
Towards the Control of the Reproduction of the Yellow Clam Amarilladesma mactroides (Reeve, 1854) in Captivity: Effects of Different Stimuli on the Spawning of Laboratory-Conditioned and Unconditioned Breeders

 ,
, 
Abstract
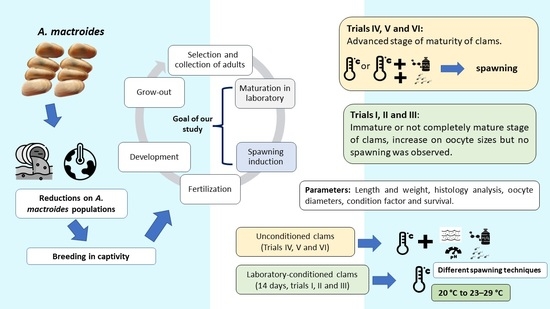
1. Introduction
2. Materials and Methods
2.1. Trials with Laboratory-Conditioned Clams
2.2. Trials with Laboratory-Unconditioned Clams
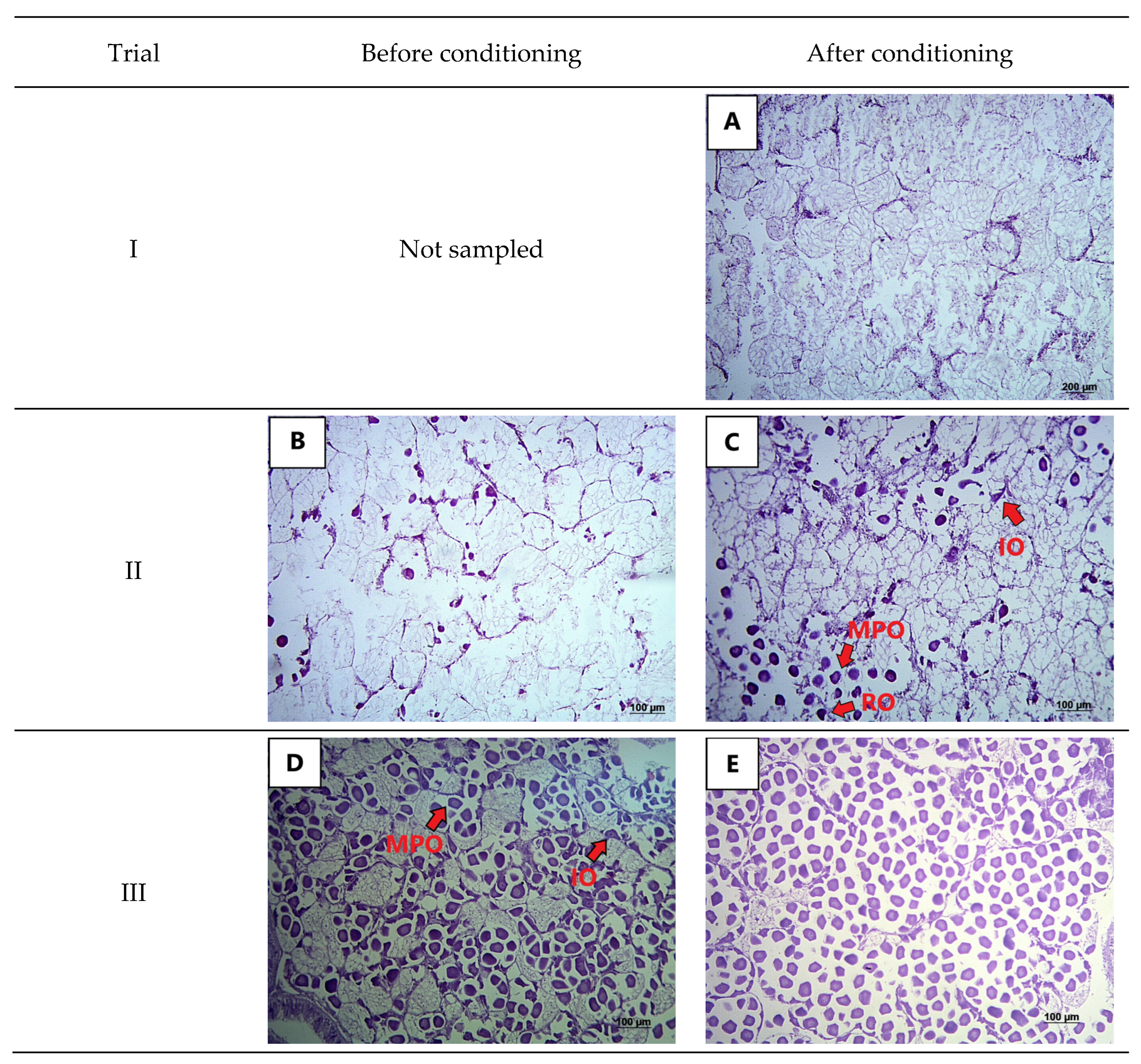
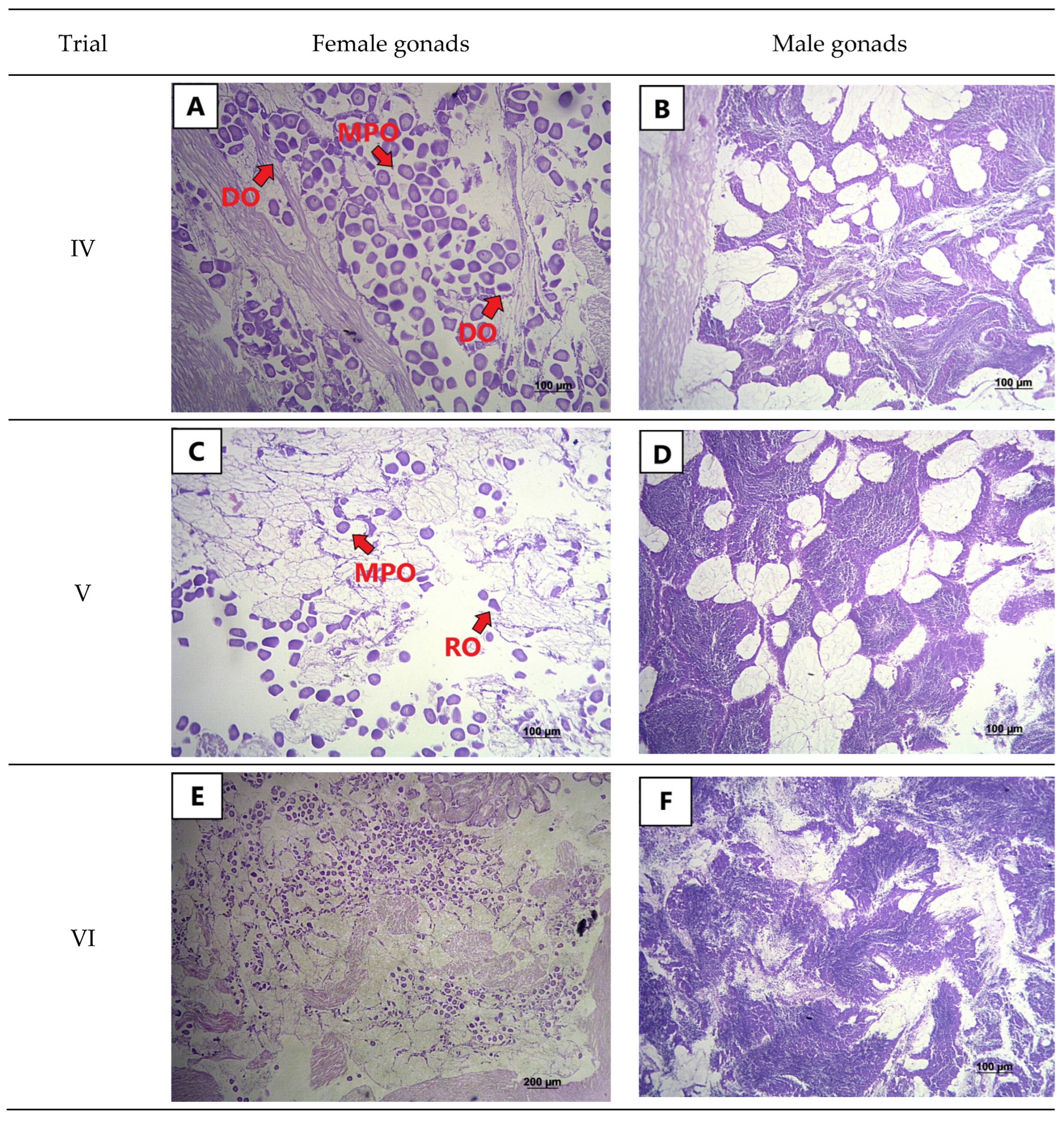
2.3. Histological Analysis
2.4. Statistical Analysis
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Coscarón, S. La almeja amarilla (Mesodesma mactroides, Deshayes) de la costa de la Provincia de Buenos Aires. Agro. Publ. Tec. 1959, 1, 1–66. [Google Scholar]
- Rios, E.C. Seashells of Brazil, 1st ed.; Fundação Universidade do Rio Grande: Rio Grande, Brazil, 1994; 368p. [Google Scholar]
- Fiori, S.M.; Morsán, E.M. Age and individual growth of Mesodesma mactroides (Bivalvia) in the southernmost range of its distribution. ICES J. Mar. Sci. 2004, 61, 1253–1259. [Google Scholar] [CrossRef]
- Herrmann, M.; Alfaya, J.E.F.; Lepore, M.L.; Penchaszadeh, P.E.; Laudien, J. Reproductive cycle and gonad development of the northern Argentinean Mesodesma mactroides (Bivalvia: Mesodesmatidae). Helgol. Mar. Res. 2009, 63, 207–218. [Google Scholar] [CrossRef]
- McLachlan, A. Fisheries. In The Ecology of Sandy Shores, 3rd ed.; McLachlan, A., Defeo, O., Eds.; Academic Press: Cambridge, UK, 2018; pp. 331–372. [Google Scholar]
- Rosa, A.O. A importância dos mariscos na subsistência de antigos grupos indígenas no litoral central Sítios RS-LC-81, 86, 87, 90, 92 e 96. Pesq. Antropol. 2006, 63, 259–288. [Google Scholar]
- Frontini, R.; Bayón, C. Archaeomalacological remains from the Puente de Fierro site (Buenos Aires province, Argentina). Arqueologia 2017, 23, 11–26. [Google Scholar]
- Defeo, O.; Gianelli, I.; Ortega, L.; Pittman, J. Responses of a small-scale shellfishery to climate change: Foundations for adaptive management. In Adaptive Management of Fisheries in Response to Climate Change. FAO Fisheries and Aquaculture Technical Paper No. 667; Bahri, T., Vasconcellos, M., Welch, D.J., Johnson, J., Perry, R.I., Ma, X., Sharma, R., Eds.; FAO: Rome, Italy, 2021; pp. 147–160. [Google Scholar]
