
Development of an Immunoassay Detection System for Koi Herpesvirus Using Recombinant Single-Chain Variable Fragments


Abstract
1. Introduction
2. Methods
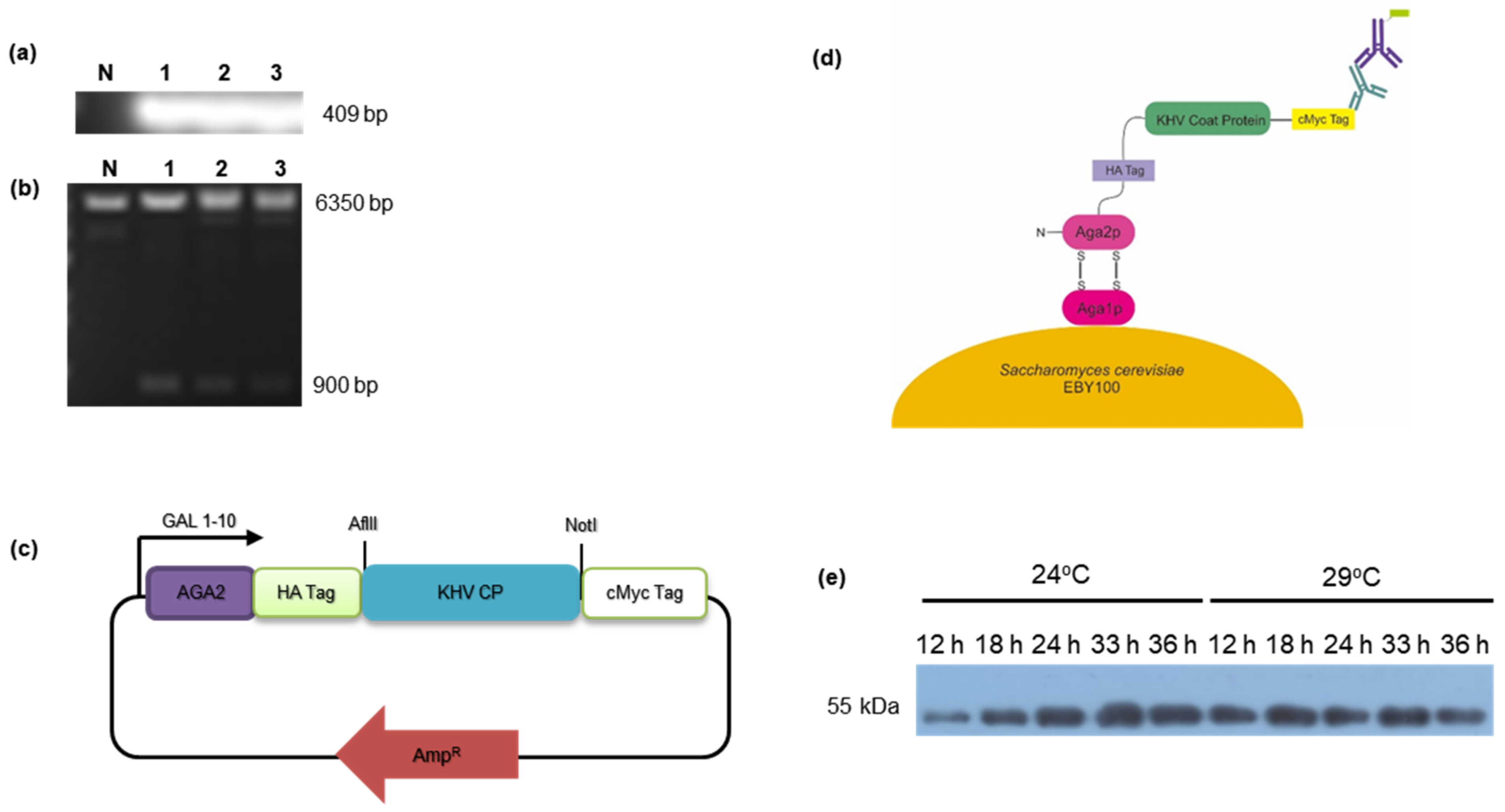
2.1. Subcloning of the pCTCON-KHV Coat Protein
2.2. Yeast Transformation and Yeast Surface Display Expression
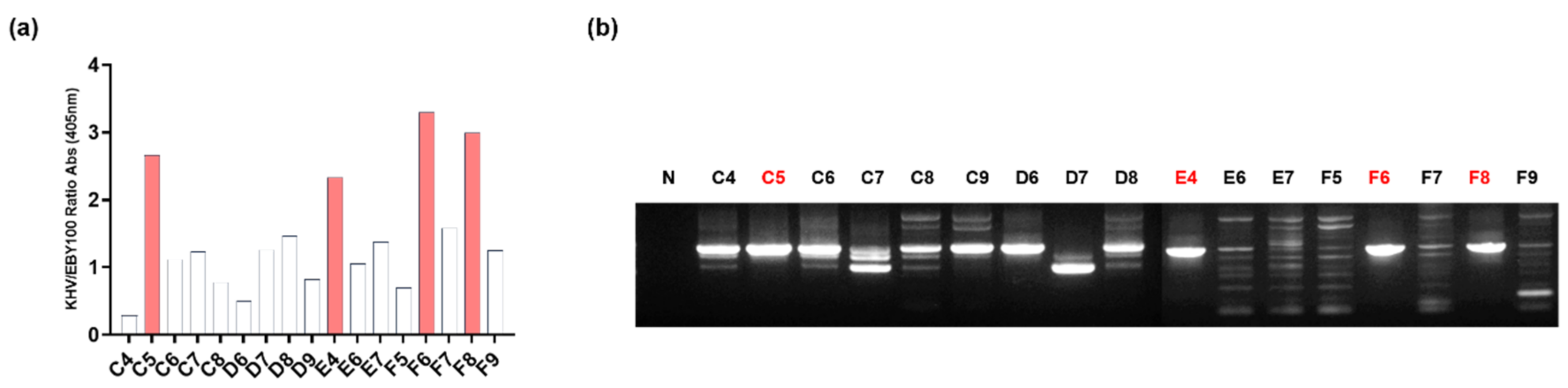
2.3. Biopanning
2.4. Phage ELISA
2.5. Sequence Analysis and Mutagenesis of Selected KHV-Specific Phage scFvs
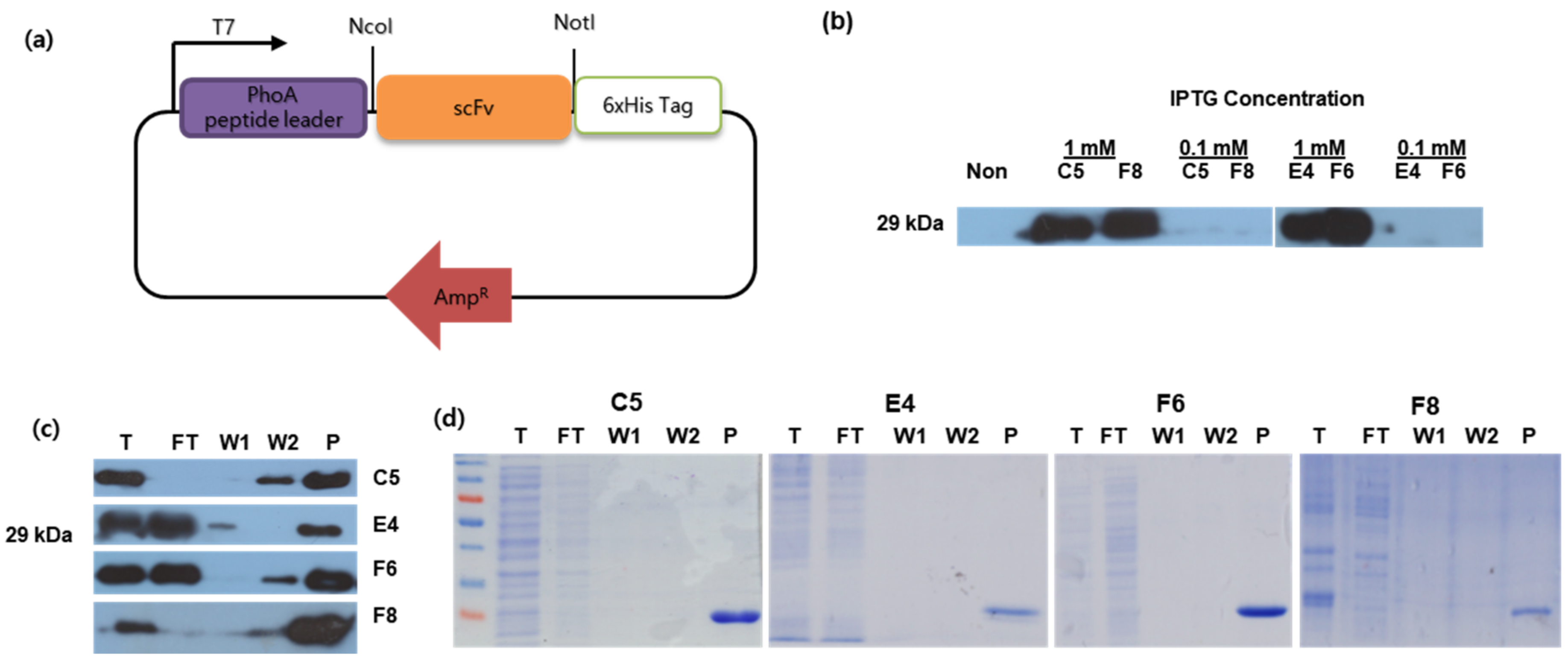
2.6. Putative scFv Protein Expression and Purification
2.7. KHV Propagation, Purification, and Viral Titer
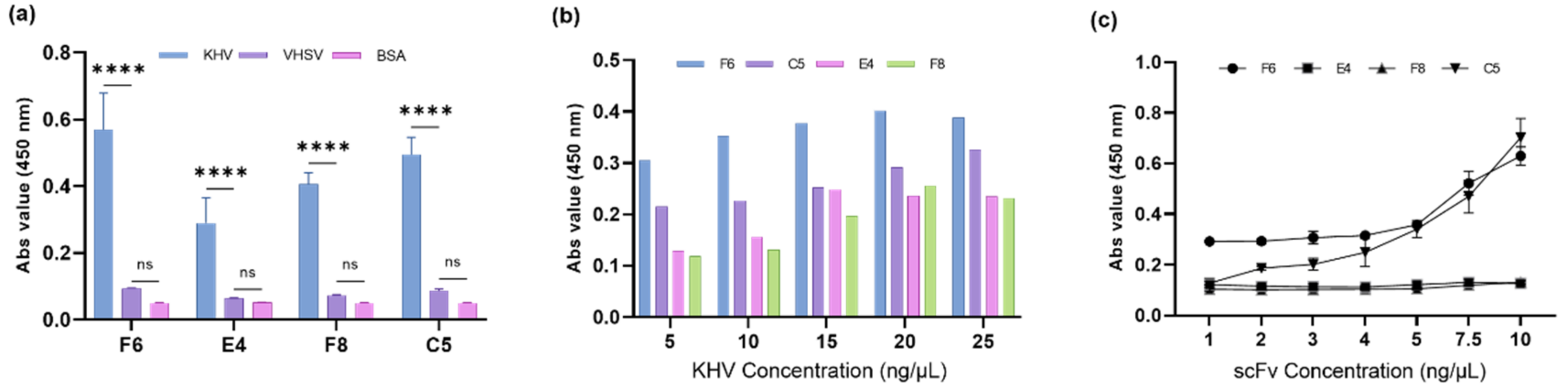
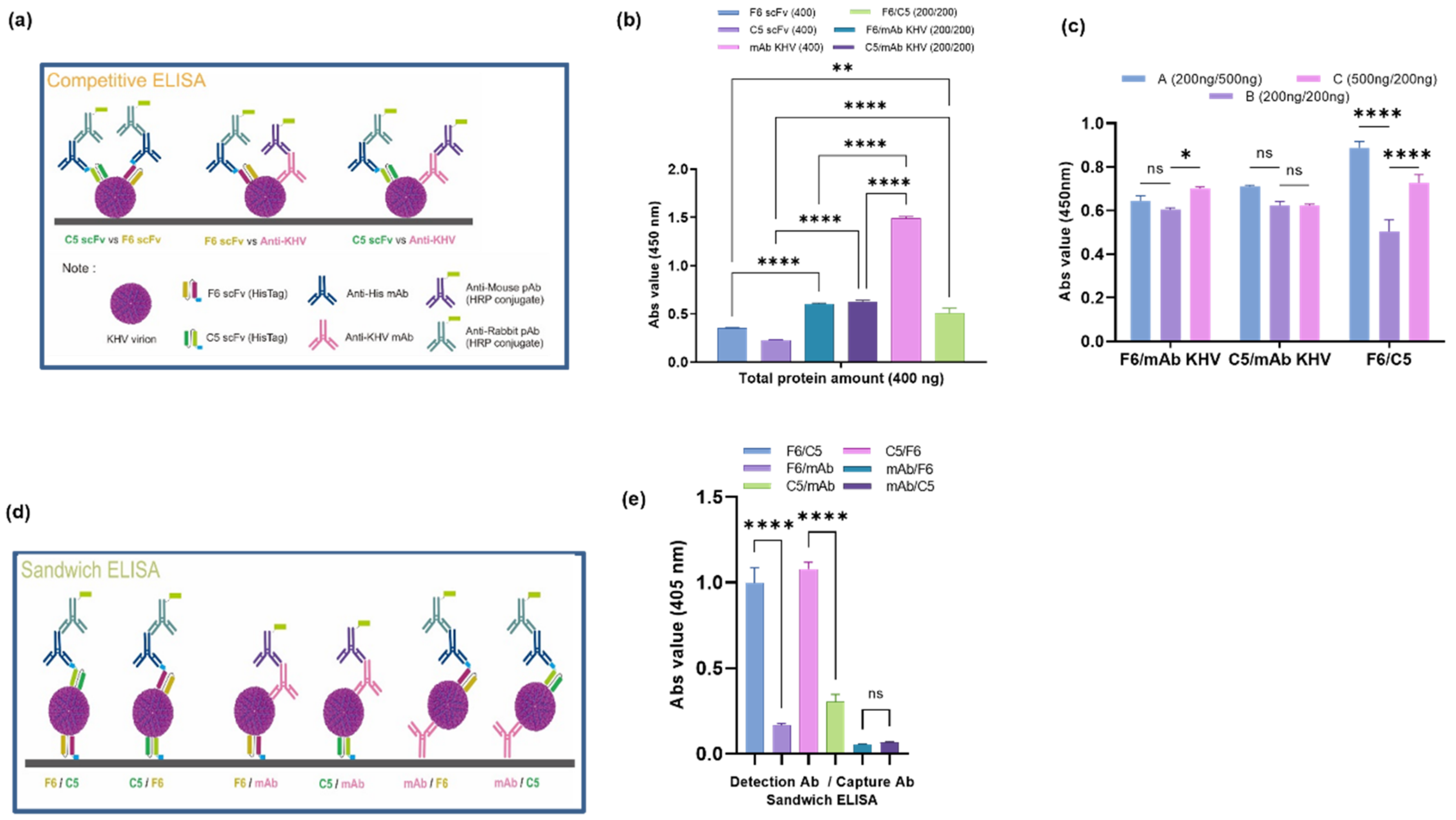
2.8. Analysis of the Binding Specificity of Putative scFv Proteins
2.9. Sensitivity Analysis of Putative scFv Proteins
2.10. Epitope Mapping of Putative scFv Proteins
3. Results
3.1. Subcloning of KHV Coat Protein and Antigen Expression by Yeast Surface Display
3.2. Screening of Anti-KHV-Specific scFv by Biopanning
3.3. ScFv Sequence Analysis and CDR Determination
3.4. Protein Expression and Characterization of Anti-KHV scFv
3.5. Epitope Mapping of Putative scFv Proteins
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- FAO. The State of World Fisheries and Aquaculture 2018; FAO: Rome, Italy, 2018. [Google Scholar]
- Pokorova, D.; Vesely, T.; Piackova, V.; Reschova, S.; Hulova, J. Current knowledge on koi herpesvirus (KHV): A review. Vet. Med. 2005, 50, 139–148. [Google Scholar] [CrossRef]
- Waltzek, T.; Kelley, G.O.; Stone, D.M.; Way, K.; Hanson, L.; Fukuda, H.; Hirono, I.; Aoki, T.; Davison, A.J.; Hedrick, R.P. Koi herpesvirus represents a third cyprinid herpesvirus (CyHV-3) in the family Herpesviridae. J. Gen. Virol. 2005, 86, 1659–1667. [Google Scholar] [CrossRef] [PubMed]
