lepomis cyanellus | 1 |
| MAYF | Fall | Etheostoma histrio | 1 |
| MAYF | Spring | Lepomis macrochirus | 3 |
| MAYF | Fall | Lythrurus bellus | 2 |
| MAYF | Spring | Minytrema melanops | 2 |
| MAYF | Fall | Micropterus henshalli | 4 |
| MAYF | Fall | Micropterus warriorensis | 1 |
| MAYF | Spring | Moxostoma poecilurum | 10 |
| MAYF | Fall | Notropis stilbius | 65 |
| MAYF | Spring | Pteronotropis hypselopterus | 1 |
| MCDI | Fall | Catostomus commersonii | 1 |
| MCDI | Fall | Etheostoma nigrum | 133 |
| MCDI | Fall | Lepomis megalotis | 3 |
| MCDI | Fall | Pimephales vigilax | 5 |
| PRIN | Fall | Cyprinus carpio | 1 |
| PRIN | Spring | Micropterus salmoides | 2 |
| PRIN | Spring | Notropis volucellus | 71 |
| PRIN | Spring | Pimephales vigilax | 1 |
| WALK | Fall | Notropis atherinoides | 1 |
| Site | Species | Years Caught | Count |
|---|---|---|---|
| ARIK | Ameiurus melas | 2017–2019 | 16 |
| ARIK | Etheostoma exile | 2017–2019 | 206 |
| ARIK | Fundulus zebrinus | 2017–2019 | 20 |
| ARIK | Lepomis cyanellus | 2017–2019 | 203 |
| BLUE | Ameiurus melas | 2017–2019 | 1 |
| BLUE | Lythrurus umbratilis | 2017–2019 | 6 |
| BLUE | Cyprinella camura | 2020–2022 | 1 |
| BLUE | Micropterus salmoides | 2017–2019 | 1 |
| BLUE | Micropterus punctulatus | 2017–2019 | 1 |
| BLUE | Nocomis asper | 2017–2019 | 10 |
| BLUE | Notropis buchanani | 2020–2022 | 6 |
| BLUE | Notropis nubilus | 2020–2022 | 1 |
| BLUE | Notropis suttkusi | 2017–2019 | 61 |
| BLUE | Notropis volucellus | 2017–2019 | 99 |
| BLUE | Pimephales notatus | 2017–2019 | 79 |
| BLUE | Pylodictis olivaris | 2017–2019 | 1 |
| CUPE | Anguilla rostrata | 2017–2019 | 3 |
| CUPE | Gobiomorus dormitor | 2020–2022 | 4 |
| CUPE | Sicydium punctatum | 2017–2019 | 30 |
| CUPE | Sicydium plumieri | 2017–2019 | 45 |
| GUIL | Gambusia affinis | 2017–2019 | 43 |
| GUIL | Tilapia rendalli | 2020–2022 | 1 |
| HOPB | Ameiurus nebulosus | 2017–2019 | 1 |
| HOPB | Noturus gyrinus | 2017–2019 | 1 |
| HOPB | Salmo trutta | 2017–2019 | 57 |
| KING | Cyprinella lutrensis | 2020–2022 | 2 |
| KING | Etheostoma pseudovulatum | 2017–2019 | 4 |
| KING | Etheostoma tennesseense | 2017–2019 | 1 |
| KING | Luxilus cornutus | 2020–2022 | 1 |
| KING | Lepomis macrochirus | 2017–2019 | 1 |
| KING | Moxostoma pisolabrum | 2020–2022 | 1 |
| KING | Notropis percobromus | 2020–2022 | 1 |
| KING | Phoxinus erythrogaster | 2017–2019 | 78 |
| KING | Noturus exilis | 2020–2022 | 2 |
| LEC0 | Campostoma anomalum | 2017–2019 | 1 |
| LEWI | Gambusia holbrooki | 2020–2022 | 46 |
| LEWI | Lepomis cyanellus | 2020–2022 | 1 |
| LEWI | Nocomis leptocephalus | 2020–2022 | 3 |
| LEWI | Etheostoma flabellare | 2017–2019 | 1 |
| LEWI | Lepomis macrochirus | 2017–2019 | 4 |
| MAYF | Elassoma zonatum | 2017–2019 | 1 |