- Olivier, S.R.; Capezzani, D.A.A.; Carreto, J.I.; Christiansen, H.E.; Aizpun de Moreno, J.E.; Penchaszadeh, P.E. Estructura de la Comunidad, Dinámica de la Población y Biología de la Almeja Amarilla (Mesodesma mactroides Desh. 1854) en Mar Azul (Pdo. de Gral Madariaga, Bs. As., Argentina); Instituto de Biología Marina: Mar del Plata, Argentina, 1971; 90p. [Google Scholar]
- Herrmann, M.; Alfaya, J.E.F.; Lepore, M.L.; Penchaszadeh, P.E.; Arntz, W. Population structure, growth and production of the yellow clam Mesodesma mactroides (Bivalvia: Mesodesmatidae) from a high-energy, temperate beach in northern Argentina. Helgol. Mar. Res. 2011, 65, 285–297. [Google Scholar] [CrossRef]
- Odebrecht, C.; Rörig, L.R.; Garcia, V.M.T.; Abreu, P.C. Shellfish mortality and a red tide event in southern Brazil. In Harmful Marine Algal Blooms; Lassus, P., Arzul, G., Erard, E., Gentien, P., Marcaillou, C., Eds.; Lavoisier Science: Paris, France, 1995; pp. 213–218. [Google Scholar]
- Fiori, S.M.; Cazzaniga, N.J. Mass mortality of the yellow clam, Mesodesma mactroides (Bivalvia: Mactracea) in Monte Hermoso beach, Argentina. Biol. Conserv. 1999, 89, 305–309. [Google Scholar] [CrossRef]
- Carvalho, Y.B.M.; Poersch, L.H.; Romano, L.A. Rickettsia-associated mortality of the yellow clam Mesodesma mactroides (Bivalvia: Mesodesmatidae) in southern Brazil. Malacologia 2013, 56, 301–307. [Google Scholar] [CrossRef]
- Cremonte, F.; Figueras, A. Parasites as possible cause of mass mortalities of the critically endangered clam Mesodesma mactroides on the Atlantic coast of Argentina. Bull. Eur. Assoc. Fish Pathol. 2004, 24, 166–171. [Google Scholar]
- Vasquez, N.; Fiori, S.; Arzul, I.; Carcedo, C.; Cremonte, F. Mass mortalities affecting populations of the yellow clam Amarilladesma mactroides along its geographic range. J. Shellfish Res. 2016, 35, 739–745. [Google Scholar] [CrossRef]
- Thompson, G.; Bock, M. Mortandad masiva de Mesodesma mactroides (Bivalvia: Mactracea) en el partido de la Costa, Buenos Aires, Argentina, en septiembre 2004. Atlântica 2007, 29, 115–119. [Google Scholar]