- Haenen, O.; Way, K.; Bergmann, S.; Ariel, E. The emergence of koi herpesvirus and its significance to European aquaculture. Bull. Eur. Assoc. Fish Pathol. 2004, 24, 293–307. [Google Scholar]
- Bretzinger, A.; Fischer-Scherl, T.; Oumouna, M.; Hoffmann, R.; Truyen, U. Mass mortalities in Koi carp, Cyprinus carpio, associated with gill and skin disease. Bull. Eur. Assoc. Fish Pathol. 1999, 19, 182–185. [Google Scholar]
- Sunarto, A.; McColl, K.A.; Crane, M.; Sumiati, T.; Hyatt, A.D.; Barnes, A.; Walker, P.J. Isolation and characterization of koi herpesvirus (KHV) from Indonesia: Identification of a new genetic lineage. J. Fish Dis. 2010, 34, 87–101. [Google Scholar] [CrossRef]
- Tu, C.; Weng, M.-C.; Shiau, J.-R.; Lin, S.-Y. Detection of Koi Herpesvirus in Koi Cyprinus carpio in Taiwan. Fish Pathol. 2004, 39, 109–110. [Google Scholar] [CrossRef]
- Gomez, D.K.; Joh, S.J.; Jang, H.; Shin, S.P.; Choresca, C.H.; Han, J.E.; Kim, J.H.; Jun, J.W.; Park, S.C. Detection of koi herpesvirus (KHV) from koi (Cyprinus carpio koi) broodstock in South Korea. Aquaculture 2011, 311, 42–47. [Google Scholar] [CrossRef]
- Cohen, A. Non-Native Bacterial and Viral Pathogens in Ballast Water: Potential for Impacts to ESA-Listed Species under NOAA’s Jurisdiction; Endangered Species Division, Silver Spring; MD Center for Research on Aquatic Bioinvasions (CRAB): Richmond, CA, USA, 2010. [Google Scholar]
- Uchii, K.; Matsui, K.; Iida, T.; Kawabata, Z. Distribution of the introduced cyprinid herpesvirus 3 in a wild population of common carp, Cyprinus carpio L. J. Fish Dis. 2009, 32, 857–864. [Google Scholar] [CrossRef]
- Kielpinski, M.; Kempter, J.; Panicz, R.; Sadowski, J.; Schütze, H.; Ohlemeyer, S.; Bergmann, S.M. Detection of KHV in Freshwater Mussels and Crustaceans from Ponds with KHV History in Common Carp (Cyprinus carpio). Isr. J. Aquac.-Bamidgeh 2010, 62, 20576. [Google Scholar] [CrossRef]
