
Ontogenetic Trophic Shifts by Ommastrephes bartramii in the North Pacific Ocean Based on Eye Lens Stable Isotopes


Abstract
1. Introduction
2. Materials and Methods
2.1. Squid Sampling
2.2. Statolith Processing and Increment Counting
2.3. Lens Delamination and Processing
2.4. Stable Isotope Analysis
2.5. Data Analysis
3. Results and Analysis
3.1. Ontogenetic Growth Distribution of the Squid
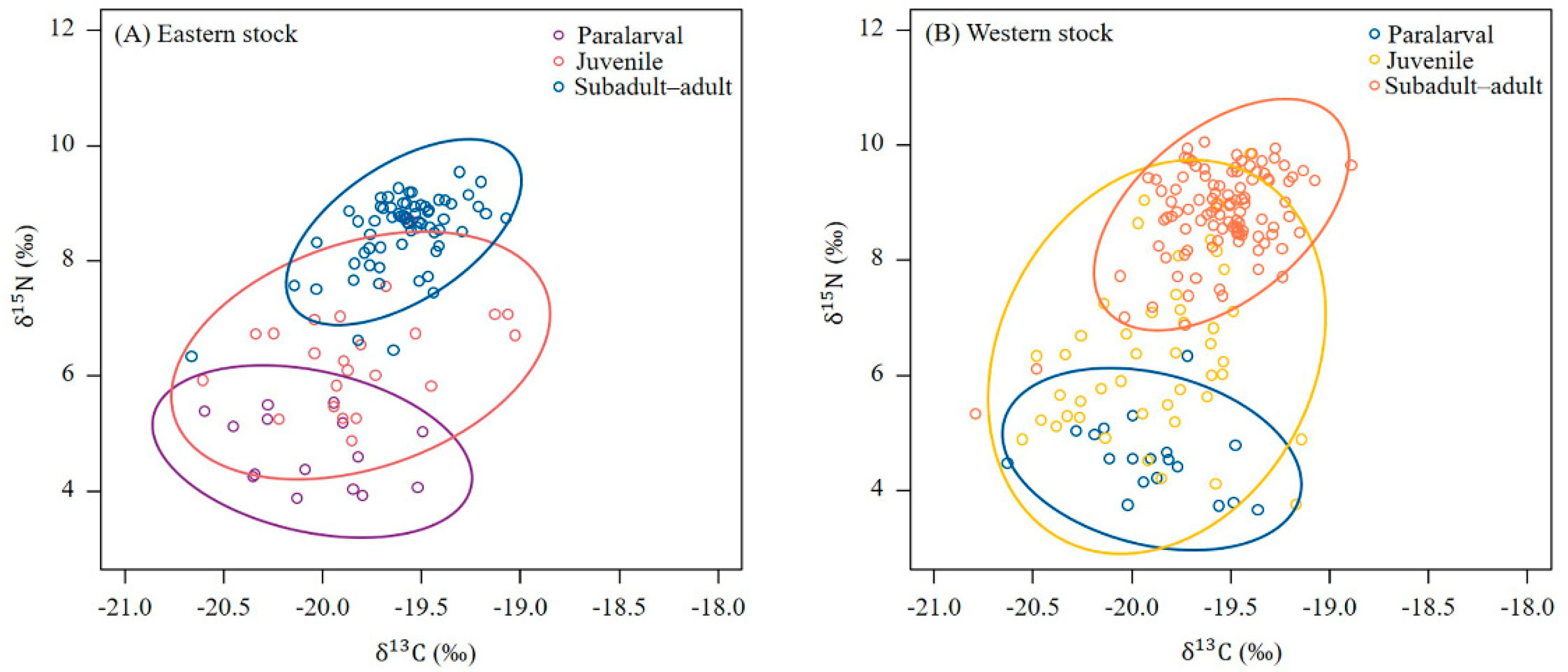
3.2. Distribution of Stable Isotope Values for the Eye Lenses
3.3. Temporal and Spatial Differences between the Stable Isotope Values of the Different Squid Stocks
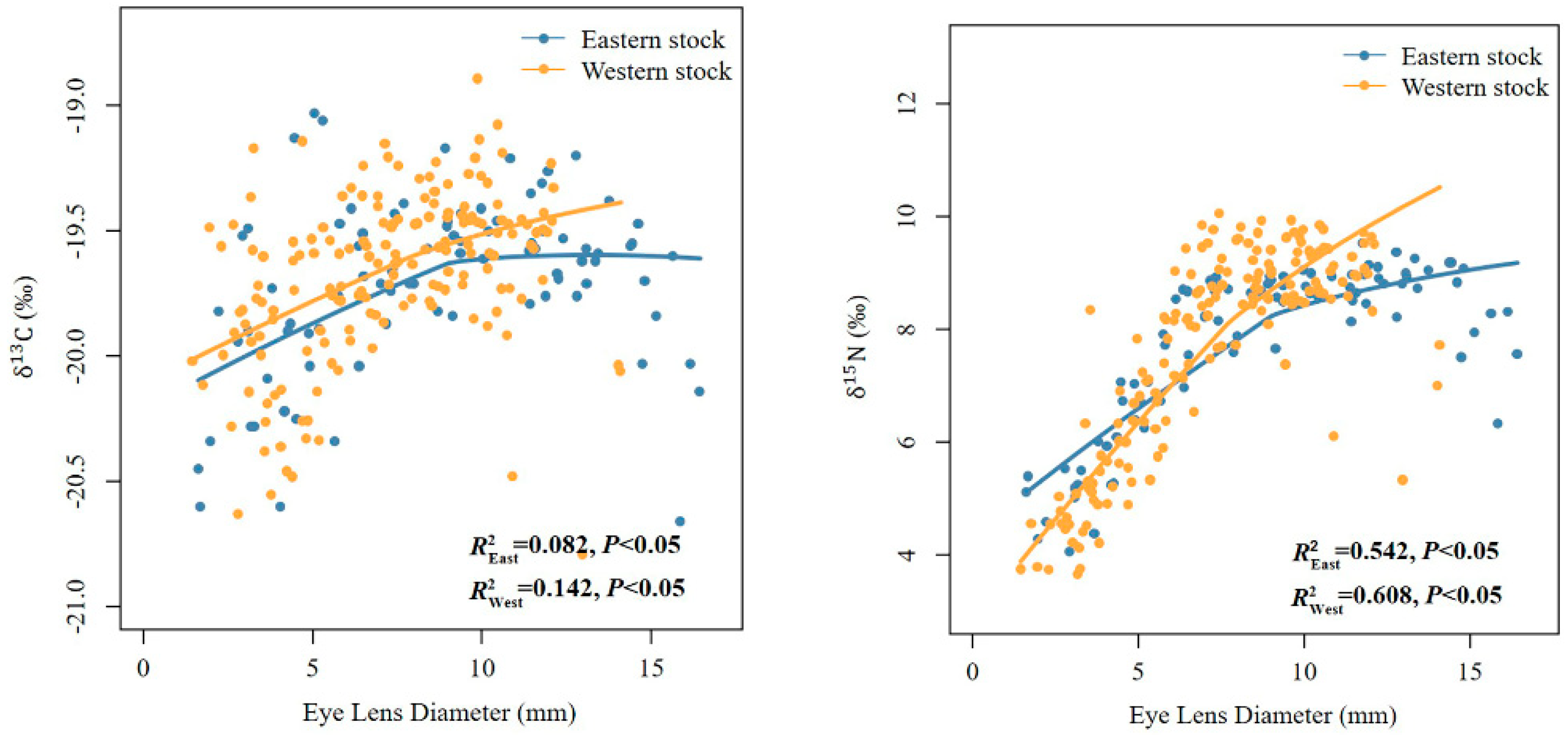
3.4. Assessment of the Suitability of Using Squid Eye Lenses to Determine the Ontogenetic Variation in Stable Isotope Values
4. Discussion
5. Conclusions and Prospects
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Roper, C.F.; Sweeney, M.J.; Nauen, C.E. Cephalopods of the World. An Annotated and Illustrated Catalogue of Species of Interest to Fisheries; United Nations Development Programme, Food and Agriculture Organization of the United Nations: Rome, Italy, 1984; Volume 3, pp. 1–277. [Google Scholar]
- Murata, M. On the flying behavior of neon flying squid Ommastrephes bartramii observed in the central and northwestern north pacific. Nippon Suisan Gakkaishi. 1988, 54, 1167–1174. [Google Scholar] [CrossRef]
- Murata, M. Oceanic resources of squids. Mar. Freshw. Behav. Phy. 1990, 18, 19–71. [Google Scholar] [CrossRef]
- Murata, M. Life history and biological information on flying squid (Ommastrephes bartramii) in the North Pacific Ocean. Bull. Int. Nat. North Pac. Comm. 1993, 53, 147–182. [Google Scholar]
- Xu, J.; Chen, X.; Chen, Y.; Ding, Q.; Tian, S. The effect of sea surface temperature increase on the potential habitat of Ommastrephes bartramii in the Northwest Pacific Ocean. Acta Ocean. Sin. 2016, 35, 109–116. [Google Scholar] [CrossRef]
- Vijai, D.; Sakai, M.; Wakabayashi, T.; Yoo, H.K.; Kato, Y.; Sakurai, Y. Effects of temperature on embryonic development and paralarval behavior of the neon flying squid Ommastrephes bartramii. Mar. Ecol. Prog. Ser. 2015, 529, 145–158. [Google Scholar] [CrossRef]
- Wang, J.; Yu, W.; Chen, X.; Lei, L.; Chen, Y. Detection of potential fishing zones for neon flying squid based on remote-sensing data in the Northwest Pacific Ocean using an artificial neural network. Int. J. Remote Sens. 2015, 36, 3317–3330. [Google Scholar] [CrossRef]
- Nigmatullin, C.M.; Shchetinnikov, A.S.; Shukhgalter, O.A. On feeding and helminth fauna of neon flying squid Ommastrephes bartramii (Lesueur, 1821) (Cephalopoda: Ommastrephidae) in the southeastern Pacific. Rev. De Biol. Mar. Y Oceanogr. 2009, 44, 227–235. [Google Scholar] [CrossRef]
- Watanabe, H.; Kubodera, T.; Ichii, T.; Kawahara, S. Feeding habits of neon flying squid Ommastrephes bartramii in the transitional region of the central North Pacific. Mar. Ecol. Prog. Ser. 2004, 266, 173–184. [Google Scholar] [CrossRef]
- Seki, M.P.; Polovina, J.J.; Kobayashi, D.R.; Bidigare, R.R.; Mitchum, G.T. An oceanographic characterization of swordfish (Xiphias gladius) longline fishing grounds in the springtime subtropical North Pacific. Fish Oceanogr. 2002, 11, 251–266. [Google Scholar] [CrossRef]
- Clarke, M.R.; Roeleveld, M.A.C. Cephalopods in the diet of sperm whales caught commercially off Durban, South Africa. Afr. J. Mar. Sci. 1998, 20, 41–45. [Google Scholar] [CrossRef]
- Yu, W.; Chen, X.; Yi, Q. Fishing ground distribution of neon flying squid (Ommastrephes bartramii) in relation to oceanographic conditions in the northwest Pacific Ocean. J. Ocean Univ. China 2017, 16, 1157–1166. [Google Scholar] [CrossRef]
- Chen, C.S.; Chiu, T.S. Variations of life history parameters in two geographical groups of the neon flying squid, Ommastrephes bartramii, from the North Pacific. Fish Res. 2003, 63, 349–366. [Google Scholar] [CrossRef]
- Yatsu, A.; Tanaka, H.; Mori, J. Population structure of the neon flying squid, Ommastrephes bartramii, in the North Pacific Ocean. In Contributed Papers to International Symposium on Large Pelagic Squids, 18–19 July 1996, for JAMARC’s 25th Anniversary of its Foundation; Japan Marine Fishery Resources Research Center: Tokyo, Japan, 1998; pp. 31–48. [Google Scholar]