- Lopes, F.C.; Castro, M.R.; Barbosa, S.C.; Primel, E.G.; Martins, C.M.G. Effect of the UV filter, Benzophenone-3, on biomarkers of the yellow clam (Amarilladesma mactroides) under different pH conditions. Mar. Pollut. Bull. 2020, 158, 111401. [Google Scholar] [CrossRef] [PubMed]
- Truchet, D.M.; Forero Lopez, A.D.; Ardusso, M.G.; Rimondino, G.N.; Buzzi, N.S.; Malanca, F.E.; Spetter, C.V.; Fernández Severini, M.D. Microplastics in bivalves, water and sediments from a touristic sandy beach of Argentina. Mar. Pollut. Bull. 2021, 173, 113023. [Google Scholar] [CrossRef] [PubMed]
- Bom, F.C.; Colling, L.A. Impact of vehicles on benthic macrofauna on a subtropical sand beach. Mar. Ecol. 2020, 41, e12595. [Google Scholar] [CrossRef]
- ICMBio—Instituto Chico Mendes de Conservação da Biodiversidade. Livro Vermelho da Fauna Brasileira Ameaçada de Extinção—Invertebrados; ICMBio: Brasília, Brazil, 2018; 492p. [Google Scholar]
- Cledón, M.; Nuñez, J.D. Siphon nipping facilitates lethal predation in the clam Mesodesma mactroides (Reeve, 1854) (Mollusca: Bivalva). Mar. Biol. 2010, 157, 737–745. [Google Scholar] [CrossRef]
- Santos, J.J.S.; Bernardes, J.P.; Ramírez, J.R.B.; Gomes, C.H.A.M.; Romano, L.A. Embryo and larval development of the yellow clam Mesodesma mactroides (Reeve, 1854) (Mesodesmatidae) in laboratory. An. Acad. Bras. Cien. 2020, 92, e20190053. [Google Scholar] [CrossRef] [PubMed]
- Proverbio, C.; Carnevia, D.; Jorge-Romero, G.; Lercari, D. Tools for handling the yellow clam Mesodesma mactroides under experimental captivity conditions. INNOTEC 2019, 18, 124–141. [Google Scholar] [CrossRef]
- Santos, J.J.S.; Bernardes, J.P.; Ramírez, J.R.B.; Gomes, C.H.A.M.; Romano, L.A. Effect of salinity on embryo-larval development of yellow clam Mesodesma mactroides (Reeve, 1854) in laboratory. An Acad. Bras. Cien. 2020, 92, e20190169. [Google Scholar] [CrossRef]
- Santos, J.J.S.; Bernardes, J.P.; Ramírez, J.R.B.; Melo, C.M.R.; Romano, L.A.; Gomes, C.H.A.M. Effect of temperature on embryo-larval development of Amarilladesma mactroides (Reeve, 1854). Aquac. Res. 2022, 53, 3090–3096. [Google Scholar] [CrossRef]