- Boutier, M.; Ronsmans, M.; Rakus, K.; Jazowiecka-Rakus, J.; Vancsok, C.; Morvan, L.; Peñaranda, M.M.D.; Stone, D.M.; Way, K.; van Beurden, S.J.; et al. Cyprinid herpesvirus 3: An archetype of fish alloherpesviruses. Adv. Virus Res. 2015, 93, 161–256. [Google Scholar]
- Uchii, K.; Minamoto, T.; Honjo, M.N.; Kawabata, Z. Seasonal reactivation enables Cyprinid herpesvirus 3 to persist in a wild host population. FEMS Microbiol. Ecol. 2014, 87, 536–542. [Google Scholar] [CrossRef]
- Gilad, O.; Yun, S.; Zagmutt-Vergara, F.; Leutenegger, C.; Bercovier, H.; Hedrick, R. Concentrations of a Koi herpesvirus (KHV) in tissues of experimentally-infected Cyprinus carpio koi as assessed by real-time TaqMan PCR. Dis. Aquat. Org. 2004, 60, 179–187. [Google Scholar] [CrossRef]
- Bercovier, H.; Fishman, Y.; Nahary, R.; Sinai, S.; Zlotkin, A.; Eyngor, M.; Gilad, O.; Eldar, A.; Hedrick, R.P. Cloning of the koi herpesvirus (KHV) gene encoding thymidine kinase and its use for a highly sensitive PCR based diagnosis. BMC Microbiol. 2005, 5, 13. [Google Scholar] [CrossRef]
- Yoshino, M.; Watari, H.; Kojima, T.; Ikedo, M.; Kurita, J. Rapid, sensitive and simple detection method for koi herpesvirus using loop-mediated isothermal amplification. Microbiol. Immunol. 2009, 53, 375–383. [Google Scholar] [CrossRef]
- Adkison, M.A.; Gilad, O.; Hedrick, R.P. An Enzyme Linked Immunosorbent Assay (ELISA) for Detection of Antibodies to the Koi Herpesvirus (KHV) in the Serum of Koi Cyprinus carpio. Fish Pathol. 2005, 40, 53–62. [Google Scholar] [CrossRef]
- Bergmann, S.M.; Wang, Q.; Zeng, W.; Li, Y.; Wang, Y.; Matras, M.; Reichert, M.; Fichtner, D.; Lenk, M.; Morin, T.; et al. Validation of a KHV antibody enzyme-linked immunosorbent assay (ELISA). J. Fish Dis. 2017, 40, 1511–1527. [Google Scholar] [CrossRef]