| MAYF | Lythrurus bellus | 2017–2019 | 2 |
| MAYF | Minytrema melanops | 2017–2019 | 2 |
| MAYF | Notropis ammophilus | 2017–2019 | 4 |
| MAYF | Erimyzon oblongus | 2020–2022 | 1 |
| MAYF | Etheostoma chlorosomum | 2020–2022 | 2 |
| MAYF | Etheostoma histrio | 2020–2022 | 1 |
| MAYF | Etheostoma nigrum | 2020–2022 | 7 |
| MAYF | Lepomis cyanellus | 2020–2022 | 1 |
| MAYF | Lepomis macrochirus | 2020–2022 | 3 |
| MAYF | Micropterus warriorensis | 2020–2022 | 1 |
| MAYF | Notropis volucellus | 2020–2022 | 30 |
| MAYF | Pteronotropis hypselopterus | 2020–2022 | 1 |
| MCDI | Ameiurus natalis | 2020–2022 | 4 |
| MCDI | Catostomus commersonii | 2020–2022 | 1 |
| MCDI | Notropis atherinoides | 2017–2019 | 15 |
| MCDI | Notropis shumardi | 2017–2019 | 4 |
| MCDI | Micropterus punctulatus | 2017–2019 | 4 |
| MCDI | Etheostoma nigrum | 2017–2019 | 133 |
| POSE | Cottus bairdii | 2017–2019 | 1010 |
| PRIN | Cyprinus carpio | 2017–2019 | 1 |
| PRIN | Micropterus salmoides | 2017–2019 | 2 |
| PRIN | Notropis stramineus | 2017–2019 | 1 |
| PRIN | Notropis volucellus | 2017–2019 | 71 |
| PRIN | Pimephales vigilax | 2017–2019 | 1 |
| WALK | Cottus caeruleomentum | 2017–2019 | 55 |
| WALK | Notropis atherinoides | 2017–2019 | 1 |
References
- Zogaris, S.; Koutsikos, N.; Chatzinikolaou, Y.; Őzeren, S.C.; Yence, K.; Vlami, V.; Kohlmeier, P.G.; Akyildiz, G.K. Fish assemblages as ecological indicators in the Büyük Menderes (Great Meander) River, Turkey. Water 2023, 15, 2292. [Google Scholar] [CrossRef]
- Pont, D.; Hughes, R.M.; Whittier, T.R.; Schmutz, S. A predictive index of biotic integrity model for aquatic-vertebrate assemblages of western U.S. streams. Trans. Am. Fish. Soc. 2009, 138, 292–305. [Google Scholar] [CrossRef]
- Pont, D.; Hugueny, B.; Beier, U.; Goffaux, D.; Melcher, A.; Noble, R.; Rogers, C.; Roset, N.; Schmutz, S. Assessing river biotic condition at a continental scale: European approach using functional metrics and fish assemblages. J. Appl. Ecol. 2006, 43, 70–80. [Google Scholar] [CrossRef]
- Vadas, R.L., Jr.; Hughes, R.M.; Bello-Gonzales, O.; Callisto, M.; Carvalho, D.; Chen, K.; Davies, P.E.; Ferreira, M.T.; Fierro, P.; Harding, J.S.; et al. Assemblage-based biomonitoring of freshwater ecosystem health via multimetric indices: A critical review and suggestions for improving their applicability. Water Biol. Secur. 2022, 1, 100054. [Google Scholar] [CrossRef]
- Cawley, K.M.; Parker, S.; Utz, R.; Goodman, K.; Scott, C.; Fitzgerald, M.; Vance, J.; Jensen, B.; Bohall, C.; Baldwin, T. NEON aquatic sampling strategy 2022, NEON.DOC.001152vB. NEON (National Ecological Observatory Network). Available online: https://data.neonscience.org/api/v0/documents/NEON.DOC.001152vA (accessed on 1 May 2023).
