
Population Structure of Wild Schizothorax kozlovi in the Upper Yangtze River Based on mtDNA and Stable Isotopes, and Their Relationship with Ambient Temperature



Abstract
1. Introduction
2. Materials and Methods
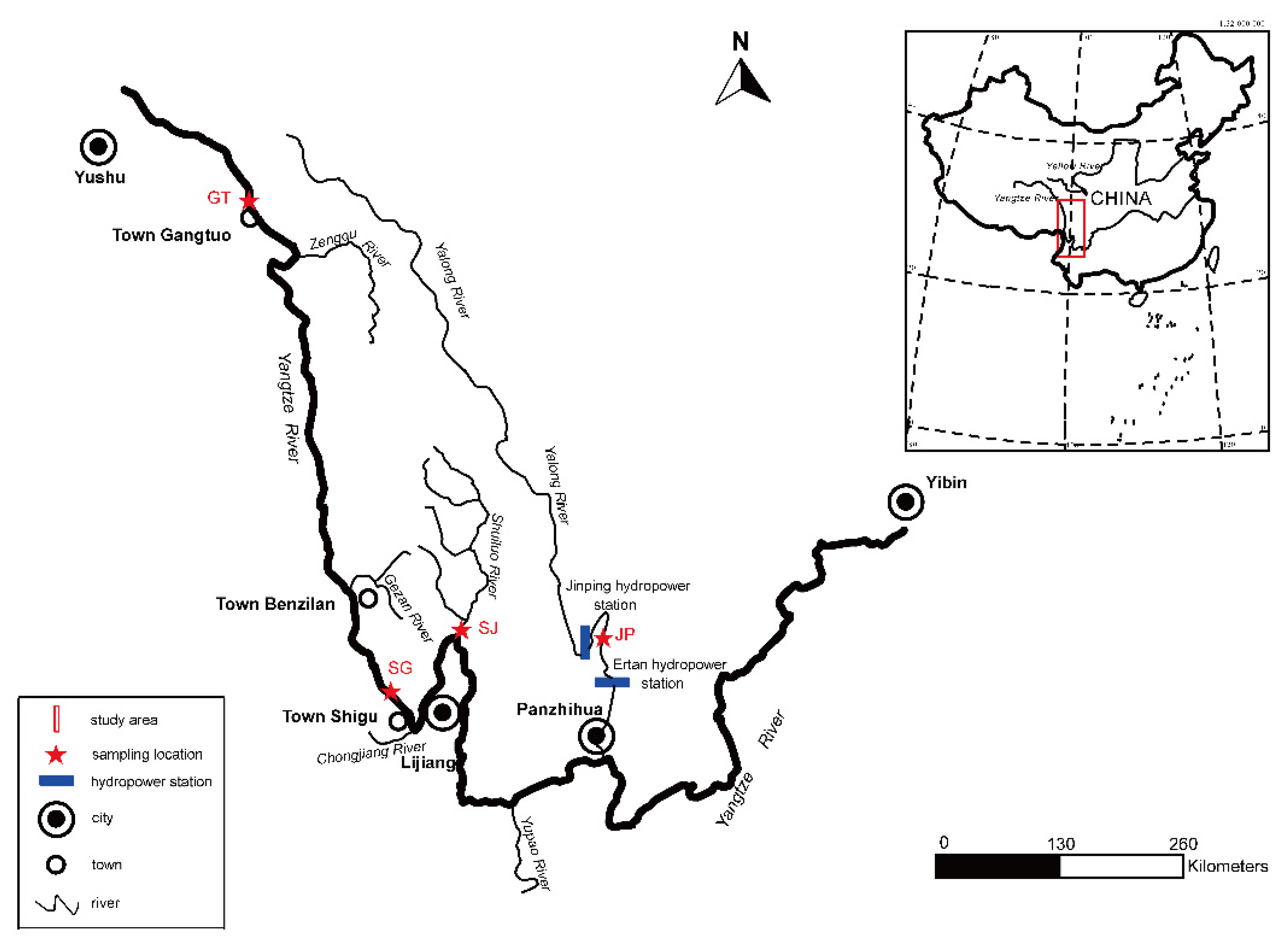
2.1. Field Sampling
2.2. Isotope Analysis
2.3. DNA Extraction, Amplification and Sequencing
2.4. Data Analysis
3. Results
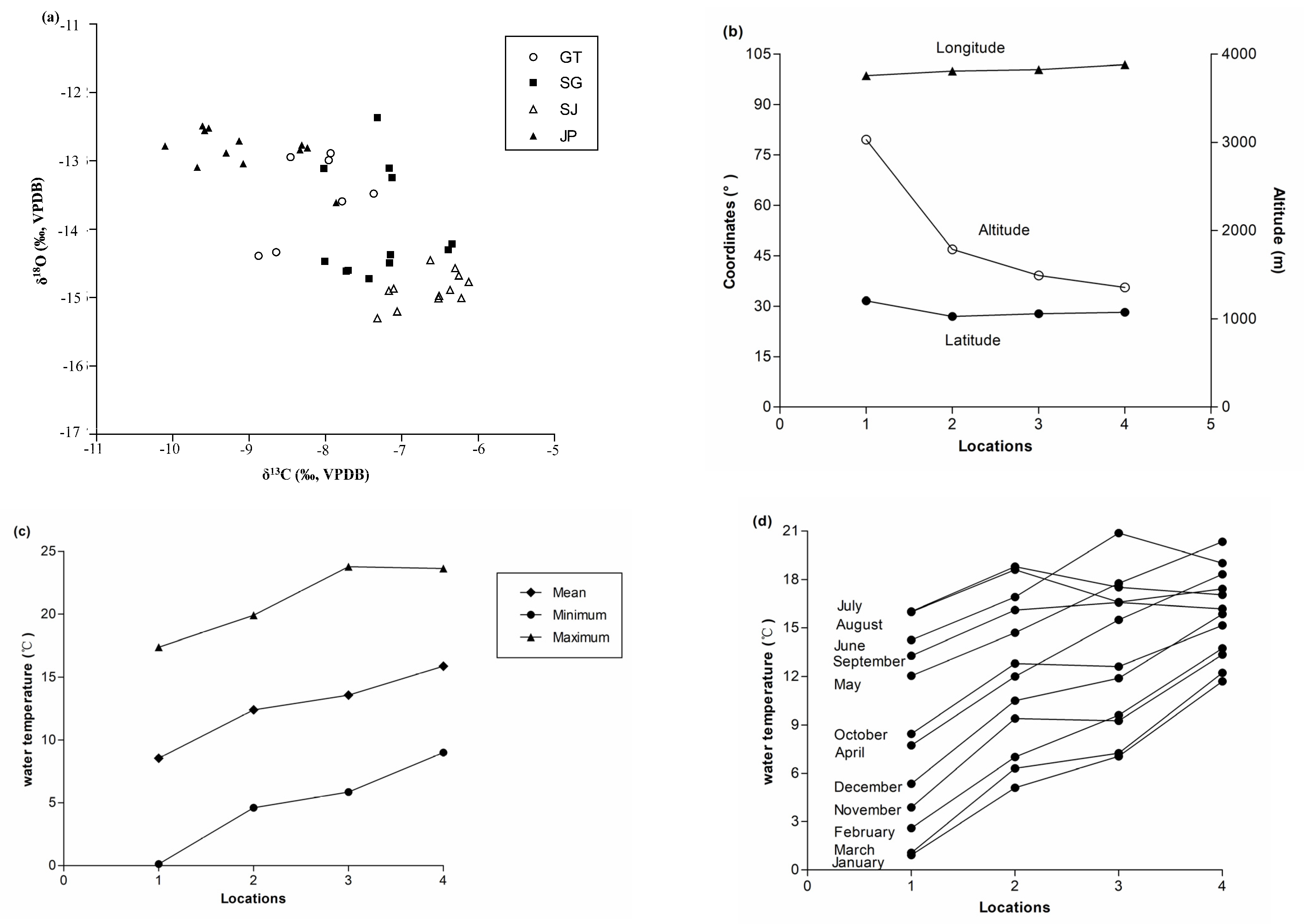