- Bergmann, S.M.; Riechardt, M.; Fichtner, D.; Lee, P.; Kempter, J. Investigation on the diagnostic sensitivity of molecular tools used for detection of koi herpesvirus. J. Virol. Methods 2010, 163, 229–233. [Google Scholar] [CrossRef]
- Monaghan, S.; Thompson, K.; Adams, A.; Bergmann, S.M. Sensitivity of seven PCR s for early detection of koi herpesvirus in experimentally infected carp, Cyprinus carpio L., by lethal and non-lethal sampling methods. J. Fish Dis. 2015, 38, 303–319. [Google Scholar] [CrossRef]
- Salazar, G.; Zhang, N.; Fu, T.-M.; An, Z. Antibody therapies for the prevention and treatment of viral infections. npj Vaccines 2017, 2, 19. [Google Scholar] [CrossRef]
- Trier, N.H.; Houen, G. Antibodies as Diagnostic Targets and as Reagents for Diagnostics; Multidisciplinary Digital Publishing Institute: Basel, Switzerland, 2020. [Google Scholar]
- Nelson, A.L.; Reichert, J.M. Development trends for therapeutic antibody fragments. Nat. Biotechnol. 2009, 27, 331–337. [Google Scholar] [CrossRef]
- Ahmad, Z.A.; Yeap, S.K.; Ali, A.M.; Ho, W.Y.; Alitheen, N.B.M.; Hamid, M. scFv Antibody: Principles and Clinical Application. J. Immunol. Res. 2012, 2012, 980250. [Google Scholar] [CrossRef] [PubMed]
- Angelini, A.; Chen, T.F.; de Picciotto, S.; Yang, N.J.; Tzeng, A.; Santos, M.S.; Van Deventer, J.A.; Traxlmayr, M.W.; Wittrup, K.D. Protein engineering and selection using yeast surface display. In Yeast Surface Display; Springer: Berlin/Heidelberg, Germany, 2015; pp. 3–36. [Google Scholar]
- Martin, A.C. Accessing the Kabat Antibody Sequence Database by Computer; Wiley Online Library: New York, NY, USA, 1996. [Google Scholar]