- Gauthier, N.B.; Goes, F.S.; Quaresma, L.; Pedrosa, V.F.; Roselet, F.; Romano, L.A.; Cavalli, R.O. Design and optimization of an experimental maintenance system for yellow clam broodstock Amarilladesma mactroides (Reeve, 1854). Braz. J. Biol. 2022, 82, e243168. [Google Scholar] [CrossRef]
- Ayerbe, R.; Zevallos, S.; Castañeda, V.; Lope, F.; Bendita, H.; Sanz, Y. Manual: Cultivo de Macha Mesodesma Donacium (Lamarck, 1818) en la Región Moquegua; Instituto del Mar del Perú: El Callao, Peru, 2018; Volume 45, pp. 242–262. [Google Scholar]
- Gadomski, K.; Moller, H.; Beentjes, M.; Lamare, M. Embryonic and larval development of the New Zealand bivalve Paphies ventricosa Gray, 1843, (Veneroida: Mesodesmatidae) at a range of temperatures. J. Mollus. Stud. 2014, 81, 356–364. [Google Scholar] [CrossRef]
- Zevallos, S.; Toledo, P. Use of microalgae for broodstock conditioning of Mesodesma donacium (Mesodesmatidae). Biologist 2017, 15, 109–117. [Google Scholar]
- Costa, F.; Aranda-Burgos, J.A.; Cerviño-Otero, A.; Fernández-Pardo, A.; Louzán, A.; Nóvoa, S.; Ojea, J.; Martínez-Patiño, D. Clam Reproduction. In Clam Fisheries and Aquaculture; González, F.C., Ed.; Nova Science Publishers: New York, NY, USA, 2013; Chapter 3, pp. 45–71. [Google Scholar]
- Taynaros, S.; Tarangkoon, W. Variability in larval period, post-setting growth and survival of the oyster Crassostrea belcheri produced by gamete stripping method. Agric. Nat. Res. 2016, 50, 295–298. [Google Scholar] [CrossRef]
- Helm, M.; Bourne, N.; Lovatelli, A. Hatchery Culture of Bivalves—A Practical Manual, 1st ed.; FAO: Rome, Italy, 2004; 203p. [Google Scholar]
- Froese, R. Cube law, condition factor and weight–length relationships: History, meta-analysis and recommendations. J. Appl. Ichthyol. 2006, 22, 241–253. [Google Scholar] [CrossRef]
- UNESCO—United Nations Educational, Scientific and Cultural Organization. Chemical Methods for Use in Marine Environmental Monitoring; Intergovernmental Oceanographic Commission: Paris, France, 1983; 53p. [Google Scholar]
- AOAC—Association of Official AnalyticaL Chemist. Official Methods of Analysis, 15th ed.; AOAC: Washington, DC, USA, 1990; 90p. [Google Scholar]
- APHA—American Public Health Association. Standard Methods for the Examination of Water and Wastewater, 20th ed.; APHA: Washington, DC, USA, 1998; 1258p. [Google Scholar]