- McDowell, W.H. NEON and STREON: Opportunities and challenges for the aquatic sciences. Freshw. Sci. 2015, 34, 386–391. [Google Scholar] [CrossRef]
- Tesfaye, C.T.; Souza, A.T.; Barton, D.; Blabolil, P.; Cech, M.; Drastik, V.; Jaroslava, F.; Holubova, M.; Kocvara, L.; Kolarik, T.; et al. Long-term monitoring of fish in a freshwater reservoir: Different ways of weighting complex spatial samples. Front. Environ. Sci. 2022, 6, 303. [Google Scholar] [CrossRef]
- da Silva, V.E.L.; Silva-Firmiano, L.P.S.; Teresa, F.B.; Batista, V.S.; Ladle, R.J.; Fabré, N.N. Functional traits of fish species: Adjusting resolution to accurately express resource partitioning. Front. Mar. Sci. 2019, 6, 303. [Google Scholar] [CrossRef]
- Dias, M.S.; Zuanon, J.; Couto, T.B.; Carvalho, M.; Carvalho, L.N.; Espírito-Santo, H.M.; Frederico, R.; Leitão, R.P.; Mortati, A.F.; Pires, T.H.; et al. Trends in studies of Brazilian stream fish assemblages. Nat. Conserv. 2016, 14, 106–111. [Google Scholar] [CrossRef]
- Tonn, W.M. Climate change and fish communities: A conceptual framework. Trans. Am. Fish. Soc. 1990, 119, 337–352. [Google Scholar] [CrossRef]
- Azaele, S.; Muneepeerakul, R.; Maritan, A.; Rinaldo, A.; Rodriguez-Iturbe, I. Predicting spatial similarity of freshwater fish biodiversity. Proc. Natl. Acad. Sci. USA 2009, 106, 7058–7062. [Google Scholar] [CrossRef]
- Smith, G.R.; Badgley, C.; Eiting, T.P.; Larson, P.S. Species diversity gradients in relation to geological history in North American freshwater fishes. Evolut. Ecol. Res. 2010, 2, 693–726. [Google Scholar]
- Hocutt, C.H.; Wiley, E.O. The Zoogeography of North American Freshwater Fishes; Wiley: New York, NY, USA, 1986. [Google Scholar]
- Parker, S.M.; Utz, R.M. Temporal design for aquatic organismal sampling across the National Ecological Observatory Network. Methods Ecol. Evol. 2022, 13, 1834–1848. [Google Scholar] [CrossRef]
- A NEON (National Ecological Observatory Network). Aquatic Field Sites—Excluding Lake Sites—Across the NEON Domains. Courtesy of National Ecological Observatory Network/Battelle. Available online: https://www.neonscience.org/field-sites/explore-field-sites (accessed on 1 November 2023).
- Monahan, D.; Jensen, B.; Parker, S.; Fischer, J.R. AOS Protocol and Procedure: FSS-Fish Sampling in Wadeable Streams. NEON.DOC.001295vG.NEON2021 (National Ecological Observatory Network). Available online: https://data.neonscience.org/documents/10179/1883159/NEON.DOC.001295vG (accessed on 1 November 2023).
- Froese, R.; Pauly, D. FishBase. World Wide Web Electronic Publication. Available online: https://www.fishbase.de/home.htm (accessed on 1 October 2016).
- NEON (National Ecological Observatory Network). Fish Electrofishing, Gill Netting, and Fyke Netting Counts (DP1.20107.001), RELEASE-2023. Available online: https://data.neonscience.org/data-products/DP1.20107.001/RELEASE-2023 (accessed on 16 April 2023). [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; O’hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.; Wagner, H. Vegan: Community Ecology Package. R Package Version 2.4-6. 2022. Available online: https://CRAN.R-project.org/package=vegan (accessed on 1 May 2023).
- Hughes, R.M.; Herlihy, A.T.; Comeleo, R.; Peck, D.V.; Mitchell, R.M.; Paulsen, S.G. Patterns in and predictors of stream and river macroinvertebrate genera and fish species richness across the conterminous USA. Knowl. Manag. Aquat. Ecosyst. 2023, 424, 1–16. [Google Scholar] [CrossRef]