- Dondelinger, M.; Filée, P.; Sauvage, E.; Quinting, B.; Muyldermans, S.; Galleni, M.; Vandevenne, M.S. Understanding the Significance and Implications of Antibody Numbering and Antigen-Binding Surface/Residue Definition. Front. Immunol. 2018, 9, 2278. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Chen, Y.; Ke, Y.; Zhang, L.; Zhang, B.; Yang, L.; Zhu, J. Single Chain Fragment Variable (scFv) Antibodies Targeting the Spike Protein of Porcine Epidemic Diarrhea Virus Provide Protection against Viral Infection in Piglets. Viruses 2019, 11, 58. [Google Scholar] [CrossRef] [PubMed]
- Mettenleiter, T.C. Intriguing interplay between viral proteins during herpesvirus assembly or: The herpesvirus assembly puzzle. Veter-Microbiol. 2006, 113, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Davison, A.J.; Kurobe, T.; Gatherer, D.; Cunningham, C.; Korf, I.; Fukuda, H.; Hedrick, R.P.; Waltzek, T.B. Comparative Genomics of Carp Herpesviruses. J. Virol. 2013, 87, 2908–2922. [Google Scholar] [CrossRef]
- Tu, C.; Lu, Y.-P.; Hsieh, C.-Y.; Huang, S.-M.; Chang, S.-K.; Chen, M.-M. Production of monoclonal antibody against ORF72 of koi herpesvirus isolated in Taiwan. Folia Microbiol. 2013, 59, 159–165. [Google Scholar] [CrossRef]
- Hong, F.; Meinander, N.Q.; Jönsson, L.J. Fermentation strategies for improved heterologous expression of laccase inPichia pastoris. Biotechnol. Bioeng. 2002, 79, 438–449. [Google Scholar] [CrossRef]
- Camarero, S.; Pardo, I.; Cañas, A.I.; Molina-Espeja, P.; Record, E.; Martínez, A.T.; Martínez, M.J.; Alcalde, M. Engineering Platforms for Directed Evolution of Laccase from Pycnoporus cinnabarinus. Appl. Environ. Microbiol. 2012, 78, 1370–1384. [Google Scholar] [CrossRef]
- Chen, X.; Wang, Y.; Li, Y. Protein L chromatography: A useful tool for monitoring/separating homodimers during the purification of IgG-like asymmetric bispecific antibodies. Protein Expr. Purif. 2020, 175, 105711. [Google Scholar] [CrossRef]
