
Growth Performance and Histomorphology of Intestine, Skin, Gills and Liver of Juvenile Colossoma macropomum Fed Diets Containing Different Levels of the Essential Oil of Nectandra grandiflora


 , , , and
, , , and 
Abstract
1. Introduction
2. Materials and Methods
2.1. EO Extraction and Composition
2.2. Animal Maintenance and Handling
2.3. Diets and Treatments
2.4. Survival and Zootechnical Performance
- Weight (W) (g) = average weight;
- Daily weight gain (DWG) (g) = average weight gain (g)/time (days);
- Daily feed consumption per fish (DC) (g) = total feed consumption/experiment time (days)/number of animals per tank;
- Feed conversion ratio (FCR) = apparent total feed intake (g)/weight gain (g);
- Daily specific growth rate (SGR) (%) = 100 × (lnWf—lnWi)/interval between biometrics (days), where Wi is the initial weight, Wf is the final weight;
- Protein efficiency rate (PER) (%) = 100 × (weight gain (g)/protein consumed (g)).
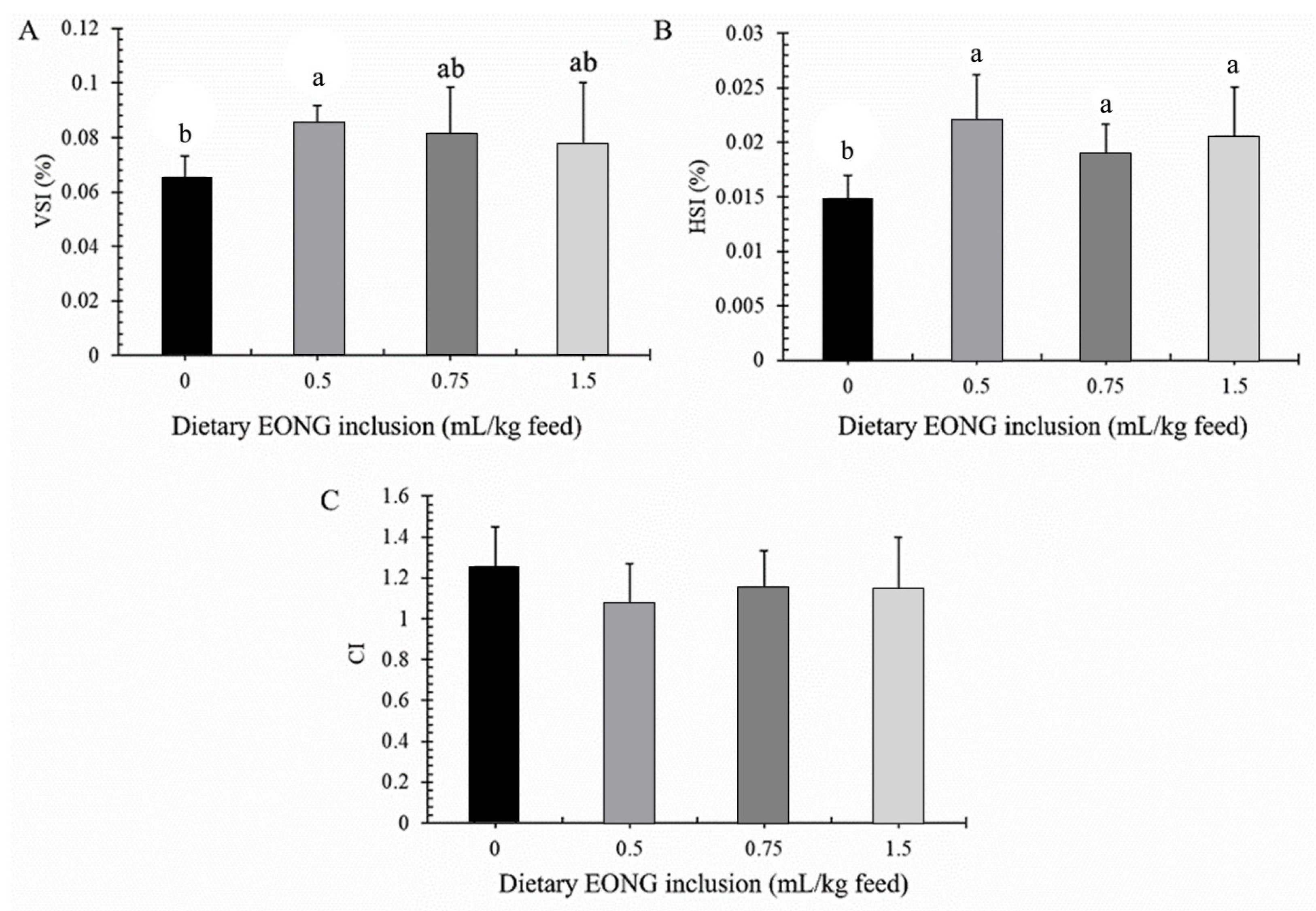
2.5. Hepatosomatic and Viscerosomatic Indices and Intestinal Coefficient
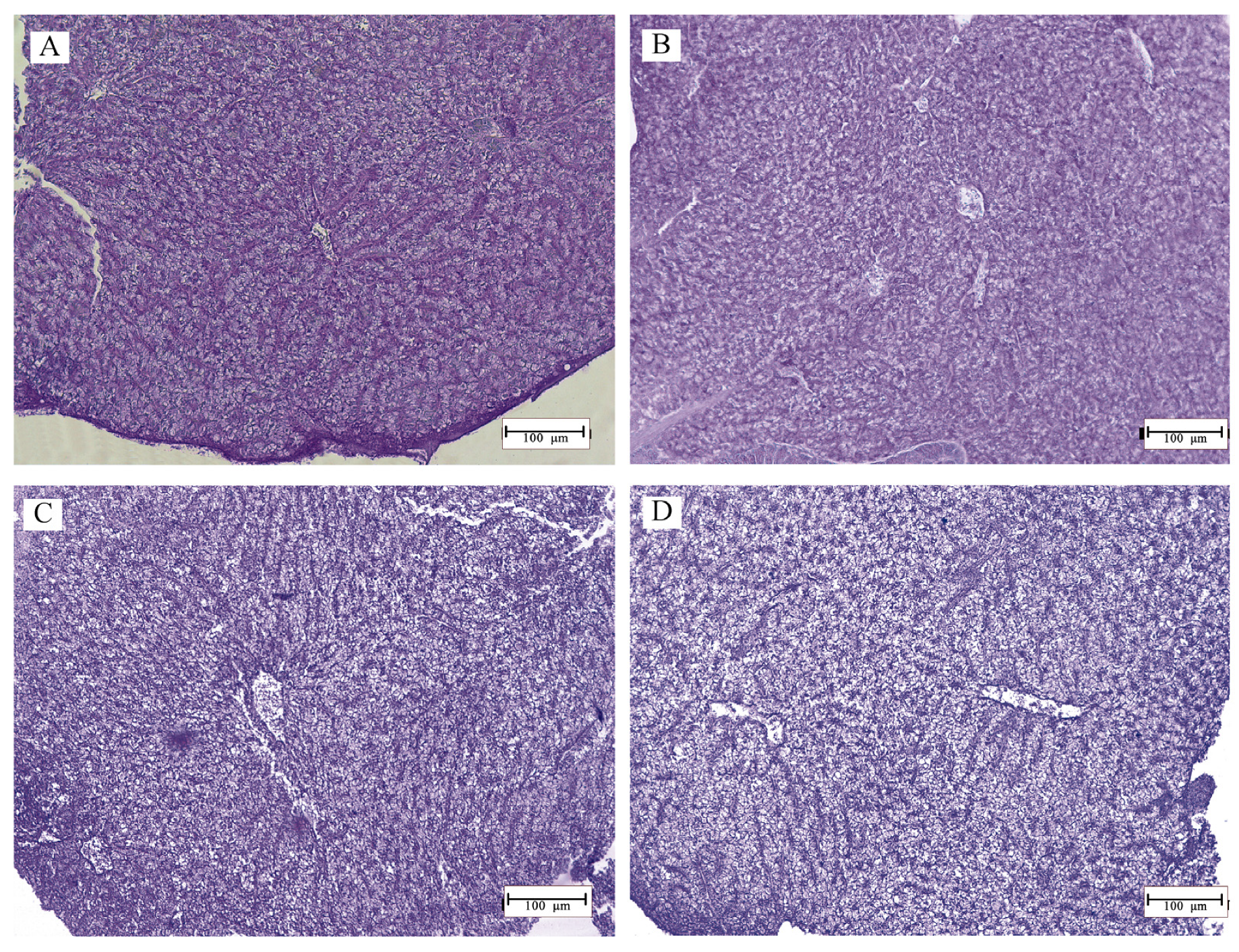
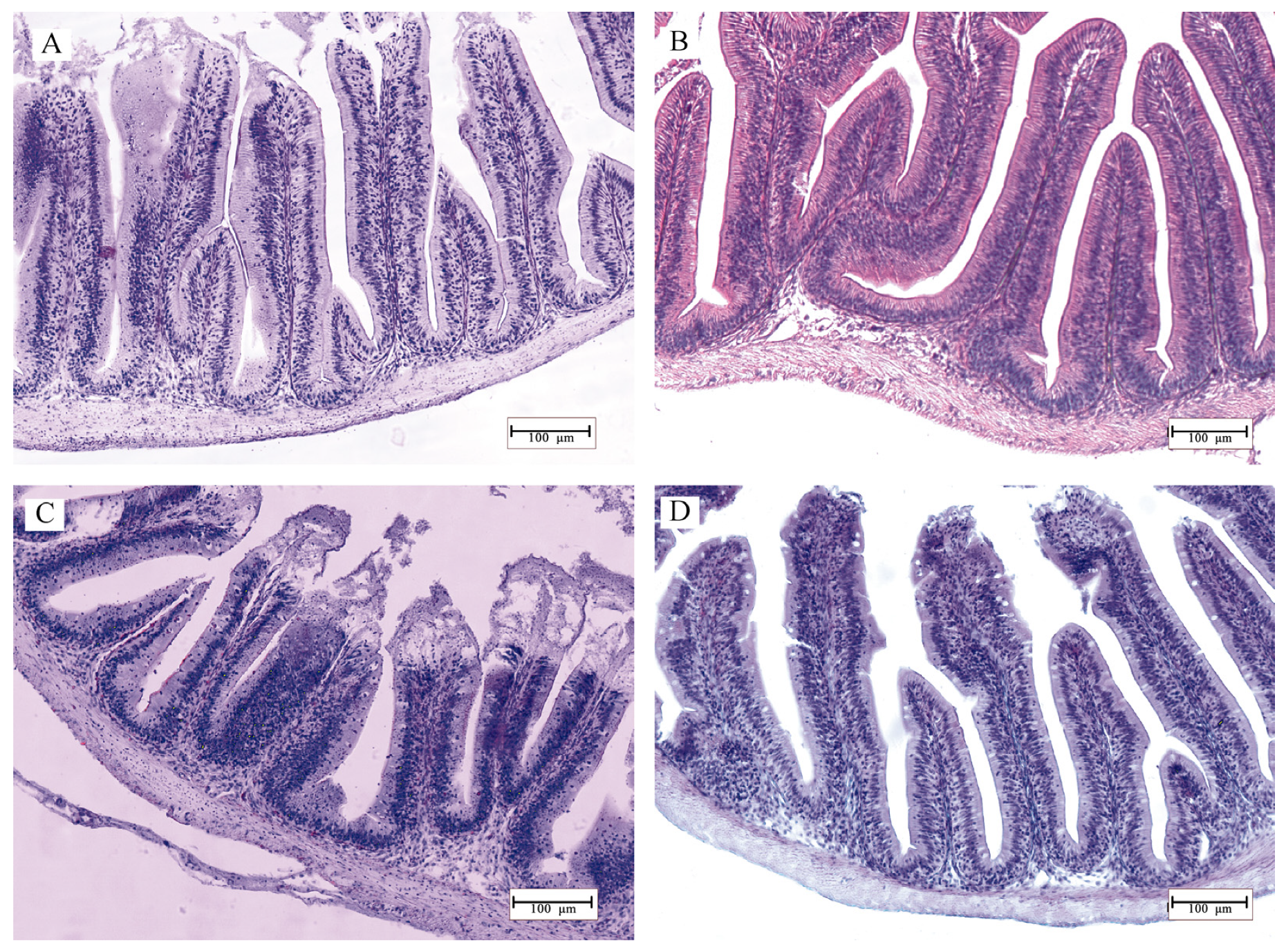
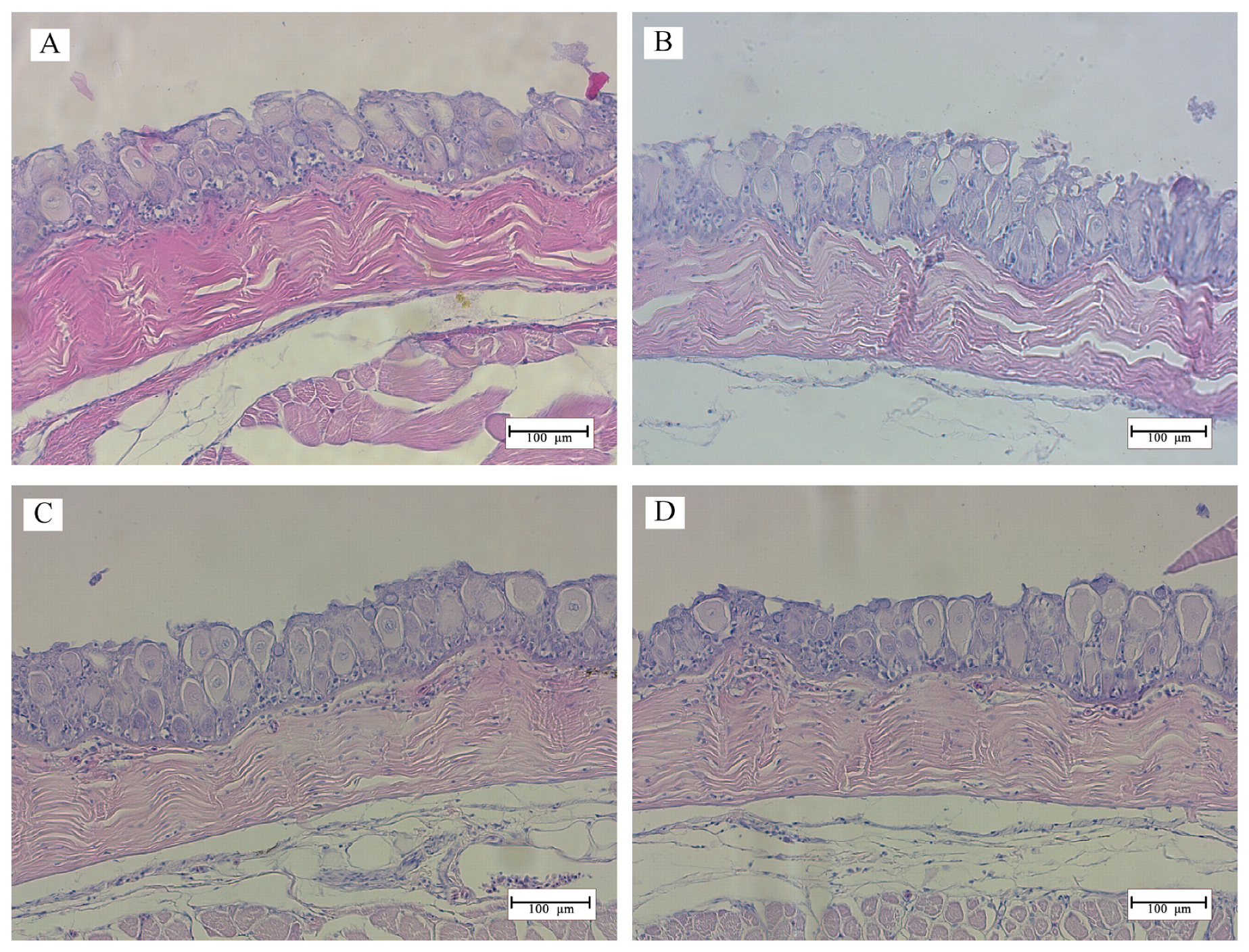
2.6. Histology
2.7. Statistical Analysis
3. Results
3.1. Performance
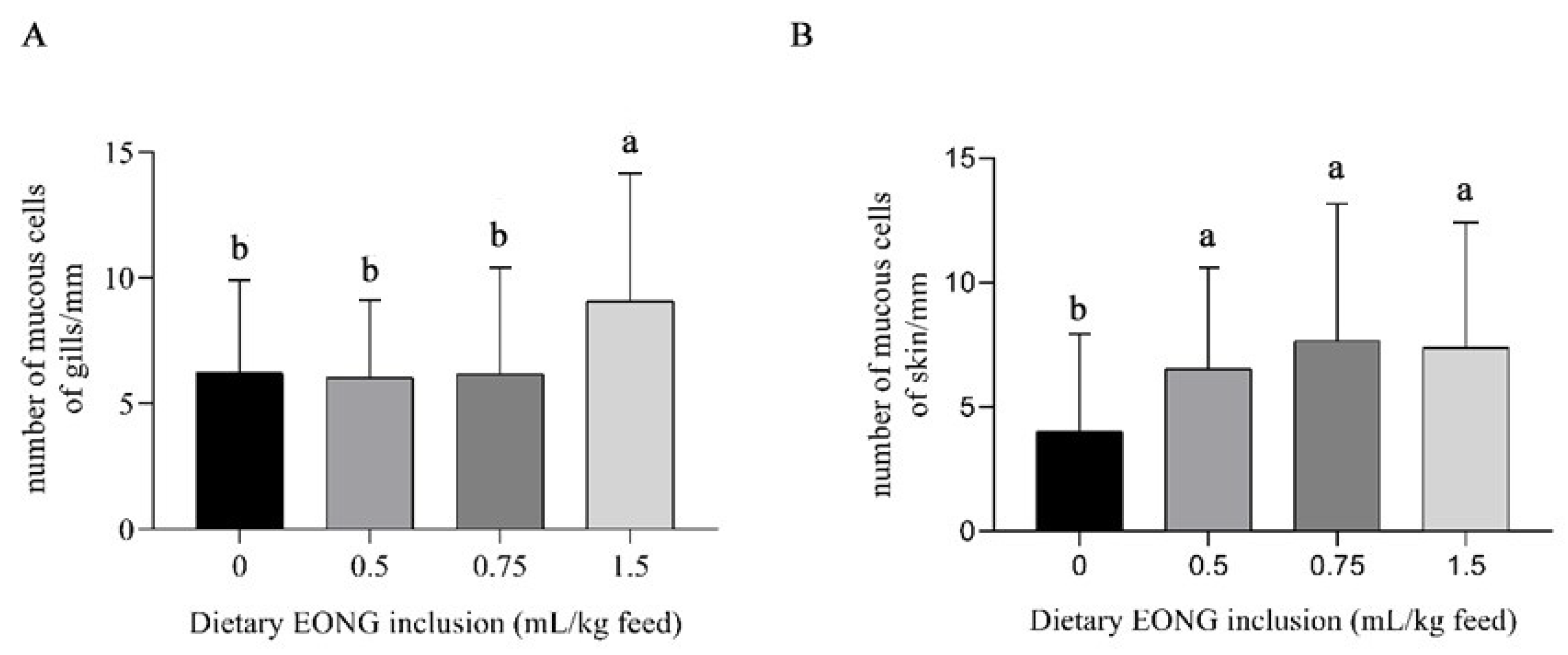
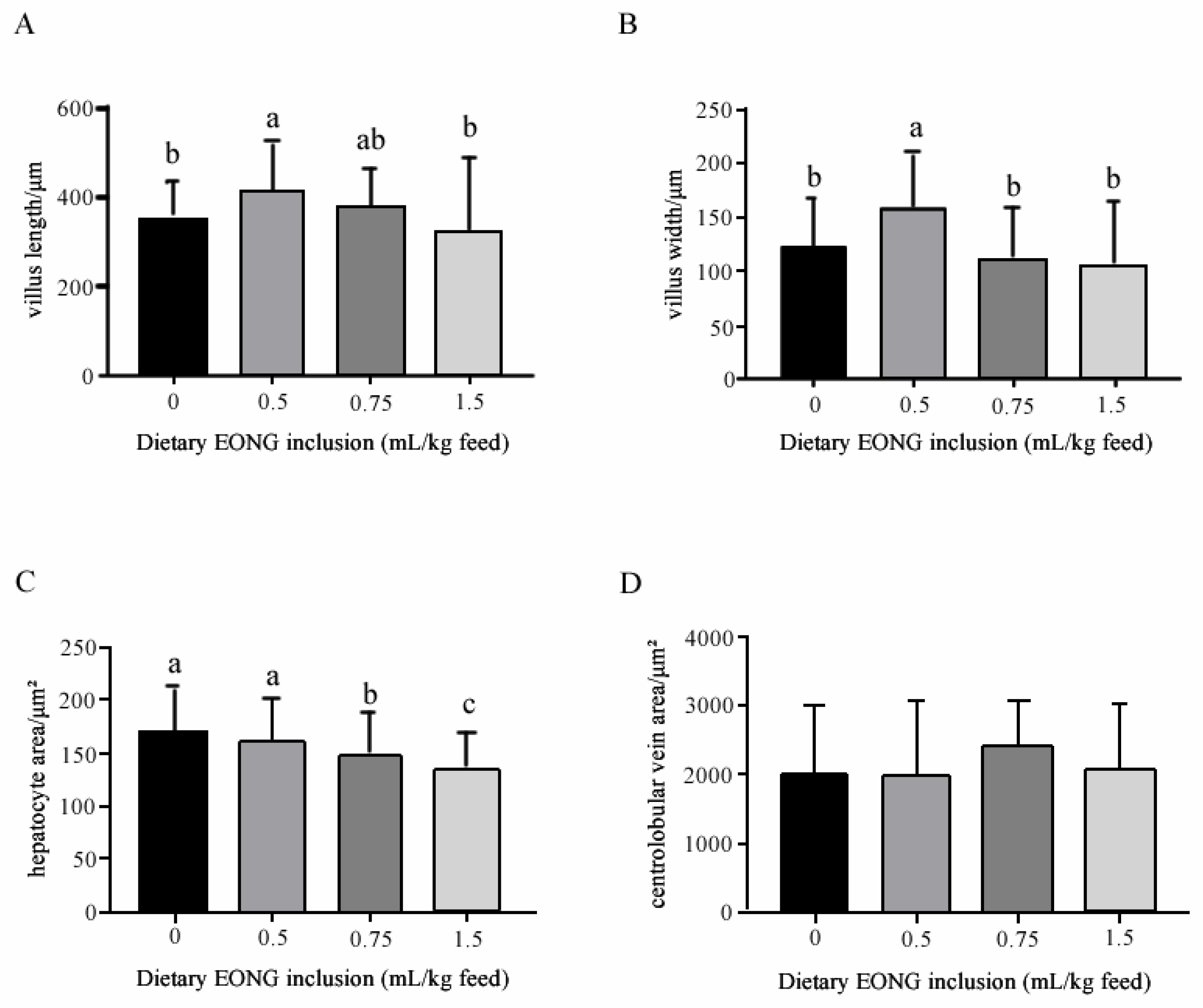
3.2. Histomorphology
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cristani, M.; D’Arrigo, M.; Mandalari, G.; Castelli, F.; Sarpietro, M.G.; Micieli, D.; Venuti, V.; Bisignano, G.; Saija, A.; Trombetta, D. Interaction of four monoterpenes contained in essential oils with model membranes: Implications for their antibacterial activity. J. Agric. Food Chem. 2007, 55, 6300–6308. [Google Scholar] [CrossRef]
- Nakatsu, T.; Lupo, A.T., Jr.; Chinn, J.W., Jr.; Kang, R.K. Biological activity of essential oils and their constituents. Stud. Nat. Prod. Chem. 2000, 21, 571–631. [Google Scholar]