- CBRA—Colégio Brasileiro de Reprodução Animal. Manual Para Exames Andrológicos e Avaliação de Sêmen Animal, 2nd ed.; CBRA: Belo Horizonte, Brazil, 1998; 49p. [Google Scholar]
- Masello, A. Consideraciones Sobre Crecimiento y Biologia Reproductiva de la Almeja Amarilla Mesodesma mactroides (Deshayes, 1854). Bachelor’s Thesis, Universidad de la Republica, Montevideo, Uruguay, 1987. [Google Scholar]
- Lango-Reynoso, F.; Chavez-Villalba, J.; Cochard, J.C.; Le Pennec, M. Oocyte size, a means to evaluate the gametogenic development of the Pacific oyster, Crassostrea gigas (Thunberg). Aquaculture 2000, 190, 183–199. [Google Scholar] [CrossRef]
- Utting, S.D. Techniques for the hatchery conditioning of bivalve broodstocks and the subsequent effect on egg quality and larval viability. Aquaculture 1997, 155, 45–54. [Google Scholar] [CrossRef]
- Gosling, E. Marine Bivalve Molluscs, 2nd ed.; John Wiley & Sons: New York, NY, USA, 2015; pp. 157–202. [Google Scholar]
- Rico-Villa, B.; Le Coz, J.R.; Mingant, C.; Robert, R. Influence of phytoplankton diet mixtures on microalgae consumption, larval development and settlement of the Pacific oyster Crassostrea gigas (Thunberg). Aquaculture 2006, 256, 377–388. [Google Scholar] [CrossRef]
- Cheng, P.; Zhou, C.; Chu, R.; Chang, T.; Xu, J.; Ruan, R.; Chen, P.; Yan, X. Effect of microalgae diet and culture system on the rearing of bivalve mollusks: Nutritional properties and potential cost improvements. Algal Res. 2020, 51, 102076. [Google Scholar] [CrossRef]
- Argüello-Guevara, W.; Loor, A.; Sonnenholzner, S. Broodstock conditioning, spawning induction, and early larval development of the tropical rock oyster Striostrea prismatica (Gray 1825). J. Shellfish Res. 2013, 32, 665–670. [Google Scholar] [CrossRef]
- Nowland, S.J.; O’Connor, W.A.; Elizur, A.; Southgate, P.C. Evaluating spawning induction methods for the tropical black-lip rock oyster, Saccostrea echinata. Aquac. Rep. 2021, 20, 100676. [Google Scholar] [CrossRef]
- Heslinga, G.; Watson, T.; Isamu, T. Giant Clam Farming; Pacific Fisheries Development Foundation (NMFS/NOAA): Honolulu, HI, USA, 1990; 179p.