- Sakamoto, S.; Putalun, W.; Vimolmangkang, S.; Phoolcharoen, W.; Shoyama, Y.; Tanaka, H.; Morimoto, S. Enzyme-linked immunosorbent assay for the quantitative/qualitative analysis of plant secondary metabolites. J. Nat. Med. 2017, 72, 32–42. [Google Scholar] [CrossRef]
- Caprari, S.; Metzler, S.; Lengauer, T.; Kalinina, O.V. Sequence and Structure Analysis of Distantly-Related Viruses Reveals Extensive Gene Transfer between Viruses and Hosts and among Viruses. Viruses 2015, 7, 5388–5409. [Google Scholar] [CrossRef]
- Kirkitadze, M.D.; Barlow, P.N.; Price, N.C.; Kelly, S.M.; Boutell, C.J.; Rixon, F.J.; McClelland, D.A. The herpes simplex virus triplex protein, VP23, exists as a molten globule. J. Virol. 1998, 72, 10066–10072. [Google Scholar] [CrossRef]
- Kuwajima, K. The Molten Globule, and Two-State vs. Non-Two-State Folding of Globular Proteins. Biomolecules 2020, 10, 407. [Google Scholar] [CrossRef]
- Heming, J.D.; Conway, J.F.; Homa, F.L. Herpesvirus capsid assembly and DNA packaging. Cell Biol. Herpes Viruses 2017, 223, 119–142. [Google Scholar]
- Packer, M.S.; Liu, D.R. Methods for the directed evolution of proteins. Nat. Rev. Genet. 2015, 16, 379–394. [Google Scholar] [CrossRef]
- Rees, D.C.; Robertson, A.D. Some thermodynamic implications for the thermostability of proteins. Protein Sci. 2001, 10, 1187–1194. [Google Scholar] [CrossRef]
- Rodrigo, G.; Gruvegård, M.; Van Alstine, J.M. Antibody fragments and their purification by protein L affinity chromatography. Antibodies 2015, 4, 259–277. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | 5′-Sequence-3′ | ||||||
|---|---|---|---|---|---|---|---|
| Oligo_MCS_Fwd | NheI | AflII | NdeI | SmaI | SalI | SpeI | NotI |