- Ngugi, C.C.; Oyoo-Okoth, E.; Muchiri, M. Effects of dietary levels of essential oil (EO) extract from bitter lemon (Citrus limon) fruit peels on growth, biochemical, haemato-immunological parameters and disease resistance in Juvenile Labeo victorianus fingerlings challenged with Aeromonas hydrophila. Aquac. Res. 2016, 47, 872–885. [Google Scholar]
- Sutili, F.J.; Gatlin, D.M.; Heinzmann, B.M.; Baldisserotto, B. Plant essential oils as fish diet additives: Benefits on fish health and stability in feed. Rev. Aquac. 2017, 10, 716–726. [Google Scholar] [CrossRef]
- Al-Sagheer, A.A.; Mahmoud, H.K.; Reda, F.M.; Mahgoub, S.A.; Ayyat, M.S. Supplementation of diets for Oreochromis niloticus with essential oil extracts from lemongrass (Cymbopogon citratus) and geranium (Pelargonium graveolens) and effects on growth, intestinal microbiota, antioxidant and immune activities. Aquac. Nutr. 2018, 24, 1006–1014. [Google Scholar] [CrossRef]
- Amer, S.A.; Metwally, A.E.; Ahmed, S.A. The influence of dietary supplementation of cinnamaldehyde and thymol on the growth performance, immunity and antioxidant status of monosex Nile tilapia fingerlings (Oreochromis niloticus). Egypt. J. Aquat. Res. 2018, 44, 251–256. [Google Scholar] [CrossRef]
- Souza, C.D.F.; Baldissera, M.D.; Baldisserotto, B.; Heinzmann, B.M.; Martos-Sitcha, J.A.; Mancera, J.M. Essential oils as stress-reducing agents for fish aquaculture: A review. Front. Physiol. 2019, 10, 785. [Google Scholar] [CrossRef]
- Heluy, G.M.; Ramos, L.R.V.; Pedrosa, V.F.; Sarturi, C.; Figueiredo, P.G.P.; Vidal, L.G.P.; França, I.F.; Pereira, M.M. Oregano (Origanum vulgare) essential oil as an additive in diets for Nile tilapia (Oreochromis niloticus) fingerlings reared in salinized water. Aquac. Res. 2020, 51, 3237–3243. [Google Scholar] [CrossRef]
- Abdel-Latif, H.M.; Abdel-Tawwab, M.; Khafaga, A.F.; Dawood, M.A. Dietary oregano essential oil improved the growth performance via enhancing the intestinal morphometry and hepato-renal functions of common carp (Cyprinus carpio L.) fingerlings. Aquaculture 2020, 526, 735432. [Google Scholar] [CrossRef]