- Trigos, S.; Vicente, N.; Prado, P.; Espinós, F.J. Adult spawning and early larval development of the endangered bivalve Pinna nobilis. Aquaculture 2018, 483, 102–110. [Google Scholar] [CrossRef]
- Carvalho, Y.B.M.; Romano, L.A.; Poersch, L.H.S. Effect of low salinity on the yellow clam Mesodesma mactroides. Braz. J. Biol. 2015, 75, 8–12. [Google Scholar] [CrossRef] [PubMed]
- Medeiros, I.; Faria, S.C.; Souza, M.M. Osmoionic homeostasis in bivalve mollusks from different osmotic niches: Physiological patterns and evolutionary perspectives. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2020, 240, 110582. [Google Scholar] [CrossRef] [PubMed]
- Morse, D.E.; Duncan, H.; Hooker, N.; Morse, A. Hydrogen peroxide induces spawning in mollusks, with activation of prostaglandin endoperoxide synthetase. Science 1977, 196, 298–300. [Google Scholar] [CrossRef]
- Ferranti, M.P.; Monteggia, D.; Asnaghi, V.; Chiantore, M. Artificial reproduction protocol, from spawning to metamorphosis, through noninvasive methods in Patella caerulea Linnaeus, 1758. Aquac. Res. 2018, 49, 3386–3391. [Google Scholar] [CrossRef]
- García, L.N.; Murillo, D.L.; Tabares, P.A.; Paredes, F.J.; Chapman, F.A. Induced spawning of the blood ark Anadara tuberculosa, using hydrogen peroxide. AACL Bioflux 2018, 11, 1248–1251. [Google Scholar]
- Flint, M.; Than, J.T. Potential spawn induction and suppression agents in Caribbean Acropora cervicornis corals of the Florida Keys. PeerJ 2016, 4, e1982. [Google Scholar] [CrossRef]
- Haque, M.N.; Kwon, S.H. Physiological effects of biocide on marine bivalve blue mussels in context prevent macrofouling. J. Ecol. Environ. 2016, 40, 1–8. [Google Scholar] [CrossRef][Green Version]



{kind=link}
{kind=link}
{kind=link}
| Trials with Conditioning | Trials with no Conditioning | |||||
|---|---|---|---|---|---|---|
| I | II | III | IV | V | VI | |
| Date | 15 February 2019 | 14 March 2019 | 19 September 2019 | 07 November 2019 | 18 November 2019 | 10 December 2019 |
| Site | Cassino | Mar Grosso | Mar Grosso | Mar Grosso | Mar Grosso | Cassino |
| Temperature (°C) | 23.0 | 24.0 | 18.0 | 21.5 | 22.0 | 21.8 |
| Salinity | 32 | 28 | 24 | 25 | 28 | 31 |
| WWi (g) | 12.44 ± 1.12 | 19.77 ± 1.14 b | 13.74 ± 1.19 b | nd | nd | nd |
| WWf (g) | 12.56 ± 1.44 | 21.31 ± 3.02 a | 16.37 ± 2.71 a | 18.27 ± 2.43 | 16.65 ± 3.64 | 17.74 ± 1.84 |
| DWi (g) | nd | 8.87 ± 0.92 | 5.24 ± 0.47 | nd | nd | nd |
| DWf (g) | 5.03 ± 0.15 | 7.81 ± 0.13 | 6.05 ± 0.17 | 6.75 ± 1.12 | 6.37 ± 1.52 | 7.35 ± 0.70 |