- Chen, X.; Tian, S.; Liu, B.; Chen, Y. Modeling a habitat suitability index for the eastern fall cohort of Ommastrephes bartramii in the central North Pacific Ocean. Chin. J. Ocean. Limnol. 2011, 29, 493–504. [Google Scholar] [CrossRef]
- Yu, W.; Chen, X.; Yi, Q.; Gao, G.; Chen, Y. Impacts of climatic and marine environmental variations on the spatial distribution of Ommastrephes bartramii in the Northwest Pacific Ocean. Acta Ocean. Sin. 2016, 35, 108–116. [Google Scholar] [CrossRef]
- Golikov, A.V.; Ceia, F.R.; Sabirov, R.M.; Zaripova, Z.I.; Blicher, M.E.; Zakharov, D.V.; Xavier, J.C. Ontogenetic changes in stable isotope (δ13C and δ15N) values in squid Gonatus fabricii (Cephalopoda) reveal its important ecological role in the Arctic. Mar. Ecol. Prog. Ser. 2018, 606, 65–78. [Google Scholar] [CrossRef]
- Yu, W.; Chen, X.; Zhang, Y.; Yi, Q. Habitat suitability modelling revealing environmental-driven abundance variability and geographical distribution shift of winter–spring cohort of neon flying squid Ommastrephes bartramii in the northwest Pacific Ocean. ICES J. Mar. Sci. 2019, 76, 1722–1735. [Google Scholar] [CrossRef]
- Ichii, T.; Mahapatra, K.; Sakai, M.; Okada, Y. Life history of the neon flying squid: Effect of the oceanographic regime in the North Pacific Ocean. Mar. Ecol. Prog. Ser. 2009, 378, 1–11. [Google Scholar] [CrossRef]
- Hobson, K.A. Tracing origins and migration of wildlife using stable isotopes: A review. Oecologia 1999, 120, 314–326. [Google Scholar] [CrossRef]
- McCutchan, J.H., Jr.; Lewis, W.M., Jr.; Kendall, C.; McGrath, C.C. Variation in trophic shift for stable isotope ratios of carbon, nitrogen, and sulfur. Oikos 2003, 102, 378–390. [Google Scholar] [CrossRef]
- Kelly, M.H.; Hagar, W.G.; Jardine, T.D.; Cunjak, R.A. Nonlethal sampling of sunfish and slimy sculpin for stable isotope analysis: How scale and fin tissue compare with muscle tissue. N. Am. J. Fish. Manag. 2006, 26, 921–925. [Google Scholar] [CrossRef]
- Cherel, Y.; Hobson, K.A. Geographical variation in carbon stable isotope signatures of marine predators: A tool to investigate their foraging areas in the Southern Ocean. Mar. Ecol. Prog. Ser. 2007, 329, 281–287. [Google Scholar] [CrossRef]
- Minagawa, M.; Wada, E. Stepwise enrichment of 15N along food chains: Further evidence and the relation between δ15N and animal age. Geochim. Et Cosmochim. Acta. 1984, 48, 1135–1140. [Google Scholar] [CrossRef]
- Jennings, S.; Warr, K.J. Environmental correlates of large-scale spatial variation in the δ15N of marine animals. Mar. Biol. 2003, 142, 1131–1140. [Google Scholar] [CrossRef]
- Tanaka, Y.; Minami, H.; Ishihi, Y.; Kumon, K.; Higuchi, K.; Eba, T.; Nishi, A.; Nikaido, H.; Shiozawa, S. Relationship between prey utilization and growth variation in hatchery-reared Pacific bluefin tuna, Thunnus orientalis (Temminck et Schlegel), larvae estimated using nitrogen stable isotope analysis. Aquac. Res. 2014, 45, 537–545. [Google Scholar] [CrossRef]
- Hesslein, R.H.; Hallard, K.A.; Ramlal, P. Replacement of sulfur, carbon, and nitrogen in tissue of growing broad whitefish (Coregonus nasus) in response to a change in diet traced by δ34S, δ13C, and δ15N. Can. J. Fish. Aquat. Sci. 1993, 50, 2071–2076. [Google Scholar] [CrossRef]
- Gratton, C.; Forbes, A.E. Changes in δ 13C stable isotopes in multiple tissues of insect predators fed isotopically distinct prey. Oecologia 2006, 147, 615–624. [Google Scholar] [CrossRef]
- Inger, R.; Bearhop, S. Applications of stable isotope analyses to avian ecology. Ibis 2008, 150, 447–461. [Google Scholar] [CrossRef]
- Malpica-Cruz, L.; Herzka, S.Z.; Sosa-Nishizaki, O.; Lazo, J.P. Tissue-specific isotope trophic discrimination factors and turnover rates in a marine elasmobranch: Empirical and modeling results. Can. J. Fish. Aquat. Sci. 2012, 69, 551–564. [Google Scholar] [CrossRef]