- Magouz, F.I.; Mahmoud, S.A.; El-Morsy, R.A.; Paray, B.A.; Soliman, A.A.; Zaineldin, A.I.; Dawood, M.A. Dietary menthol essential oil enhanced the growth performance, digestive enzyme activity, immune-related genes, and resistance against acute ammonia exposure in Nile tilapia (Oreochromis niloticus). Aquaculture 2021, 530, 735944. [Google Scholar] [CrossRef]
- Magouz, F.I.; Amer, A.A.; Faisal, A.; Sewilam, H.; Aboelenin, S.M.; Dawood, M.A. The effects of dietary oregano essential oil on the growth performance, intestinal health, immune, and antioxidative responses of Nile tilapia under acute heat stress. Aquaculture 2022, 548, 737632. [Google Scholar] [CrossRef]
- Ghafarifarsani, H.; Hoseinifar, S.H.; Javahery, S.; Van Doan, H. Effects of dietary vitamin C, thyme essential oil, and quercetin on the immunological and antioxidant status of common carp (Cyprinus carpio). Aquaculture 2022, 553, 738053. [Google Scholar] [CrossRef]
- Ghafarifarsani, H.; Hoseinifar, S.H.; Javahery, S.; Yazici, M.; Van Doan, H. Growth performance, biochemical parameters, and digestive enzymes in common carp (Cyprinus carpio) fed experimental diets supplemented with vitamin C, thyme essential oil, and quercetin. Ital. J. Anim. Sci. 2022, 21, 291–302. [Google Scholar] [CrossRef]
- Yousefi, M.; Ghafarifarsani, H.; Hoseini, S.M.; Hoseinifar, S.H.; Abtahi, B.; Vatnikov, Y.A.; Van Doan, H. Effects of dietary thyme essential oil and prebiotic administration on rainbow trout (Oncorhynchus mykiss) welfare and performance. Fish Shellfish Immunol. 2022, 120, 737–744. [Google Scholar] [CrossRef]
- Aguiar, G.A.C.C.D.; Carneiro, C.L.D.S.; Campelo, D.A.V.; Rusth, R.C.T.; Maciel, J.F.R.; Baldisserotto, B.; Salaro, A.L. Effects of Dietary Peppermint (Mentha piperita) Essential Oil on Growth Performance, Plasma Biochemistry, Digestive Enzyme Activity, and Oxidative Stress Responses in Juvenile Nile Tilapia (Oreochromis niloticus). Fishes 2023, 8, 374. [Google Scholar] [CrossRef]
- Viana, R.A.; Rapachi Fortes, C.; Rodrigues, P.; Ribeiro, S.; Maria Heinzmann, B.; Alves da Cunha, M.; Dellaméa Baldissera, M. Impact of dietary supplementation with Cinnamomum cassia essential oil on silver catfish (Rhamdia quelen): Toxicological effects and muscle antioxidant and fatty acid profiles. J. Toxicol. Environ. Health Part A 2023, 86, 313–325. [Google Scholar] [CrossRef]