| Li (mm) | 53.10 ± 1.93 | 62.30 ± 2.51 | 56.87 ± 2.68 | nd | nd | nd |
| Lf (mm) | 53.23 ± 1.73 | 62.45 ± 2.71 | 57.01 ± 2.79 | 59.78 ± 3.14 | 58.06 ± 3.49 | 59.00 ± 2.34 |
| Hi (mm) | 28.57 ± 0.02b | 33.25 ± 0.02 | 30.27 ± 0.03 b | nd | nd | nd |
| Hf (mm) | 28.84 ± 1.03a | 33.35 ± 1.04 | 30.61 ± 1.00 a | 32.58 ± 1.74 | 31.24 ± 1.71 | 31.95 ± 1.50 |
| Cfi | 8.34 ± 0.66 | 8.36 ± 0.63 b | 7.97 ± 0.76 b | nd | nd | nd |
| CFf | 8.31 ± 0.58 | 8.73 ± 0.87 a | 8.86 ± 0.86 a | 8.54 ± 0.97 | 8.41 ± 0.92 | 8.65 ± 0.75 |
| Survival during conditioning (%) | 88.3 | 85.0 | 78.33 | nd | nd | nd |
| Survival post-induction (%) | 88.3 | 80.0 | 76.67 | 82.5–87.5 | 83.3–100 | 55.55–100 |
| Trial | Di | Df | di | df |
|---|---|---|---|---|
| II | 27.62 ± 7.37 b | 32.26 ± 2.71 a | 20.31 ± 7.43 b | 27.04 ± 4.32 a |
| III | 40.82 ± 6.51 a | 44.17 ± 5.39 a | 31.71 ± 4.56 b | 36.37 ± 3.84 a |
| IV | --- | 42.54 ± 4.27 | --- | 30.37 ± 4.40 |
| V | --- | 36.65 ± 4.95 | --- | 27.27 ± 3.21 |
| VI | --- | 32.29 ± 4.68 | --- | 26.93 ± 2.95 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gauthier, N.B.; Góes, F.S.; Pedrosa, V.F.; Roselet, F.; Romano, L.A.; Cavalli, R.O. Towards the Control of the Reproduction of the Yellow Clam Amarilladesma mactroides (Reeve, 1854) in Captivity: Effects of Different Stimuli on the Spawning of Laboratory-Conditioned and Unconditioned Breeders. Fishes 2023, 8, 37. https://doi.org/10.3390/fishes8010037
Gauthier NB, Góes FS, Pedrosa VF, Roselet F, Romano LA, Cavalli RO. Towards the Control of the Reproduction of the Yellow Clam Amarilladesma mactroides (Reeve, 1854) in Captivity: Effects of Different Stimuli on the Spawning of Laboratory-Conditioned and Unconditioned Breeders. Fishes. 2023; 8(1):37. https://doi.org/10.3390/fishes8010037
Chicago/Turabian StyleGauthier, Nathália Byrro, Filipe Scavone Góes, Virgínia Fonseca Pedrosa, Fábio Roselet, Luis Alberto Romano, and Ronaldo Olivera Cavalli. 2023. "Towards the Control of the Reproduction of the Yellow Clam Amarilladesma mactroides (Reeve, 1854) in Captivity: Effects of Different Stimuli on the Spawning of Laboratory-Conditioned and Unconditioned Breeders" Fishes 8, no. 1: 37. https://doi.org/10.3390/fishes8010037
APA StyleGauthier, N. B., Góes, F. S., Pedrosa, V. F., Roselet, F., Romano, L. A., & Cavalli, R. O. (2023). Towards the Control of the Reproduction of the Yellow Clam Amarilladesma mactroides (Reeve, 1854) in Captivity: Effects of Different Stimuli on the Spawning of Laboratory-Conditioned and Unconditioned Breeders. Fishes, 8(1), 37. https://doi.org/10.3390/fishes8010037