- Kato, Y.; Sakai, M.; Nishikawa, H.; Igarashi, H.; Ishikawa, Y.; Vijai, D.; Sakurai, Y.; Wakabayshi, T.; Awaji, T. Stable isotope analysis of the gladius to investigate migration and trophic patterns of the neon flying squid (Ommastrephes bartramii). Fish Res. 2016, 173, 169–174. [Google Scholar] [CrossRef]
- Guerra, A.; Rodriguez-Navarro, A.B.; González, Á.F.; Romanek, C.S.; Alvarez-Lloret, P.; Pierce, G.J. Life-history traits of the giant squid Architeuthis dux revealed from stable isotope signatures recorded in beaks. ICES J. Mar. Sci. 2010, 67, 1425–1431. [Google Scholar] [CrossRef][Green Version]
- Quaeck, K. Stable Isotope Analysis of Fish Eye Lenses: Reconstruction of Ontogenetic Trends in Spatial and Trophic Ecology of Elasmobranchs and Deep-Water Teleosts. Ph.D. Thesis, University of Southampton, Southampton, UK, 2017. [Google Scholar]
- Campana, S.E. Chemistry and composition of fish otoliths: Pathways, mechanisms and applications. Mar. Ecol. Prog. Ser. 1999, 188, 263–297. [Google Scholar] [CrossRef]
- Trueman, C.N.; MacKenzie, K.M.; Palmer, M.R. Stable isotopes reveal linkages between ocean climate, plankton community dynamics, and survival of two populations of Atlantic salmon (Salmo salar). ICES J. Mar. Sci. 2012, 69, 784–794. [Google Scholar] [CrossRef]
- Tzadik, O.E.; Curtis, J.S.; Granneman, J.E.; Kurth, B.N.; Pusack, T.J.; Wallace, A.A.; Hollander, D.J.; Peebles, E.B.; Stallings, C.D. Chemical archives in fishes beyond otoliths: A review on the use of other body parts as chronological recorders of microchemical constituents for expanding interpretations of environmental, ecological, and life-history changes. Limnol. Oceanogr. Methods 2017, 15, 238–263. [Google Scholar] [CrossRef]
- Xu, W.; Chen, X.; Liu, B.; Chen, Y.; Huan, M.; Liu, N.; Lin, J. Inter-individual variation in trophic history of Dosidicus gigas, as indicated by stable isotopes in eye lenses. Aquac. Fish. 2019, 4, 261–267. [Google Scholar] [CrossRef]
- Quaeck-Davies, K.; Bendall, V.A.; MacKenzie, K.M.; Hetherington, S.; Newton, J.; Trueman, C.N. Teleost and elasmobranch eye lenses as a target for life-history stable isotope analyses. PeerJ 2018, 6, e4883. [Google Scholar] [CrossRef]
- Hunsicker, M.E.; Essington, T.E.; Aydin, K.Y.; Ishida, B. Predatory role of the commander squid Berryteuthis magister in the eastern Bering Sea: Insights from stable isotopes and food habits. Mar. Ecol. Prog. Ser. 2010, 415, 91–108. [Google Scholar] [CrossRef]
- Onthank, K.L. Exploring the Life Histories of Cephalopods Using Stable Isotope Analysis of An Archival Tissue; Washington State University: Washington, DC, USA, 2013. [Google Scholar]
- Meath, B.; Peebles, E.B.; Seibel, B.A.; Judkins, H. Stable isotopes in the eye lenses of Doryteuthis plei (Blainville 1823): Exploring natal origins and migratory patterns in the eastern Gulf of Mexico. Cont. Shelf. Res. 2019, 174, 76–84. [Google Scholar] [CrossRef]
- Miserez, A.; Schneberk, T.; Sun, C.; Zok, F.W.; Waite, J.H. The transition from stiff to compliant materials in squid beaks. Science 2008, 319, 1816–1819. [Google Scholar] [CrossRef]
- Arkhipkin, A.; Perez, J.A.A. Life history reconstruction. In Squid Recruitment Dynamics; United Nations Development Programme, Food and Agriculture Organization of the United Nations: Rome, Italy, 1998; pp. 157–180. [Google Scholar]
- Xu, W.; Liu, B.; Chen, X.; Huan, M. Research advancements on role of eye lens in reconstruction of cephalopod life history. J. Dalian Fish. Univ. 2018, 33, 408–412. (In Chinese) [Google Scholar] [CrossRef]
- Arkhipkin, A.I. Statoliths as ‘black boxes’(life recorders) in squid. Mar. Freshw. Res. 2005, 56, 573–583. [Google Scholar] [CrossRef]
- Rodhouse, P.G.; Hatfield, E.M.C. Age determination in squid using statolith growth increments. Fish Res. 1990, 8, 323–334. [Google Scholar] [CrossRef]
- Anken, R.H.; Werner, K.; Ibsch, M.; Rahmann, H. Fish inner ear otolith size and bilateral asymmetry during development. Hear. Res. 1998, 121, 77–83. [Google Scholar] [CrossRef]
- Saito, T.; Kaga, T.; Seki, J.; Otake, T. Otolith microstructure of chum salmon Oncorhynchus keta: Formation of sea entry check and daily deposition of otolith increments in seawater conditions. Fish. Sci. 2007, 73, 27–37. [Google Scholar] [CrossRef]