- Farag, M.R.; Alagawany, M.; Khalil, S.R.; Abd El-Aziz, R.M.; Zaglool, A.W.; Moselhy, A.A.; Abou-Zeid, S.M. Effect of parsley essential oil on digestive enzymes, intestinal morphometry, blood chemistry and stress-related genes in liver of Nile tilapia fish exposed to Bifenthrin. Aquaculture 2022, 546, 737322. [Google Scholar] [CrossRef]
- Chung, S.; Ribeiro, K.; Teixeira, D.V.; Copatti, C.E. Inclusion of essential oil from ginger in the diet improves physiological parameters of tambaqui juveniles (Colossoma macropomum). Aquaculture 2021, 543, 736934. [Google Scholar] [CrossRef]
- Dawood, M.A.; El Basuini, M.F.; Zaineldin, A.I.; Yilmaz, S.; Hasan, M.T.; Ahmadifar, E.; Sewilam, H. Antiparasitic and antibacterial functionality of essential oils: An alternative approach for sustainable aquaculture. Pathogens 2021, 10, 185. [Google Scholar] [CrossRef]
- Mzengereza, K.; Ishikawa, M.; Koshio, S.; Yokoyama, S.; Yukun, Z.; Shadrack, R.S.; Dawood, M.A. Effect of substituting fish oil with camelina oil on growth performance, fatty acid profile, digestibility, liver histology, and antioxidative status of red seabream (Pagrus major). Animals 2021, 11, 1990. [Google Scholar] [CrossRef]
- Dawood, M.A.; Gewaily, M.S.; Sewilam, H. The growth performance, antioxidative capacity, and histological features of intestines, gills, and livers of Nile tilapia reared in different water salinities and fed menthol essential oil. Aquaculture 2022, 554, 738122. [Google Scholar] [CrossRef]
- Dawood, M.A.; El Basuini, M.F.; Yilmaz, S.; Abdel-Latif, H.M.; Alagawany, M.; Kari, Z.A.; Van Doan, H. Exploring the roles of dietary herbal essential oils in aquaculture: A review. Animals 2022, 12, 823. [Google Scholar] [CrossRef] [PubMed]
- Lorenzi, H. Árvores Brasileiras: Manual de Identificação e Cultivo de Plantas Arbóreas Nativas do Brasil, 2nd ed.; Plantarum: Nova Odessa, Brazil, 2002; Volume 2. [Google Scholar]
- NIST/EPA/NIH Mass Spectral Library & Search Program, 2nd ed.; John Wiley & Sons: Hoboken, NJ, USA, 2009.