- Kanno, Y.; Vokoun, J.C.; Dauwalter, D.C.; Hughes, R.M.; Herlihy, A.T.; Maret, T.R.; Patton, T.M. Influence of rare species on electrofishing distance–species richness relationships at stream sites. Trans. Am. Fish. Soc. 2009, 138, 1240–1251. [Google Scholar] [CrossRef]
- Hughes, R.M.; Herlihy, A.T.; Peck, D.V. Sampling effort for estimating fish species richness in western USA river sites. Limnologica 2021, 87, 125859. [Google Scholar] [CrossRef] [PubMed]
- Schlosser, I.J.; Angermeier, P.L. Spatial variation in demographic processes in lotic fishes: Conceptual models, empirical evidence, and implications for conservation. Am. Fish. Soc. Symp. 1995, 17, 360–370. [Google Scholar]
- Fausch, K.D.; Torgersen, C.E.; Baxter, C.V.; Li, H.W. Landscapes to riverscapes: Bridging the gap between research and conservation of stream fishes. BioScience 2002, 52, 483–498. [Google Scholar] [CrossRef]
- Vadas, R.L., Jr. Seasonal habitat use, species associations, and assemblage structure of forage fishes in Goose Creek, northern Virginia. I. Macrohabitat patterns. J. Freshw. Ecol. 1991, 6, 403–417. [Google Scholar] [CrossRef]
- White, E.P.; Ernest, S.M.; Kerkhoff, A.J.; Enquist, B.J. Relationships between body size and abundance in ecology. Trends Ecol. Evol. 2007, 22, 323–330. [Google Scholar] [CrossRef]
- Brown, J.H.; Gillooly, J.F.; Allen, A.P.; Savage, V.M.; West, G.B. Toward a Metabolic Theory of Ecology. Ecology 2004, 85, 1771–1789. [Google Scholar] [CrossRef]
- Jennings, S. Size-based analyses of aquatic food webs. In Aquatic Food Webs: An Ecosystem Approach; Oxford University Press: Oxford, UK, 2005; pp. 86–97. [Google Scholar]
- Sprules, W.G.; Barth, L.E. Surfing the biomass size spectrum: Some remarks on history, theory, and application. Can. J. Fish. Aquat. Sci. 2016, 73, 477–495. [Google Scholar] [CrossRef]
- Arranz, I.; Grenouillet, G.; Cucherousset, J. Biological invasions and eutrophication reshape the spatial patterns of stream fish size spectra in France. Divers. Distrib. 2023, 29, 590–597. [Google Scholar] [CrossRef]
- Pomeranz, J.P.F.; Junker, J.R.; Wesner, J.S. Individual size distributions across North American streams vary with local temperature. Glob. Chang. Biol. 2021, 28, 848–858. [Google Scholar] [CrossRef] [PubMed]
- Pompeu, P.S.; Wouters, L.; Hilário, H.O.; Loures, R.C.; Peressin, A.; Prado, I.G.; Suzuki, F.M.; Carvalho, D.C. Inadequate Sampling Frequency and Imprecise Taxonomic Identification Mask Results in Studies of Migratory Freshwater Fish Ichthyoplankton. Fishes 2023, 8, 518. [Google Scholar] [CrossRef]






| NEON Site Name | Drainage | Domain Number | Domain Name |
|---|---|---|---|
| HOPB | Atlantic | 01 | Northeast |
| LEWI | Atlantic | 02 | Mid-Atlantic |
| POSE | Atlantic | 02 | Mid-Atlantic |
| CUPE | Atlantic | 04 | Atlantic Neotropical |
| GUIL | Atlantic | 04 | Atlantic Neotropical |
| MCDI | Atlantic | 06 | Prairie Peninsula |
| KING | Atlantic | 06 | Prairie Peninsula |
| LECO | Atlantic | 07 | Appalachian |
| WALK | Atlantic | 07 | Appalachian |
| MAYF | Atlantic | 08 | Ozark Complex |
| MAYF | Atlantic | 08 | Ozark Complex |
| ARIK | Atlantic | 10 | Central Plains |
| BLUE | Atlantic | 11 | Southern Plains |
| PRIN | Atlantic | 11 | Southern Plains |
| BLDE | Atlantic | 12 | Northern Rockies |
| WLOU | Pacific | 13 | Southern Rockies |
| SYCA | Pacific | 14 | Desert Southwest |
| REDB | Pacific | 15 | Great Basin |
| MART | Pacific | 16 | Pacific Northwest |
| MCRA | Pacific | 16 | Pacific Northwest |
| BIGC | Pacific | 17 | Pacific Southwest |