- Bigelow, K.A.; Landgraf, K.C. Hatch Dates and Growth of Ommastrephes bartramii Paralarvae from Hawaiian Waters as Determined from Statolith Analysis. In Recent Advances in Cephalopod Fisheries Biology; Tokai University Press: Tokyo, Japan, 1993; pp. 15–24. [Google Scholar]
- Parry, M. Trophic variation with length in two ommastrephid squids, Ommastrephes bartramii and Sthenoteuthis oualaniensis. Mar. Biol. 2008, 153, 249–256. [Google Scholar] [CrossRef]
- Wang, Y.; Han, P.; Chen, X.; Fang, Z. Interannual and ontogenetic stage differences on trophic ecology of neon flying squid Ommastrephes bartramii inferred from stable isotope analyses in beaks. Fish Res. 2022, 249, 106252. [Google Scholar] [CrossRef]
- Ehrhardt, N.M. On the fishery and biology of the giant squid Dosidicus gigas in the Gulf of California, Mexico, in ”Advanced in Assessment of World Cephalopod Resources”. FAO Fish. Tech. Pap. 1983, 231, 306–339. [Google Scholar]
- Dawe, E.G.; Natsukari, Y. Light microscopy. In Squid Age Determination Using Statoliths, Proceedings of the International Workshop, Mazara del Vallo, Italy, 9–14 October 1989; Jereb, P., Ragonese, S., BoletzkyVon, S., Eds.; Instituto di Tecnologiadella Pesca e del Pescato: Mazara del Vallo, Italy, 1991; pp. 83–96, NTR–ITPP Special Publications, No. 1. [Google Scholar]
- Uozumi, Y.; Ohara, H. Age and Growth of Nototodarus sloanii (Cephalopoda: Oegopsida) Based on Daily Increment Counts in Statoliths. Nippon Suisan Gakkaishi 1993, 59, 1469–1477. [Google Scholar] [CrossRef]
- Jin, Y.; Lin, F.; Chen, X.; Liu, B.; Li, J. Microstructure comparison of hard tissues (statoliths, beaks, and eye lenses) of Uroteuthis chinensis in the South China Sea. Bull. Mar. Sci. 2019, 95, 13–26. [Google Scholar] [CrossRef]
- Liu, B.; Chen, X.; Zhong, J. Age, growth and population strcture of squid Sthenoteuthis oualaniensis in northwest Indian Ocean by statolith microstructure. Chin. J. Dalian Fish. Univ. 2009, 24, 206–212. (In Chinese) [Google Scholar] [CrossRef]
- Post, D.M.; Layman, C.A.; Arrington, D.A.; Takimoto, G.; Quattrochi, J.; Montana, C.G. Getting to the fat of the matter: Models, methods and assumptions for dealing with lipids in stable isotope analyses. Oecologia 2007, 152, 179–189. [Google Scholar] [CrossRef]
- Jackson, A.L.; Inger, R.; Parnell, A.C.; Bearhop, S. Comparing isotopic niche widths among and within communities: SIBER–Stable Isotope Bayesian Ellipses in R. J Anim. Ecol. 2011, 80, 595–602. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Layman, C.A.; Arrington, D.A.; Montaña, C.G.; Post, D.M. Can stable isotope ratios provide for community-wide measures of trophic structure? Ecology 2007, 88, 42–48. [Google Scholar] [CrossRef]
- Layman, C.A.; Araujo, M.S.; Boucek, R.; Hammerschlag-Peyer, C.M.; Harrison, E.; Jud, Z.R.; Matich, P.; Rosenblatt, A.E.; Vaudo, J.J.; Yeager, L.A.; et al. Applying stable isotopes to examine food-web structure: An overview of analytical tools. Biol. Rev. 2012, 87, 545–562. [Google Scholar] [CrossRef] [PubMed]
- Langton, R.W. Diet overlap between Atlantic cod, Gadus morhua, silver hake, Merluccius bilinearis, and fifteen other northwest Atlantic finfish. Fish. Bull. 1982, 80, 45–759. [Google Scholar]
- Moreno, R.; Jover, L.; Velando, A.; Munilla, I.; Sanpera, C. Influence of trophic ecology and spatial variation on the isotopic fingerprints of seabirds. Mar. Ecol. Prog. Ser. 2011, 442, 229–239. [Google Scholar] [CrossRef]
- Faletti, M.E.; Stallings, C.D. Life history through the eyes of a hogfish: Trophic growth and differential juvenile habitat use from stable isotope analysis. Mar. Ecol. Prog. Ser. 2021, 666, 183–202. [Google Scholar] [CrossRef]
- Watanabe, H.; Kubodera, T.; Ichii, T.; Sakai, M.; Moku, M.; Seitou, M. Diet and sexual maturation of the neon flying squid Ommastrephes bartramii during autumn and spring in the Kuroshio–Oyashio transition region. J. Mar. Biol. Assoc. UK 2008, 88, 381–389. [Google Scholar] [CrossRef]
- Sassa, C.; Moser, H.G.; Kawaguchi, K. Horizontal and vertical distribution patterns of larval myctophid fishes in the Kuroshio Current region. Fish. Oceanogr. 2002, 11, 1–10. [Google Scholar] [CrossRef]