- Rodrigues, P.; Ferrari, F.T.; Barbosa, L.B.; Righi, A.; Laporta, L.; Garlet, Q.I.; Heinzmann, B.M. Nanoemulsion Boosts Anesthetic Activity and Reduces the Side Effects of Nectandra Grandiflora Nees Essential Oil in Fish. Aquaculture 2021, 545, 737146. [Google Scholar] [CrossRef]
- Barbas, L.A.L.; Maltez, L.C.; Stringhetta, G.R.; de Oliveira Garcia, L.; Monserrat, J.M.; da Silva, D.T.; Sampaio, L.A. Properties of Two Plant Extractives as Anaesthetics and Antioxidants for Juvenile Tambaqui Colossoma Macropomum. Aquaculture 2017, 469, 79–87. [Google Scholar] [CrossRef]
- Garlet, Q.I.; Pires, L.C.; Silva, D.T.; Spall, S.; Gressler, L.T.; Bürger, M.E.; Heinzmann, B.M. Effect of (+)-Dehydrofukinone on GABA A Receptors and Stress Response in Fish Model. Braz. J. Med. Biol. Res. 2015, 49, e4872. [Google Scholar]
- Garlet, Q.I.; Souza, C.F.; Rodrigues, P.; Descovi, S.N.; Martinez-Rodríguez, G.; Baldisserotto, B.; Heinzmann, B.M. GABAa Receptor Subunits Expression in Silver Catfish (Rhamdia quelen) Brain and Its Modulation by Nectandra grandiflora Nees Essential Oil and Isolated Compounds. Behav. Brain Res. 2019, 376, 112178. [Google Scholar] [CrossRef]
- Araujo-Lima, C.A.R.M.; Goulding, M.; Forsberg, B.; Victoria, R.; Martinelli, L. The Economic Value of the Amazonian Flooded Forest from a Fisheries Perspective. Verh. Int. Verein. Limnol. 1998, 26, 2177–2179. [Google Scholar] [CrossRef]
- Mirande, J.M. Phylogeny of the Family Characidae (Teleostei: Characiformes): From Characters to Taxonomy. Neotrop. Ichthyol. 2010, 8, 385–568. [Google Scholar] [CrossRef]
- Baldisserotto, B. Capítulo do Tambaqui. In Espécies Nativas para Piscicultura no Brasil: 3a Edição Revista, Atualizada e Ampliada; Fundação de Apoio a Tecnologia e Ciencia-Editora UFSM: Santa Maria, Brazil, 2020. [Google Scholar]
- Peixe, B.R. Anuário de Piscicultura Brasileira; Associação Brasileira da Piscicultura: São Paulo, Brazil, 2023. [Google Scholar]
- Woynárovich, A.; Van Anrooy, R. Field Guide to the Culture of Tambaqui (Colossoma macropomum, Cuvier, 1816); FAO Fisheries and Aquaculture Technical Paper No. 624; FAO: Rome, Italy, 2019; 132p. [Google Scholar]
- Guimarães, I.G.; Miranda, E.C.; Araújo, J.G. Coefficients of Total Tract Apparent Digestibility of Some Feedstuffs for Tambaqui (Colossoma Macropomum). Anim. Feed Sci. Technol. 2014, 188, 150–155. [Google Scholar] [CrossRef]
- Santos, F.A.; Boaventura, T.P.; da Costa Julio, G.S.; Cortezzi, P.P.; Figueiredo, L.G.; Favero, G.C.; Luz, R.K. Growth Performance and Physiological Parameters of Colossoma macropomum in a Recirculating Aquaculture System (RAS): Importance of Stocking Density and Classification. Aquaculture 2021, 534, 736274. [Google Scholar] [CrossRef]
- Boijink, L.C.; Queiroz, C.A.; Chagas, E.C.; Chaves, F.C.M.; Inoue, L.A.K.A. Anesthetic and anthelminthic effects of clove basil (Ocimum gratissimum) essential oil for tambaqui (Colossoma macropomum). Aquaculture 2016, 457, 24–28. [Google Scholar] [CrossRef]
- Saccol, E.M.; Toni, C.; Pês, T.S.; Ourique, G.M.; Gressler, L.T.; Silva, L.V.; Mourão, R.H.V.; Oliveira, R.B.; Baldisserotto, B.; Pavanato, M.A. Anaesthetic and antioxidant effects of Myrcia sylvatica (G. Mey.) DC. and Curcuma longa L. essential oils on tambaqui (Colossoma macropomum). Aquac. Res. 2017, 48, 2012–2031. [Google Scholar] [CrossRef]
- Barbas, L.A.L.; Hamoy, M.; de Mello, V.J.; Barbosa, R.P.M.; de Lima, H.S.T.; Torres, M.F.; Nascimento, L.A.S.D.; da Silva, J.K.D.R.; de Aguiar Andrade, E.H.; Gomes, M.R.F. Essential oil of citronella modulates electrophysiological responses in tambaqui Colossoma macropomum: A new anaesthetic for use in fish. Aquaculture 2017, 479, 60–68. [Google Scholar] [CrossRef]
- Baldisserotto, B.; Barata, L.E.; Silva, A.S.; Lobato, W.F.; Silva, L.L.; Toni, C.; Silva, L.V. Anesthesia of tambaqui Colossoma macropomum (Characiformes: Serrasalmidae) with the essential oils of Aniba rosaeodora and Aniba parviflora and their major compound, linalool. Neotrop. Ichthyol. 2018, 16, e170128. [Google Scholar] [CrossRef]
- da Silva, H.N.P.; dos Santos Machado, S.D.; de Andrade Siqueira, A.M.; da Silva, E.C.C.; de Oliveira Canto, M.Â.; Jensen, L.; da Silva, L.V.F.; Fugimura, M.M.S.; Barroso, A.d.S.; Mourão, R.H.V.; et al. Sedative and anesthetic potential of the essential oil and hydrolate from the fruit of Protium heptaphyllum and their isolated compounds in Colossoma macropomum juveniles. Aquaculture 2020, 529, 735629. [Google Scholar] [CrossRef]
- dos Santos Maia, J.L.; de Oliveira Sousa, E.M.; da Silva, H.N.P.; Pinheiro, M.T.L.; Mourão, R.H.V.; Maia, J.G.S.; de Oliveira Coelho Lameirão, S.V.; Baldisserotto, B.; da Silva, L.V.F. Hydrolate toxicity of Lippia alba (Mill.) NE Brown (Verbenaceae) in juvenile tambaqui (Colossoma macropomum) and its potential anaesthetic properties. Aquaculture 2019, 503, 367–372. [Google Scholar] [CrossRef]
- Brandão, F.R.; Farias, C.F.S.; de Melo Souza, D.C.; de Oliveira, M.I.B.; de Matos, L.V.; Majolo, C.; de Oliveira, M.R.; Chaves, F.C.M.; de Almeida O’Sullivan, F.L.; Chagas, E.C. Anesthetic potential of the essential oils of Aloysia triphylla, Lippia sidoides and Mentha piperita for Colossoma macropomum. Aquaculture 2021, 534, 736275. [Google Scholar] [CrossRef]
- Boaventura, T.P.; dos Santos FA, C.; de Sena Souza, A.; Batista, F.S.; Júlio GS, C.; Luz, R.K. Thymol and linalool chemotypes of the essential oil of Thymus vulgaris as anesthetic for Colossoma macropomum: Physiology and feed consumption. Aquaculture 2022, 554, 738161. [Google Scholar] [CrossRef]
- Brandão, F.R.; Duncan, W.P.; Farias, C.F.S.; de Melo Souza, D.C.; de Oliveira, M.I.B.; Rocha, M.J.S.; Monteiro, P.C.; Majolo, C.; Chaves, F.C.M.; de Almeida O’Sullivan, F.L.; et al. Essential oils of Lippia sidoides and Mentha piperita as reducers of stress during the transport of Colossoma macropomum. Aquaculture 2022, 560, 738515. [Google Scholar] [CrossRef]