| 5′-CTAGC CTTAAG CATATG CCCGGG GTCGAC ACTAGT A GCGGCCGC G-3′ | |||||||
| Oligo_MCS_Rev | 5′-GATCC GCGGCCGC T ACTAGT GTCGAC CCCGGG CATATG CTTAAG G-3′ | ||||||
| KHV_Part_Fwd | 5′-AAAGTACCAGATCGTAAACACGG-3′ | ||||||
| KHV_Part_Rev | 5′-ATCATGGAGAAGATCACGTCCA-3′ | ||||||
| Primer | 5′-Sequence-3′ | |||||
|---|---|---|---|---|---|---|
| Oligo_Ig20_MCS_Fwd | NcoI | AflII | NotI | XhoI | (6×)HisTag | Stop |
| 5′-GGG CCATGG CTTAAG GCGGCCGC C CTCGAG CATCATCATCATCATCAT TAA-3′ | ||||||
| Oligo_Ig20_MCS_Rev | 5′-TCGA TTA ATGATGATGATGATGATG CTCGAG G GCGGCCGC CTTAAG CCATGG CCC-3′ | |||||
| Round | Antigen [OD600] | Phage Added (cfu/mL) | Phage Eluted (cfu/mL) | Yield | Enrichment Fold |
|---|---|---|---|---|---|
| 1 | 10 | 1 × 109 | 2 × 108 | 2 × 10−1 | - |
| 2 | 7.5 | 5 × 107 | 2 × 107 | 4 × 10−1 | 2 |
| 3 | 5 | 5 × 108 | 2 × 107 | 4 × 10−2 | 20 |
| 4 | 2.5 | 3 × 109 | 1 × 107 | 3 × 10−3 | 150 |
| scFv | Variable Heavy Region | ||||||
| FR1 | CDR1 | FR2 | CDR2 | FR3 | CDR3 | FR4 | |
| C5 | EVQLLESGGGLVQPGGSLRLSCAAS | GFTFSSYA | MSWVRQAPGKGLEWVSS | SAAPGNAT | FYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYC | AKARVAFDY | WGQGTLVTVS |
| F8 | EVPLLESGGGLVQPGGSLRLSCAAS | GFTFSSYA | MSWVRQAPGKGLEWVSS | ILKRGVDT | CYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYC | AKLAETFDY | WGQGTLVTVS |
| F6 | EVQLLESGGGLVQPGGSLRLSCAAS | GFTFSSYA | MSWVRQAPGKGLEWVSK | IAANGVQT | DYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYC | AKLARLFDY | WGQGTLVTVS |
| E4 | EVQLLESGGGLVQPGGSLRLSCAAS | GFTFSSYA | MSWVRQAPGKGLEWVSS | IPKLGSKT | CFADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYC | AKTLNLFDY | WGQGTLVTVS |
| scFv | Variable Kappa Region | ||||||
| FR1 | CDR1 | FR2 | CDR2 | FR3 | CDR3 | FR4 | |
| C5 | DIQMTQSPSSLSASVGDRVTITCRAS | QSISSY | LNWYQQKPGKAPKLLIY | TAS | YLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYC | QQSEWLPMT | FGQGTKVEIK |