- Moku, M.; Tsuda, A.; Kawaguchi, K. Spawning season and migration of the myctophid fish Diaphus theta in the western North Pacific. Ichthyol. Res. 2003, 50, 0052–0058. [Google Scholar] [CrossRef]
- Fang, Z.; Thompson, K.; Jin, Y.; Chen, X.; Chen, Y. Preliminary analysis of beak stable isotopes (δ13C and δ15N) stock variation of neon flying squid, Ommastrephes bartramii, in the North Pacific Ocean. Fish. Res. 2016, 177, 153–163. [Google Scholar] [CrossRef]
- Lorrain, A.; Argüelles, J.; Alegre, A.; Bertrand, A.; Munaron, J.M.; Richard, P.; Cherel, Y. Sequential isotopic signature along gladius highlights contrasted individual foraging strategies of jumbo squid (Dosidicus gigas). PLoS ONE 2011, 6, e22194. [Google Scholar] [CrossRef] [PubMed]
- Bower, J.R.; Ichii, T. The red flying squid (Ommastrephes bartramii): A review of recent research and the fishery in Japan. Fish. Res. 2005, 76, 39–55. [Google Scholar] [CrossRef]
- Chen, P.; Fang, Z.; Chen, X. Population identification of Ommastrephes bartramii based on beak’s external morphological characters. Mar. Fish. 2015, 37, 1–9. (In Chinese) [Google Scholar] [CrossRef]
- Ichii, T.; Mahapatra, K.; Sakai, M.; Inagake, D.; Okada, Y. Differing body size between the autumn and the winter–spring cohorts of neon flying squid (Ommastrephes bartramii) related to the oceanographic regime in the North Pacific: A hypothesis. Fish Oceanogr. 2004, 13, 295–309. [Google Scholar] [CrossRef]
- Yatsu, A.; Midorikawa, S.; Shimada, T.; Uozumi, Y. Age and growth of the neon flying squid, Ommastrephes bartramii, in the North Pacific Ocean. Fish. Res. 1997, 29, 257–270. [Google Scholar] [CrossRef]
- Yu, W.; Chen, X.; Yi, Q.; Tian, S. A review of interaction between neon flying squid (Ommastrephes bartramii) and oceanographic variability in the North Pacific Ocean. J. Ocean Univ. China 2015, 14, 739–748. [Google Scholar] [CrossRef]
- Wang, W.; Shao, Q.; Xue, Y.; Ji, T. On the relationship between the resources of Ommastrephes bartrami and marine environment in the Northwest Pacific Ocean based on GIS. J. Geogr. Inf. Sci. 2003, 5, 39–44. [Google Scholar]
- Parrish, R.H.; Nelson, C.S.; Bakun, A. Transport mechanisms and reproductive success of fishes in the California Current. Biol. Oceanogr. 1981, 1, 175–203. [Google Scholar] [CrossRef]
- Fang, Z. Fisheries Ecology of Neon Flying Squid Ommastrephes bartramii in North Pacific Ocean based on beak. Ph.D. Thesis, University of Shanghai Ocean, Shanghai, China, 2016. [Google Scholar]
- Somes, C.J.; Schmittner, A.; Galbraith, E.D.; Lehmann, M.F.; Altabet, M.A.; Montoya, J.P.; Letelier, R.M.; Mix, A.C.; Bourbonnais, A.; Eby, M. Simulating the global distribution of nitrogen isotopes in the ocean. Glob. Biogeochem. Cycles 2010, 24, GB4019. [Google Scholar] [CrossRef]
- Parry, M. Feeding behavior of two ommastrephid squids Ommastrephes bartramii and Sthenoteuthis oualaniensis off Hawaii. Mar. Ecol. Prog. Ser. 2006, 318, 229–235. [Google Scholar] [CrossRef]
- Jin, Y.; Chen, X. Information in beaks of Ommastrephes bartramii in the North Pacific Ocean using stable isotope technology. Chin. J. Ecology 2014, 33, 2101–2107, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Rodhouse, P.G.; Nigmatullin, C.M. Role as consumers. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 1996, 351, 1003–1022. [Google Scholar] [CrossRef]
- Ichii, T.; Mahapatra, K.; Sakai, M.; Wakabayashi, T.; Okamura, H.; Igarashi, H.; Inagake, D.; Okada, Y. Changes in abundance of the neon flying squid Ommastrephes bartramii in relation to climate change in the central North Pacific Ocean. Mar. Ecol. Prog. Ser. 2011, 441, 151–164. [Google Scholar] [CrossRef]
- Sauer, W.H.H.; Lipiński, M.R. Food of squid Loligo vulgaris reynaudii (Cephalopoda: Loliginidae) on their spawning grounds off the Eastern Cape, South Africa. S. Afr. J. Mar. Sci. 1991, 10, 193–201. [Google Scholar] [CrossRef]
- Carlisle, A.B.; Goldman, K.J.; Litvin, S.Y.; Madigan, D.J.; Bigman, J.S.; Swithenbank, A.M.; Kline, T.C.; Block, B.A. Stable isotope analysis of vertebrae reveals ontogenetic changes in habitat in an endothermic pelagic shark. Proc. R. Soc. B-Biol. Sci. 2015, 282, 20141446. [Google Scholar] [CrossRef]