- Ferreira, A.L.; dos Santos, F.A.C.; Souza, A.d.S.; Favero, G.C.; Baldisserotto, B.; Pinheiro, C.G.; Heinzmann, B.M.; Luz, R.K. Efficacy of Hesperozygis ringens essential oil as an anesthetic and for sedation of juvenile tambaqui (Colossoma macropomum) during simulated transport. Aquacult. Int. 2022, 30, 1549–1561. [Google Scholar] [CrossRef]
- Monteiro, P.C.; Brandão, F.R.; Farias, C.F.S.; de Alexandre Sebastião, F.; Majolo, C.; Dairiki, J.K.; de Oliveira, M.R.; Chaves, F.C.M.; de Almeida O’Sullivan, F.L.; Martins, M.L.; et al. Dietary supplementation with essential oils of Lippia sidoides Ocimum gratissimum and Zingiber officinale on the growth and hemato-immunological parameters of Colossoma macropomum challenged with Aeromonas hydrophila. Aquac. Rep. 2021, 19, 100561. [Google Scholar] [CrossRef]
- Malheiros, D.F.; Videira, M.N.; Ferreira, I.M.; Tavares-Dias, M. Anthelmintic efficacy of Copaifera reticulata oleoresin in the control of monogeneans and haematological and histopathological effects on Colossoma macropomum. Aquac. Res. 2022, 53, 4087–4094. [Google Scholar] [CrossRef]
- Majolo, C.; Bizzo, H.R.; Rodrigues Brandão, F.; da Silva, A.M.S.; Campos Chagas, E.; Chaves, F.C.M.; Muniz, A.W. Chemical composition of Lippia Linn. (Verbenaceae) essential oils and their antibacterial potential against Aeromonas spp. isolates from Colossoma macropomum. J. Essent. Oil Res. 2022, 34, 251–261. [Google Scholar] [CrossRef]
- Queiroz, M.N.; dos Santos Torres, Z.E.; Pohlit, A.M.; Ono, E.A.; Affonso, E.G. Therapeutic potential of Piper aduncum leaf extract in the control of monogeneans in tambaqui (Colossoma macropomum). Aquaculture 2022, 552, 738024. [Google Scholar] [CrossRef]
- Silva, T.V.; Torres, M.F.; Sampaio, L.A.; Hamoy, M.; Monserrat, J.M.; Barbas, L.A.L. Dietary Euterpe oleracea Mart. attenuates seizures and damage to lipids in the brain of Colossoma macropomum. Fish Physiol. Biochem. 2021, 47, 1851–1864. [Google Scholar] [CrossRef]
- Copatti, C.E.; Felix e Silva, A.; Lorenzo, V.P.; da Costa, M.M.; Melo, J.F. Addition of essential oil from lemongrass to the tambaqui (Colossoma macropomum) diet: Effects on growth, intestinal enzymes, haematological and metabolic variables, and antimicrobial challenge. Aquac. Res. 2022, 53, 5656–5666. [Google Scholar] [CrossRef]
- Zeppenfeld, C.C.; Hernández, D.R.; Santinón, J.J.; Heinzmann, B.M.; Da Cunha, M.A.; Schmidt, D.; Baldisserotto, B. Essential oil of Aloysia triphylla as feed additive promotes growth of silver catfish (Rhamdia quelen). Aquac. Nutr. 2016, 22, 933–940. [Google Scholar] [CrossRef]
- Association of Official Analytical Chemists (AOAC). Official Methods of Analysis, 21st ed.; AOAC International: Gaithersburg, MD, USA, 2019. [Google Scholar]
- Kozloski, G.V.; Zilio, E.M.C.; Ongarato, F.; Kuinchtner, B.C.; Saccol, A.G.; Genro, T.C.M.; Quadros, F.L.F. Faecal N excretion as an approach for estimating organic matter intake by free-ranging sheep and cattle—CORRIGENDUM. J. Agric. Sci. 2018, 156, 855. [Google Scholar] [CrossRef]
- Mattioli, C.C.; Takata, R.; Leme, F.D.O.P.; Costa, D.C.; Melillo Filho, R.; e Silva, W.D.S.; Luz, R.K. The effects of acute and chronic exposure to water salinity on juveniles of the carnivorous freshwater catfish Lophiosilurus alexandri. Aquaculture 2017, 481, 255–266. [Google Scholar] [CrossRef]
- Guardiola, F.A.; Cuesta, A.; Esteban, M.A. Using skin mucus to evaluate stress in gilthead seabream (Sparus aurata L.). Fish Shellfish Immunol. 2016, 59, 323–330. [Google Scholar] [CrossRef]
- Sotoudeh, A.; Yeganeh, S. Effects of Supplementary Fennel (Foeniculum vulgare) Essential Oil in Diet on Growth and Reproductive Performance of the Ornamental Fish Convict Cichlid (Cichlasoma nigrofasciatum). Aquac. Res. 2017, 48, 4284–4291. [Google Scholar] [CrossRef]
- Souza, E.M.; de Souza, R.C.; Melo, J.F.; da Costa, M.M.; de Souza, A.M.; Copatti, C.E. Evaluation of the Effects of Ocimum basilicum Essential Oil in Nile Tilapia Diet: Growth Biochemical Intestinal Enzymes Haematology Lysozyme and Antimicrobial Challenges. Fishes 2019, 504, 7–12. [Google Scholar] [CrossRef]
- Shourbela, R.M.; El-Hawarry, W.N.; Elfadadny, M.R.; Dawood, M.A.O. Growth Performance and Antioxidant Enzyme Activities in Nile Tilapia (Oreochromis niloticus) Reared Under Intensive Systems Supplemented with Thyme Essential Oil in Diet as a Natural Growth Promoter. Fishes 2021, 542, 736868. [Google Scholar]
- Ebrahimi, E.; Haghjou, M.; Nematollahi, A.; Goudarzian, F. Effects of Rosemary Essential Oil on Growth Performance and Hematological Parameters of Beluga (Huso huso). Fishes 2020, 521, 734909. [Google Scholar]
- Sönmez, A.Y.; Bilen, S.; Alak, G.; Hisar, O.; Yanık, T.; Biswas, G. Growth Performance and Antioxidant Enzyme Activities in Rainbow Trout (Oncorhynchus mykiss) Juveniles Fed Diets Supplemented with Sage, Mint, and Thyme Oils. Fishes 2015, 41, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Bandaranayake, W.M. Quality Control, Screening, Toxicity, and Regulation of Herbal Drugs. In Modern Phytomedicine: Turning Medicinal Plants into Drugs; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2006; pp. 25–57. [Google Scholar]
- Piazzon, M.C.; Naya-Català, F.; Pereira, G.V.; Estensoro, I.; Del Pozo, R.; Calduch-Giner, J.A.; Pérez-Sánchez, J. A Novel Fish Meal-Free Diet Formulation Supports Proper Growth and Does Not Impair Intestinal Parasite Susceptibility in Gilthead Sea Bream (Sparus aurata) with a Reshape of Gut Microbiota and Tissue-Specific Gene Expression Patterns. Aquaculture 2022, 558, 738362. [Google Scholar] [CrossRef]
- Zeng, Z.; Zhang, S.; Wang, H.; Piao, X. Essential Oils and Aromatic Plants as Feed Additives in Non-Ruminant Nutrition: A Review. J. Anim. Sci. Biotechnol. 2015, 6, 7. [Google Scholar] [CrossRef] [PubMed]
- Laparra, J.M.; Sanz, Y. Interactions of Gut Microbiota with Functional Food Components and Nutraceuticals. Pharmacol. Res. 2010, 61, 219–225. [Google Scholar] [CrossRef]