| F8 | DIQMTQSPSSLSASVGDRVTITCRAS | QSISSY | LNWYQQKPGKAPKLLIY | PAS | HLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYC | QQTRVHPPT | FGQGTKVEIK |
| F6 | DIQMTQSPSSLSASVGDRVTITCRAS | QSISSY | LNWYQQKPGKAPKLLIY | PAS | YLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYC | QQPDLPPGT | FGQGTKVEIK |
| E4 | DIQMTQSPSSLSASVGDRVTITCRAS | QSISSY | LNWYQQKPGKAPKLLIY | KAS | NLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYC | QQPPRRPRT | FGQGTKVEIK |
| scFv | Additivity Index | ||
|---|---|---|---|
| C5 | F6 | Anti-KHV | |
| C5 | - | 11.58 | 4.01 |
| F6 | 11.58 | - | 2.82 |
| KHV mAb | 4.01 | 2.82 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seo, H.; Lubis, A.D.M.; Choi, T.-J.; Jung, T.-S.; Lee, T.-K.; Lee, S. Development of an Immunoassay Detection System for Koi Herpesvirus Using Recombinant Single-Chain Variable Fragments. Fishes 2022, 7, 370. https://doi.org/10.3390/fishes7060370
Seo H, Lubis ADM, Choi T-J, Jung T-S, Lee T-K, Lee S. Development of an Immunoassay Detection System for Koi Herpesvirus Using Recombinant Single-Chain Variable Fragments. Fishes. 2022; 7(6):370. https://doi.org/10.3390/fishes7060370
Chicago/Turabian StyleSeo, Haneul, Andre Ditya Maulana Lubis, Tae-Jin Choi, Tae-Sung Jung, Taek-Kyun Lee, and Sukchan Lee. 2022. "Development of an Immunoassay Detection System for Koi Herpesvirus Using Recombinant Single-Chain Variable Fragments" Fishes 7, no. 6: 370. https://doi.org/10.3390/fishes7060370
APA StyleSeo, H., Lubis, A. D. M., Choi, T.-J., Jung, T.-S., Lee, T.-K., & Lee, S. (2022). Development of an Immunoassay Detection System for Koi Herpesvirus Using Recombinant Single-Chain Variable Fragments. Fishes, 7(6), 370. https://doi.org/10.3390/fishes7060370