- Zhang, G.; Xie, S.C. Development of niche concept and its perspectives. Chin. J. Ecology 1997, 16, 46–51. [Google Scholar]
- Hurlbert, S.H. The measurement of niche overlap and some relatives. Ecology 1978, 591, 67–77. [Google Scholar] [CrossRef]
- Bearhop, S.; Adams, C.E.; Waldron, S.; Fuller, R.A.; MacLeod, H. Determining trophic niche width: A novel approach using stable isotope analysis. J. Anim. Ecol. 2004, 73, 1007–1012. [Google Scholar] [CrossRef]
- Hjelm, J.; Persson, L.; Christensen, B. Growth, morphological variation and ontogenetic niche shifts in perch (Percafluviatilis) in relation to resource availability. Oecologia 2000, 122, 190–199. [Google Scholar] [CrossRef]
- Werner, E.E.; Gilliam, J.F. The ontogenetic niche and species interactions in size-structured populations. Annu. Rev. Ecol. Syst. 1984, 15, 393–425. [Google Scholar] [CrossRef]
- Fang, Z.; Chen, X.J.; Lu, H.J.; Li, J.H.; Liu, B.L. Morphology and growth of beaks in two cohorts for neon flying squid (Ommastrephes bartramii) in the North Pacific Ocean. Acta Ecol. Sin. 2014, 34, 5405–5415. [Google Scholar]
- Bartol, I.K.; Krueger, P.S.; Thompson, J.T.; Stewart, W.J. Swimming dynamics and propulsive efficiency of squids throughout ontogeny. Integr. Comp. Biol. 2008, 48, 720–733. [Google Scholar] [CrossRef]
- Bethea, D.M.; Buckel, J.A.; Carlson, J.K. Foraging ecology of the early life stages of four sympatric shark species. Mar. Ecol. Prog. Ser. 2004, 268, 245–264. [Google Scholar] [CrossRef]
- Camarillo-Coop, S.; Salinas-Zavala, C.A.; Lavaniegos, B.E.; Markaida, U. Food in early life stages of Dosidicus gigas (Cephalopoda: Ommastrephidae) from the Gulf of California, Mexico. J. Mar. Biol. Assoc. UK 2013, 93, 1903–1910. [Google Scholar] [CrossRef]
- Nigmatullin, C.M.; Nesis, K.N.; Arkhipkin, A.I. A review of the biology of the jumbo squid Dosidicus gigas (Cephalopoda: Ommastrephidae). Fish. Res. 2001, 54, 9–19. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Location | Sex | N | Mantle Length (mm) | Lens Diameter (mm) | ||
|---|---|---|---|---|---|---|---|
| Mean ± SD | Range | Mean ± SD | Range | ||||
| Eastern stock | 173°11′–175°36′ W, 39°33′–41°32′ N | Female | 11 | 461.73 ± 70.64 | 357~534 | 14.23 ± 1.64 | 11.95~16.43 |
| Male | 4 | 217.25 ± 29.95 | 176~246 | 6.35 ± 1.50 | 4.26~7.71 | ||
| Western stock | 149°25′–161°04′ E, 39°42′–44°47′ N | Female | 12 | 381.50 ± 65.64 | 299~490 | 11.13 ± 1.40 | 8.94~14.09 |
| Male | 13 | 292.00 ± 21.42 | 259~329 | 10.05 ± 1.49 | 7.44~12.04 | ||
| ID | Collection Date | Hatching Date | Location | ML | Sex | Age (days) | |||
|---|---|---|---|---|---|---|---|---|---|
| Emb | Par | Juv | Sub-Adu | ||||||
| 5 | 1 November 2020 | 3 March 2020 | 156°26′ E, 43°39′ N | 490 | F | 0 | 22 | 84 | 243 |
| 9 | 15 October 2020 | 14 April 2020 | 156°50′ E, 44°23′ N | 325 | F | 0 | 20 | 67 | 184 |
| 15 | 15 October 2020 | 9 April 2020 | 156°50′ E, 44°23′ N | 329 | M | 0 | 16 | 83 | 189 |
| 19 | 15 October 2020 | 3 May 2020 | 156°50′ E, 44°23′ N | 275 | M | 0 | 34 | 110 | 165 |
| 21 | 15 October 2020 | 18 May 2020 | 156°50′ E, 44°23′ N | 270 | M | 0 | 22 | 73 | 150 |
| 24 | 15 October 2020 | 23 April 2020 | 156°50′ E, 44°23′ N | 294 | M | 0 | 25 | 66 | 175 |
| 25 | 23 May 2020 | 18 October 2019 | 175°36′ W, 39°33′ N | 523 | F | 0 | 25 | 71 | 218 |
| 37 | 16 July 2020 | 8 December 2020 | 174°29′ W, 41°23′ N | 364 | F | 0 | 23 | 64 | 221 |
| 45 | 31 October 2020 | 20 April 2020 | 156°26′ E, 43°39′ N | 416 | F | 0 | 87 | 194 | |
| 48 | 31 October 2020 | 2 April 2020 | 156°26′ E, 43°39′ N | 485 | F | 0 | 19 | 93 | 212 |
| 49 | 11 August 2020 | 19 February 2020 | 156°40′ E, 39°57′ N | 291 | M | 0 | 27 | 88 | 174 |
| 56 | 11 August 2020 | 1 January 2020 | 156°40′ E, 39°57′ N | 348 | F | 0 | 20 | 77 | 223 |
| 57 | 11 August 2020 | 25 January 2020 | 156°40′ E, 39°57′ N | 320 | M | 0 | 88 | 199 | |
| 71 | 19 November 2020 | 9 June 2020 | 149°25′ E, 39°42′ N | 385 | F | 0 | 39 | 102 | 163 |