- Ni, J.; Yan, Q.; Yu, Y.; Zhang, T. Fish gut microecosystem: A model for detecting spatial pattern of microorganisms. Chin. J. Oceanol. Limnol. 2014, 32, 54–57. [Google Scholar] [CrossRef][Green Version]
- Lazado, C.C.; Caipang, C.M.A. Mucosal immunity and probiotics in fish. Fish Shellfish Immunol. 2014, 39, 78–89. [Google Scholar] [CrossRef]
- Deng, Y.; Zhang, Y.; Chen, H.; Xu, L.; Wang, Q.; Feng, J. Gut–liver immune response and gut microbiota profiling reveal the pathogenic mechanisms of vibrio harveyi in pearl gentian grouper (Epinephelus lanceolatus♂ × E. fuscoguttatus♀). Front. Immunol. 2020, 11, 607754. [Google Scholar] [CrossRef]
- Acar, Ü.; Kesbiç, O.S.; Yılmaz, S.; Gültepe, N.; Türker, A. Evaluation of the Effects of Essential Oil Extracted from Sweet Orange Peel (Citrus sinensis) on Growth Rate of Tilapia (Oreochromis mossambicus) and Possible Disease Resistance against Streptococcus iniae. Aquaculture 2015, 437, 282–286. [Google Scholar] [CrossRef]
- Diler, O.; Gormez, O.; Diler, I.; Metin, S. Effect of Oregano (Origanum onites L.) Essential Oil on Growth Lysozyme and Antioxidant Activity and Resistance Against Lactococcus garvieae in Rainbow Trout Oncorhynchus mykiss (Walbaum). Aquac. Nutr. 2017, 23, 844–851. [Google Scholar] [CrossRef]
- Brunner, S.R.; Varga, J.F.; Dixon, B. Antimicrobial Peptides of Salmonid Fish: From Form to Function. Biology 2020, 9, 233. [Google Scholar] [CrossRef] [PubMed]
- Magnadóttir, B. Innate Immunity of Fish (Overview). Fishes 2006, 20, 137–151. [Google Scholar] [CrossRef]
- Cain, K.; Swan, C. Barrier Function and Immunology. In Fish Physiology; Academic Press: Cambridge, MA, USA, 2010; Volume 30, pp. 111–134. [Google Scholar]
- Kiron, V. Fish Immune System and Its Nutritional Modulation for Preventive Health Care. Anim. Feed Sci. Technol. 2012, 173, 111–133. [Google Scholar] [CrossRef]
- Wang, S.; Wang, Y.; Ma, J.; Ding, Y.; Zhang, S. Phosvitin Plays a Critical Role in the Immunity of Zebrafish Embryos via Acting as a Pattern Recognition Receptor and an Antimicrobial Effector. J. Biol. Chem. 2011, 286, 22653–22664. [Google Scholar] [CrossRef]
- Guardiola, F.A.; Cuesta, A.; Abellán, E.; Meseguer, J.; Esteban, M.A. Comparative Analysis of the Humoral Immunity of Skin Mucus from Several Marine Teleost Fish. Fish Shellfish Immunol. 2014, 40, 24–31. [Google Scholar] [CrossRef]
- Guardiola, F.A.; Dioguardi, M.; Parisi, M.G.; Trapani, M.R.; Meseguer, J.; Cuesta, A.; Esteban, M.A. Evaluation of Waterborne Exposure to Heavy Metals in Innate Immune Defenses Present on Skin Mucus of Gilthead Seabream (Sparus aurata). Fish Shellfish Immunol. 2015, 45, 112–123. [Google Scholar] [CrossRef] [PubMed]
- Cerezuela, R.; Guardiola, F.A.; Cuesta, A.; Esteban, M.A. Enrichment of Gilthead Seabream (Sparus aurata L.) Diet with Palm Fruit Extracts and Probiotics: Effects on Skin Mucosal Immunity. Fish Shellfish Immunol. 2016, 49, 100–109. [Google Scholar] [CrossRef]
- Reverter, M.; Tapissier-Bontemps, N.; Lecchini, D.; Banaigs, B.; Sasal, P. Biological and Ecological Roles of External Fish Mucus: A Review. Fishes 2018, 3, 41. [Google Scholar] [CrossRef]
- Santoso, H.B.; Suhartono, E.; Yunita, R.; Biyatmoko, D. Epidermal Mucus as a Potential Biological Matrix for Fish Health Analysis. Egypt. J. Aquat. Biol. Fish. 2020, 24, 361–382. [Google Scholar] [CrossRef]
- Aranishi, F.; Mano, N. Response of Skin Cathepsins to Infection of Edwardsiella tarda in Japanese Flounder. J. Aquat. Anim. Health 2000, 66, 169–170. [Google Scholar] [CrossRef]
- Concha, M.I.; Molina, S.; Oyarzún, C.; Villanueva, J.; Amthauer, R. Local Expression of Apolipoprotein AI Gene and a Possible Role for HDL in Primary Defense in the Carp Skin. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2003, 14, 259–273. [Google Scholar]
- Firth, K.J.; Johnson, S.C.; Ross, N.W. Characterization of Proteases in the Skin Mucus of Atlantic Salmon (Salmo salar) Infected with the Salmon Louse (Lepeophtheirus salmonis) and in Whole-Body Louse Homogenate. J. Aquat. Anim. Health 2000, 86, 1199–1205. [Google Scholar]
- Hatten, F.; Fredriksen, Å.; Hordvik, I.; Endresen, C. Presence of IgM in Cutaneous Mucus, but Not in Gut Mucus of Atlantic Salmon, Salmo salar. Serum IgM Is Rapidly Degraded When Added to Gut Mucus. Dev. Comp. Immunol. 2001, 11, 257–268. [Google Scholar] [CrossRef] [PubMed]
- Núñez-Acuña, G.; Marambio, J.P.; Valenzuela, T.; Wadsworth, S.; Gallardo-Escárate, C. Antimicrobial Peptides from Salmon Salar Skin Induce Frontal Filament Development and Olfactory/Cuticle-Related Genes in the Sea Louse Caligus rogercresseyi. Aquaculture 2016, 464, 171–177. [Google Scholar] [CrossRef]
- Núñez-Acuña, G.; Gallardo-Escárate, C.; Fields, D.M.; Shema, S.; Skiftesvik, A.B.; Ormazábal, I.; Browman, H.I. The Atlantic Salmon (Salmo salar) Antimicrobial Peptide Cathelicidin-2 Is a Molecular Host-Associated Cue for the Salmon Louse (Lepeophtheirus salmonis). Sci. Rep. 2018, 8, 13738. [Google Scholar] [CrossRef]
- Firmino, J.P.; Vallejos-Vidal, E.; Sarasquete, C.; Ortiz-Delgado, J.B.; Balasch, J.C.; Tort, L.; Gisbert, E. Unveiling the Effect of Dietary Essential Oils Supplementation in Sparus aurata Gills and Its Efficiency Against the Infestation by Sparicotyle chrysophrii. Sci. Rep. 2020, 10, 17764. [Google Scholar] [CrossRef]
- Firmino, J.P.; Fernández-Alacid, L.; Vallejos-Vidal, E.; Salomón, R.; Sanahuja, I.; Tort, L.; Gisbert, E. Carvacrol, Thymol, and Garlic Essential Oil Promote Skin Innate Immunity in Gilthead Seabream (Sparus aurata) Through the Multifactorial Modulation of the Secretory Pathway and Enhancement of Mucus Protective Capacity. Front. Immunol. 2021, 12, 633621. [Google Scholar] [CrossRef]
- Sklan, D.; Prag, T.; Lupatsch, I. Structure and Function of the Small Intestine of the Tilapia Oreochromis niloticus × Oreochromis aureus (Teleostei, Cichlidae). Fish Physiol. Biochem. 2004, 35, 350–357. [Google Scholar] [CrossRef]