| TECR | Pacific | 17 | Pacific Southwest |
| OKSR | Pacific | 18 | Tundra |
| CARI | Pacific | 19 | Taiga |
| Domain | Site | Spring Sampling Window | Fall Sampling Window |
|---|---|---|---|
| 1 | HOPB | 11 Apr–9 May | 3 Oct–31 Oct |
| 2 | POSE | 19 Mar–16 Apr | 18 Oct–15 Nov |
| 2 | LEWI | 19 Mar–16 Apr | 18 Oct–15 Nov |
| 4 | CUPE | 24 Jan–21 Feb | 10 Nov–8 Dec |
| 4 | GUIL | 26 Jan–23 Feb | 9 Nov–7 Dec |
| 6 | KING | 23 Mar–20 Apr | 3 Oct–31 Oct |
| 6 | MCDI | 20 Mar–17 Apr | 27 Sep–25 Oct |
| 7 | LECO | 15 Mar–12 Apr | 12 Oct–9 Nov |
| 7 | WALK | 09 Mar–06 Apr | 19 Oct–16 Nov |
| 8 | MAYF | 05 Mar–02 Apr | 24 Oct–28 Nov |
| 10 | ARIK | 21 Mar–18 Apr | 20 Sep–18 Oct |
| 11 | PRIN | 17 Feb–17 Mar | 23 Oct–20 Nov |
| 11 | BLUE | 07 Mar–04 Apr | 12 Oct–9 Nov |
| 12 | BLDE | 10 Jun–08 Jul | 30 Aug–27 Sep |
| 13 | COMO | 05 Jul–02 Aug | 5 Sep–3 Oct |
| 13 | WLOU | 02 Jul–30 Jul | 3 Sep–1 Oct |
| 14 | SYCA | 12 Jan–11 Feb | 3 Jun–3 Jul |
| 15 | REDB | 29 Mar–26 Apr | 29 Sep–27 Oct |
| 16 | MCRA | 10 Apr–08 May | 23 Sep–21 Oct |
| 16 | MART | 06 Apr–04 May | 22 Sep–20 Oct |
| 17 | TECR | 06 May–17 Jun | 17 Sep–15 Oct |
| 17 | BIGC | 02 Apr–30 Apr | 28 Sep–26 Oct |
| 18 | OKSR | 21 May–18 Jun | 7 Aug–4 Sep |
| 19 | CARI | 02 May–30 May | 18 Aug–15 Sep |
| Site | N | Total Length (mm) | Wet Weight (g) |
|---|---|---|---|
| CARI | 186 | 144 (66 to 353) | 22.1 (3 to 361) |
| REDB | 242 | 142.5 (44 to 244) | 27.25 (1 to 140) |
| TECR | 876 | 123 (51 to 221) | 17 (0 to 95) |
| BLDE | 722 | 121 (65 to 215) | 17 (3 to 89) |
| BIGC | 2036 | 105 (31 to 215) | 11 (0 to 96) |
| WLOU | 840 | 104 (40 to 173) | 10.5 (0 to 52) |
| MCRA | 852 | 101 (41 to 158) | 8.75 (1 to 37) |
| MART | 1006 | 98 (57 to 154) | 7.9 (2 to 32) |
| LECO | 4456 | 75 (35 to 177) | 3.8 (0 to 53) |
| HOPB | 4144 | 62 (27 to 154) | 2.2 (0 to 32) |
| LEWI | 5641 | 59 (34 to 125) | 2.4 (0 to 20) |
| MCDI | 3488 | 55 (32 to 122) | 1.6 (0 to 21) |
| WALK | 4680 | 55 (23 to 91) | 1.4 (0 to 8) |
| MAYF | 417 | 52 (6 to 144) | 1.2 (0 to 35) |
| POSE | 6019 | 52 (24 to 89) | 1.3 (0 to 8) |
| OKSR | 180 | 50 (40 to 175) | 0.9 (0 to 39) |
| BLUE | 1519 | 45 (18 to 126) | 1.1 (0 to 25) |
| KING | 3652 | 42 (22 to 107) | 0.7 (0 to 12) |
| PRIN | 3861 | 42 (17 to 135) | 0.8 (0 to 36) |
| ARIK | 3201 | 41 (21 to 95) | 0.3 (0 to 10) |
| CUPE | 890 | 37 (14 to 220) | 0.6 (0 to 118) |
| GUIL | 1880 | 33 (13 to 82) | 0.3 (0 to 4) |
| SYCA | 2094 | 31 (17 to 67) | 0.3 (0 to 4) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Monahan, D.; Wesner, J.S.; Parker, S.M.; Schartel, H. Spatial Patterns in Fish Assemblages across the National Ecological Observation Network (NEON): The First Six Years. Fishes 2023, 8, 552. https://doi.org/10.3390/fishes8110552
Monahan D, Wesner JS, Parker SM, Schartel H. Spatial Patterns in Fish Assemblages across the National Ecological Observation Network (NEON): The First Six Years. Fishes. 2023; 8(11):552. https://doi.org/10.3390/fishes8110552
Chicago/Turabian StyleMonahan, Dylan, Jeff S. Wesner, Stephanie M. Parker, and Hannah Schartel. 2023. "Spatial Patterns in Fish Assemblages across the National Ecological Observation Network (NEON): The First Six Years" Fishes 8, no. 11: 552. https://doi.org/10.3390/fishes8110552
APA StyleMonahan, D., Wesner, J. S., Parker, S. M., & Schartel, H. (2023). Spatial Patterns in Fish Assemblages across the National Ecological Observation Network (NEON): The First Six Years. Fishes, 8(11), 552. https://doi.org/10.3390/fishes8110552