| 74 | 19 November 2020 | 3 April 2020 | 149°25′ E, 39°42′ N | 376 | F | 0 | 15 | 85 | 230 |
| 77 | 19 November 2020 | 5 March 2020 | 149°25′ E, 39°42′ N | 341 | F | 0 | 28 | 87 | 259 |
| 91 | 16 June 2020 | 14 December 2019 | 173°52′ W, 40°55′ N | 534 | F | 0 | 29 | 81 | 185 |
| 94 | 16 June 2020 | 14 September 2019 | 173°52′ W, 40°55′ N | 508 | F | 0 | 31 | 77 | 276 |
| 95 | 16 June 2020 | 23 September 2019 | 173°52′ W, 40°55′ N | 518 | F | 0 | 31 | 73 | 267 |
| 100 | 20 October 2020 | 16 May 2020 | 156°20′ E, 44°10′ N | 364 | F | 0 | 30 | 70 | 157 |
| 111 | 26 September 2020 | 27 March 2020 | 161°04′ E, 44°06′ N | 301 | M | 0 | 32 | 107 | 183 |
| 113 | 26 September 2020 | 28 March 2020 | 161°04′ E, 44°06′ N | 302 | F | 0 | 40 | 71 | 182 |
| 114 | 26 September 2020 | 15 April 2020 | 161°04′ E, 44°06′ N | 297 | M | 0 | 31 | 87 | 164 |
| 118 | 26 September 2020 | 9 April 2020 | 161°04′ E, 44°06′ N | 277 | M | 0 | 24 | 78 | 170 |
| 145 | 7 July 2020 | 23 February 2020 | 174°44′ W, 41°32′ N | 176 | M | 0 | 26 | 87 | 135 |
| 152 | 7 July 2020 | 28 February 2020 | 174°44′ W, 41°32′ N | 246 | M | 0 | 41 | 69 | 130 |
| 155 | 7 July 2020 | 9 February 2020 | 174°44′ W, 41°32′ N | 217 | M | 0 | 32 | 89 | 149 |
| 158 | 7 July 2020 | 15 January 2020 | 174°44′ W, 41°32′ N | 230 | M | 0 | 29 | 70 | 174 |
| 161 | 20 June 2020 | 23 September 2019 | 174°31′ W, 40°25′ N | 497 | F | 0 | 35 | 73 | 271 |
| 172 | 14 July 2020 | 29 January 2020 | 174°14′ W, 40°16′ N | 357 | F | 0 | 38 | 77 | 167 |
| 176 | 14 July 2020 | 20 November 2019 | 174°14′ W, 40°16′ N | 394 | F | 0 | 99 | 237 | |
| 178 | 11 June 2020 | 1 November 2019 | 174°04′ W, 40°54′ N | 495 | F | 0 | 37 | 107 | 223 |
| 183 | 24 September 2020 | 7 May 2020 | 160°11′ E, 44°47′ N | 267 | M | 0 | 29 | 76 | 140 |
| 186 | 24 September 2020 | 4 May 2020 | 160°11′ E, 44°47′ N | 259 | M | 0 | 41 | 103 | 143 |
| 188 | 24 September 2020 | 5 May 2020 | 160°11′ E, 44°47′ N | 299 | F | 0 | 32 | 87 | 142 |
| 189 | 24 September 2020 | 6 March 2020 | 160°11′ E, 44°47′ N | 311 | M | 0 | 41 | 79 | 202 |
| 192 | 24 September 2020 | 6 March 2020 | 160°11′ E, 44°47′ N | 305 | M | 0 | 98 | 202 | |
| 204 | 6 June 2020 | 1 November 2019 | 175°25′ W, 40°55′ N | 383 | F | 0 | 30 | 101 | 218 |
| 207 | 3 December 2020 | 21 February 2020 | 153°36′ E, 42°13′ N | 447 | F | 0 | 32 | 76 | 256 |
| 214 | 30 May 2020 | 8 September 2019 | 173°11′ W, 40°09′ N | 506 | F | 0 | 34 | 98 | 265 |
| ID | N | δ13C/‰ | δ15N/‰ | δ15N/δ13C | ||||
|---|---|---|---|---|---|---|---|---|
| Mean ± SD | Range | Mean ± SD | Range | Slope | p-Value | R2 | ||
| 5 | 8 | −19.81 ± 0.34 | −20.38~−19.47 | 7.44 ± 1.59 | 4.56~9.13 | 4.083 | 0.05 * | 0.753 |
| 9 | 7 | −19.90 ± 0.17 | −20.19~−19.68 | 7.93 ± 1.68 | 4.97~9.43 | 7.038 | 0.695 | 0.483 |
| 15 | 7 | −19.78 ± 0.17 | −20.19~−19.68 | 7.79 ± 0.51 | 4.89~9.64 | 2.978 | 0.001 ** | 0.904 |
| 19 | 6 | −19.73 ± 0.30 | −20.14~−19.40 | 8.32 ± 1.89 | 5.09~9.85 | 5.538 | 0.022 * | 0.770 |
| 21 | 7 | −19.66 ± 0.08 | −19.78~−19.59 | 8.37 ± 1.64 | 5.19~10.06 | 6.784 | 0.487 | 0.101 |
| 24 | 6 | −19.52 ± 0.33 | −19.92~−19.08 | 8.02 ± 1.79 | 4.52~9.55 | 4.1 | 0.081 | 0.574 |
| 25 | 8 | −19.66 ± 0.43 | −20.66~−19.13 | 8.10 ± 0.87 | 6.34~8.88 | 1.152 | 0.143 | 0.322 |
| 37 | 8 | −19.74 ± 0.44 | −20.34~−19.20 | 7.60 ± 1.85 | 4.29~9.54 | 4.005 | 0.000 ** | 0.892 |
| 45 | 6 | −19.80 ± 0.17 | −20.16~−19.62 | 8.10 ± 1.17 | 5.77~9.05 | 5.956 | 0.018 * | 0.789 |
| 48 | 7 | −19.56 ± 0.20 | −20.00~−19.46 | 7.50 ± 1.60 | 4.55~8.76 | 6.357 | 0.031 * | 0.641 |
| 49 | 6 | −19.56 ± 0.22 | −19.85~−19.15 | 7.27 ± 1.75 | 4.22~9.07 | 5.841 | 0.087 | 0.560 |
| 56 | 7 | −19.46 ± 0.09 | −19.60~−19.33 | 7.46 ± 1.60 | 4.12~8.51 | 13.736 | 0.028 * | 0.650 |
| 57 | 6 | −19.62 ± 0.28 | −20.13~−19.24 | 7.77 ± 1.64 | 4.92~9.30 | 4.294 | 0.098 | 0.536 |
| 71 | 7 | −19.69 ± 0.26 | −20.26~−19.49 | 6.99 ± 1.94 | 3.79~8.80 | 0.991 | 0.780 | 0.017 |
| 74 | 7 | −19.62 ± 0.24 | −19.95~−19.17 | 7.51 ± 1.97 | 3.76~9.54 | −2.088 | 0.581 | 0.065 |
| 77 | 7 | −19.86 ± 0.46 | −20.63~−19.33 | 7.38 ± 1.71 | 4.49~9.52 | 3.715 | 0.000 ** | 0.989 |
| 91 | 8 | −19.61 ± 0.23 | −19.82~−19.06 | 8.06 ± 1.46 | 4.59~9.19 | 0.041 | 0.988 | 0.000 |