- Huerta-Aguirre, G.; Paredes-Ramos, K.M.; Becerra-Amezcua, M.P.; Hernández-Calderas, I.; Matadamas-Guzman, M.; Guzmán-García, X. Histopathological Analysis of the Intestine from Mugil cephalus on Environment Reference Sites. In Pollution of Water Bodies in Latin America: Impact of Contaminants on Species of Ecological Interest; MDPI: Basel, Switzerland, 2019; pp. 319–328. [Google Scholar]
- Lopes, J.M.; Marques, N.C.; dos Santos, M.D.D.M.C.; Souza, C.F.; Baldissera, M.D.; Carvalho, R.C.; Baldisserotto, B. Dietary Limon (Citrus × latifolia) Fruit Peel Essential Oil Improves Antioxidant Capacity of Tambaqui (Colossoma macropomum) Juveniles. Animals 2020, 51, 4852–4862. [Google Scholar]
- Valladão, G.M.R.; Gallani, S.U.; Jerônimo, G.T.; Seixas, A.T.D. Challenges in the Control of Acanthocephalosis in Aquaculture: Special Emphasis on Neoechinorhynchus buttnerae. Parasites Vectors 2020, 12, 1360–1372. [Google Scholar] [CrossRef]
- Adeoye, A.A.; Yomla, R.; Jaramillo-Torres, A.; Rodiles, A.; Merrifield, D.L.; Davies, S.J. Combined Effects of Exogenous Enzymes and Probiotic on Nile Tilapia (Oreochromis niloticus) Growth, Intestinal Morphology and Microbiome. Aquaculture 2016, 463, 61–70. [Google Scholar] [CrossRef]
- Caballero, M.J.; Izquierdo, M.S.; Kjørsvik, E.; Fernandez, A.J.; Rosenlund, G. Histological Alterations in the Liver of Sea Bream, Sparus aurata L., Caused by Short- or Long-term Feeding with Vegetable Oils. Recovery of Normal Morphology after Feeding Fish Oil as the Sole Lipid Source. Aquaculture 2004, 27, 531–541. [Google Scholar] [CrossRef]
- Ferreira, P.M.; Caldas, D.W.; Salaro, A.L.; Sartori, S.S.; Oliveira, J.M.; Cardoso, A.J.; Zuanon, J.A. Intestinal and Liver Morphometry of the Yellow Tail Tetra (Astyanax altiparanae) Fed with Oregano Oil. Aquaculture 2016, 88, 911–922. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients | (%) |
|---|---|
| Soybean meal | 35.00 |
| Fish meal | 30.00 |
| Rice bran | 12.00 |
| Corn | 15.00 |
| Canola oil | 3.00 |
| Salt | 1.00 |
| Vitamin and mineral premix 1 | 3.00 |
| Phosphate dicalcium | 1.00 |
| Analyzed proximate composition | |
| Dry matter content | 93.68 |
| Protein | 35.33 |
| Ether extract | 7.13 |
| Mineral matter | 22.77 |
| Acid detergent fiber | 1.40 |
| Neutral detergent fiber | 19.62 |
| EONG (mL/kg Feed) | 0.00 | 0.50 | 0.75 | 1.50 | p-Value |
|---|---|---|---|---|---|
| Day 10 | |||||
| W (g) | 2.26 ± 0.09 | 2.41 ± 0.02 | 2.22 ± 0.15 | 2.35 ± 0.08 | 0.0593 |
| DWG (g) | 0.15 ± 0.01 | 0.17 ± 0.00 | 0.15 ± 0.02 | 0.16 ± 0.01 | 0.0628 |
| DC (g) | 0.07 ± 0.0002 | 0.08 ± 0.0004 | 0.08 ± 0.0006 | 0.08 ± 0.0007 | 0.5524 |
| FCR | 0.5 ± 0.028 | 0.45 ± 0.008 | 0.52 ± 0.055 | 0.47 ± 0.021 | 0.0757 |
| SGR (%) | 11.05 ± 0.38 | 11.66 ± 0.12 | 10.8 ± 0.71 | 11.37 ± 0.30 | 0.0737 |
| PER (%) | 0.58 ± 0.03 | 0.63 ± 0.01 | 0.56 ± 0.06 | 0.60 ± 0.03 | 0.0730 |
| Survival (%) | 100 | 100 | 100 | 100 | |
| Day 20 | |||||
| W (g) | 3.88 ± 0.14 b | 5.04 ± 0.40 a | 4.35 ± 0.39 ab | 4.32 ± 0.49 ab | 0.0078 |
| DWG (g) | 0.16 ± 0.02 b | 0.26 ± 0.04 a | 0.21 ± 0.04 ab | 0.20 ± 0.04 ab | 0.0146 |
| DC (g) | 0.23 ± 0.01 b | 0.26 ± 0.01 a | 0.23 ± 0.02 ab | 0.24 ± 0.01 ab | 0.0424 |
| FCR | 1.44 ± 0.16 | 0.99 ± 0.17 | 1.12 ± 0.23 | 1.25 ± 0.29 | 0.0721 |
| SGR (%) | 5.41 ± 0.67 b | 7.35 ± 0.84 a | 6.72 ± 0.90 ab | 6.04 ± 0.91 ab | 0.0333 |
| PER (%) | 0.20 ± 0.02 | 0.29 ± 0.05 | 0.26 ± 0.06 | 0.24 ± 0.06 | 0.1283 |
| Survival (%) | 99.11 ± 1.79 | 100.00 ± 0.00 | 95.24 ± 5.46 | 99.1 ± 1.79 | 0.1665 |
| Day 30 | |||||
| W (g) | 5.64 ± 0.22 b | 7.49 ± 0.52 a | 6.58 ± 0.88 ab | 6.63 ± 0.69 ab | 0.0421 |
| DWG (g) | 0.18 ± 0.02 | 0.25 ± 0.03 | 0.22 ± 0.05 | 0.23 ± 0.11 | 0.4884 |
| DC (g) | 0.35 ± 0.01 c | 0.46 ± 0.00 a | 0.41 ± 0.01 b | 0.42 ± 0.01 b | <0.001 |
| FCR | 1.99 ± 0.20 | 1.90 ± 0.20 | 1.94 ± 0.55 | 2.21 ± 1.10 | 0.8990 |
| SGR (%) | 3.73 ± 0.42 | 3.97 ± 0.40 | 4.12 ± 0.53 | 4.12 ± 1.36 | 0.8956 |
| PER (%) | 0.14 ± 0.01 | 0.15 ± 0.02 | 0.15 ± 0.04 | 0.16 ± 0.08 | 0.9802 |
| Survival (%) | 94.64 ± 4.61 | 94.64 ± 4.61 | 94.64 ± 4.61 | 94.64 ± 4.61 | >0.999 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santos, F.A.C.; Batista, F.S.; Souza, A.S.; Julio, G.S.C.; Favero, G.C.; Junior, J.F.V.; Costa, S.T.; Zeppenfeld, C.C.; Bianchini, N.H.; Heinzmann, B.M.; et al. Growth Performance and Histomorphology of Intestine, Skin, Gills and Liver of Juvenile Colossoma macropomum Fed Diets Containing Different Levels of the Essential Oil of Nectandra grandiflora. Fishes 2023, 8, 509. https://doi.org/10.3390/fishes8100509
Santos FAC, Batista FS, Souza AS, Julio GSC, Favero GC, Junior JFV, Costa ST, Zeppenfeld CC, Bianchini NH, Heinzmann BM, et al. Growth Performance and Histomorphology of Intestine, Skin, Gills and Liver of Juvenile Colossoma macropomum Fed Diets Containing Different Levels of the Essential Oil of Nectandra grandiflora. Fishes. 2023; 8(10):509. https://doi.org/10.3390/fishes8100509
Chicago/Turabian StyleSantos, Fabio A. C., Felipe S. Batista, André S. Souza, Gustavo S. C. Julio, Gisele C. Favero, José F. V. Junior, Sílvio T. Costa, Carla C. Zeppenfeld, Nadia H. Bianchini, Berta M. Heinzmann, and et al. 2023. "Growth Performance and Histomorphology of Intestine, Skin, Gills and Liver of Juvenile Colossoma macropomum Fed Diets Containing Different Levels of the Essential Oil of Nectandra grandiflora" Fishes 8, no. 10: 509. https://doi.org/10.3390/fishes8100509
APA StyleSantos, F. A. C., Batista, F. S., Souza, A. S., Julio, G. S. C., Favero, G. C., Junior, J. F. V., Costa, S. T., Zeppenfeld, C. C., Bianchini, N. H., Heinzmann, B. M., Baldisserotto, B., & Luz, R. K. (2023). Growth Performance and Histomorphology of Intestine, Skin, Gills and Liver of Juvenile Colossoma macropomum Fed Diets Containing Different Levels of the Essential Oil of Nectandra grandiflora. Fishes, 8(10), 509. https://doi.org/10.3390/fishes8100509