| 94 | 8 | −19.91 ± 0.32 | −20.45~−19.55 | 7.56 ± 1.51 | 5.12~9.09 | 4.365 | 0.001 ** | 0.836 |
| 95 | 9 | −19.45 ± 0.17 | −19.60~−19.03 | 8.03 ± 1.24 | 5.03~9.05 | −2.361 | 0.407 | 0.100 |
| 100 | 7 | −19.45 ± 0.09 | −19.62~−19.34 | 7.53 ± 1.91 | 3.67~8.99 | 0.342 | 0.971 | 0.000 |
| 111 | 7 | −19.54 ± 0.38 | −19.87~−18.89 | 8.19 ± 1.83 | 4.22~9.65 | 2.823 | 0.170 | 0.339 |
| 113 | 7 | −19.68 ± 0.13 | −19.90~−19.47 | 8.23 ± 1.93 | 4.56~9.94 | 7.902 | 0.215 | 0.288 |
| 114 | 6 | −19.57 ± 0.37 | −20.33~−19.24 | 8.77 ± 1.66 | 5.29~9.94 | 3.545 | 0.059 | 0.632 |
| 118 | 7 | −19.61 ± 0.43 | −20.28~−19.20 | 7.90 ± 1.94 | 5.03~9.78 | 4.194 | 0.003 ** | 0.853 |
| 145 | 3 | −19.59 ± 0.15 | −19.80~−19.45 | 5.50 ± 1.17 | 3.92~6.74 | 6.725 | 0.345 | 0.734 |
| 152 | 5 | −19.68 ± 0.24 | −19.94~−19.29 | 6.6 ± 1.58 | 4.04~8.51 | 5.548 | 0.064 | 0.733 |
| 155 | 5 | −19.83 ± 0.23 | −20.13~−19.41 | 5.88 ± 1.49 | 3.88~8.25 | 5.897 | 0.021 * | 0.869 |
| 158 | 5 | −19.67 ± 0.42 | −20.35~−19.07 | 6.43 ± 1.58 | 4.26~8.74 | 3.64 | 0.003 ** | 0.961 |
| 161 | 9 | −19.78 ± 0.32 | −20.60~−19.47 | 7.56 ± 1.41 | 5.40~9.11 | 3.053 | 0.040 * | 0.475 |
| 172 | 6 | −19.60 ± 0.32 | −20.09~−19.26 | 7.47 ± 1.69 | 4.38~9.15 | 5.007 | 0.002 ** | 0.930 |
| 176 | 8 | −19.81 ± 0.45 | −20.60~−19.17 | 7.94 ± 1.15 | 5.93~9.26 | 2.26 | 0.004 ** | 0.779 |
| 178 | 8 | −19.81 ± 0.27 | −20.28~−19.54 | 8.03 ± 1.26 | 5.25~9.19 | 4.001 | 0.006 ** | 0.744 |
| 183 | 6 | −19.59 ± 0.18 | −19.82~−19.36 | 7.86 ± 1.97 | 4.54~9.77 | 2.795 | 0.624 | 0.066 |
| 186 | 5 | −19.59 ± 0.21 | −20.00~−19.39 | 7.84 ± 1.72 | 5.30~9.41 | 6.528 | 0.092 | 0.666 |
| 188 | 8 | −19.72 ± 0.16 | −20.02~−19.56 | 6.73 ± 2.17 | 3.74~9.16 | 6.626 | 0.207 | 0.25 |
| 189 | 7 | −19.72 ± 0.49 | −20.46~−19.21 | 7.28 ± 1.95 | 4.15~8.94 | 3.327 | 0.019 * | 0.698 |
| 192 | 6 | −19.70 ± 0.30 | −20.26~−19.29 | 7.89 ± 1.19 | 5.55~8.94 | 3.439 | 0.031 * | 0.728 |
| 204 | 7 | −19.71 ± 0.14 | −19.90~−19.52 | 7.05 ± 1.59 | 4.06~8.81 | −0.502 | 0.927 | 0.002 |
| 207 | 8 | −19.91 ± 0.51 | −20.79~−19.14 | 6.22 ± 1.06 | 4.79~7.73 | −0.384 | 0.663 | 0.034 |
| 214 | 8 | −19.67 ± 0.23 | −20.03~−19.35 | 7.93 ± 1.42 | 5.19~9.06 | 4.775 | 0.027 * | 0.585 |
| Group | Stage | δ13C/‰ | δ15N/‰ | ||
|---|---|---|---|---|---|
| Mean ± SD | Range | Mean ± SD | Range | ||
| Western stock | Paralarvae | −19.90 ± 0.31 b | −20.63~−19.36 | 4.56 ± 0.64 b | 3.67~6.34 |
| Juvenile | −19.88 ± 0.35 b | −20.55~−19.14 | 6.33 ± 1.39 b | 3.76~9.85 | |
| Subadult–adult | −19.55 ± 0.26 a | −20.79~−18.89 | 8.80 ± 0.82 a | 5.33~10.06 | |
| Eastern stock | Paralarvae | −20.06 ± 0.33 b | −20.60~−19.49 | 4.70 ± 0.61 b | 3.88~5.54 |
| Juvenile | −19.81 ± 0.39 b | −20.60~−19.03 | 6.36 ± 0.88 b | 4.87~8.71 | |
| Subadult–adult | −19.59 ± 0.24 a | −20.66~−19.07 | 8.51 ± 0.66 a | 6.34~9.54 | |
| Group | Stage | Paralarval | Juvenile | Subadult–Adult |
|---|---|---|---|---|
| Eastern stock | Paralarval | 0.65‰2 | ||
| Juvenile | 0.24 | 1.06‰2 | ||
| Subadult–adult | 0.00 | 0.14 | 0.43‰2 | |
| Western stock | Paralarval | 0.64‰2 | ||
| Juvenile | 0.29 | 1.52‰2 | ||
| Subadult–adult | 0.00 | 0.21 | 0.59‰2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Liu, B.; Hu, S.; Chen, X.; Zhang, X. Ontogenetic Trophic Shifts by Ommastrephes bartramii in the North Pacific Ocean Based on Eye Lens Stable Isotopes. Fishes 2022, 7, 295. https://doi.org/10.3390/fishes7050295
Zhang J, Liu B, Hu S, Chen X, Zhang X. Ontogenetic Trophic Shifts by Ommastrephes bartramii in the North Pacific Ocean Based on Eye Lens Stable Isotopes. Fishes. 2022; 7(5):295. https://doi.org/10.3390/fishes7050295
Chicago/Turabian StyleZhang, Jiaqi, Bilin Liu, Song Hu, Xinjun Chen, and Xinhao Zhang. 2022. "Ontogenetic Trophic Shifts by Ommastrephes bartramii in the North Pacific Ocean Based on Eye Lens Stable Isotopes" Fishes 7, no. 5: 295. https://doi.org/10.3390/fishes7050295
APA StyleZhang, J., Liu, B., Hu, S., Chen, X., & Zhang, X. (2022). Ontogenetic Trophic Shifts by Ommastrephes bartramii in the North Pacific Ocean Based on Eye Lens Stable Isotopes. Fishes, 7(5), 295. https://doi.org/10.3390/fishes7050295