3.1. Stable Isotope Ratios of S. kozlovi Otolith
3.2. Correlations between Ambient Temperature and Otolith Stable Isotope Ratios
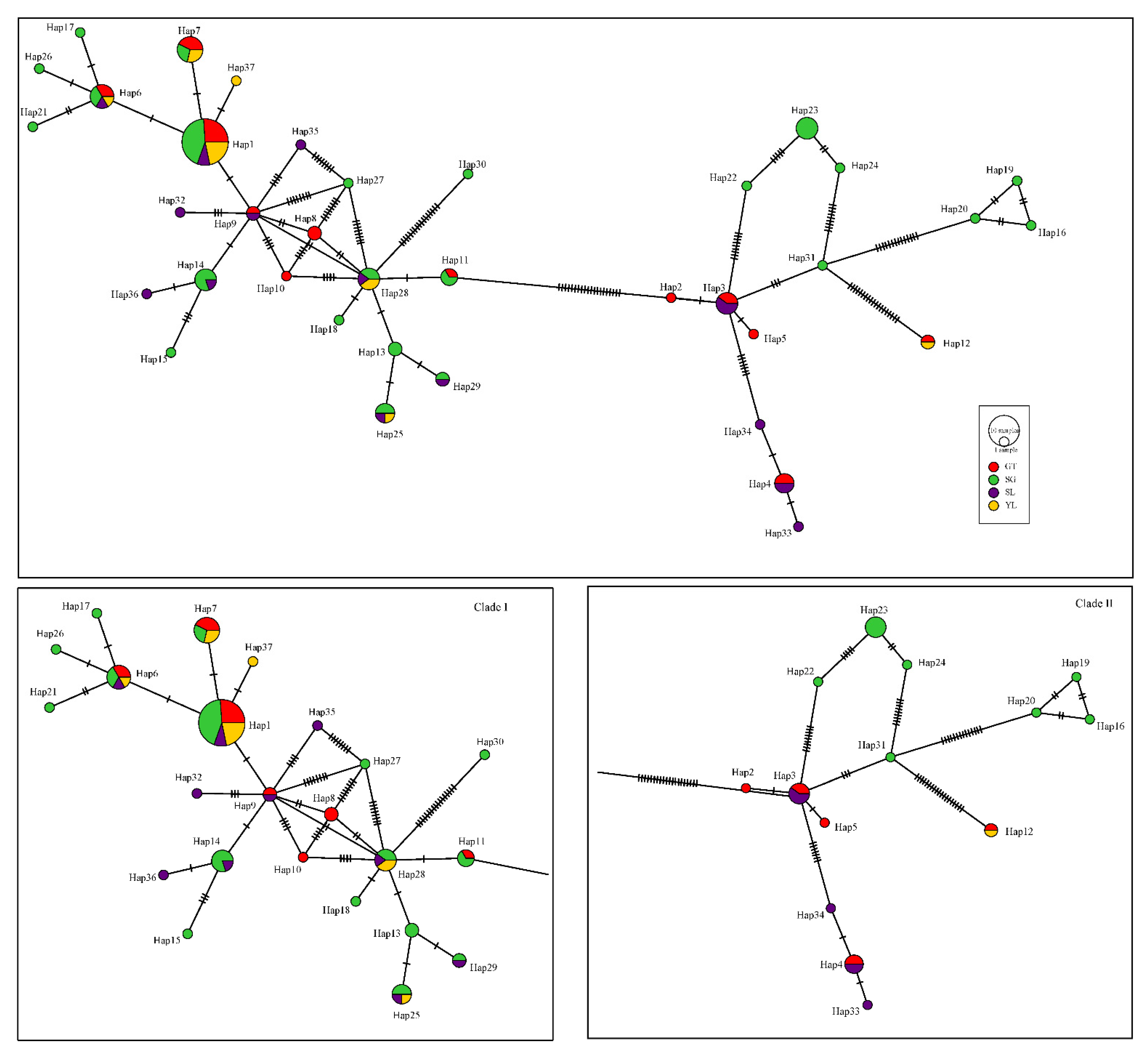
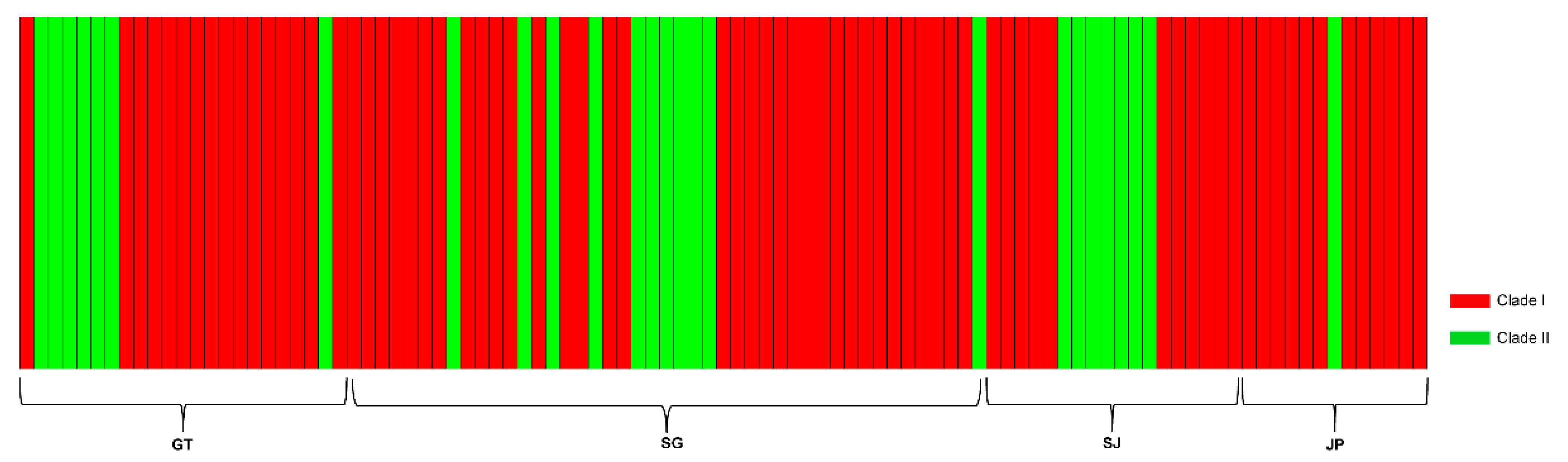
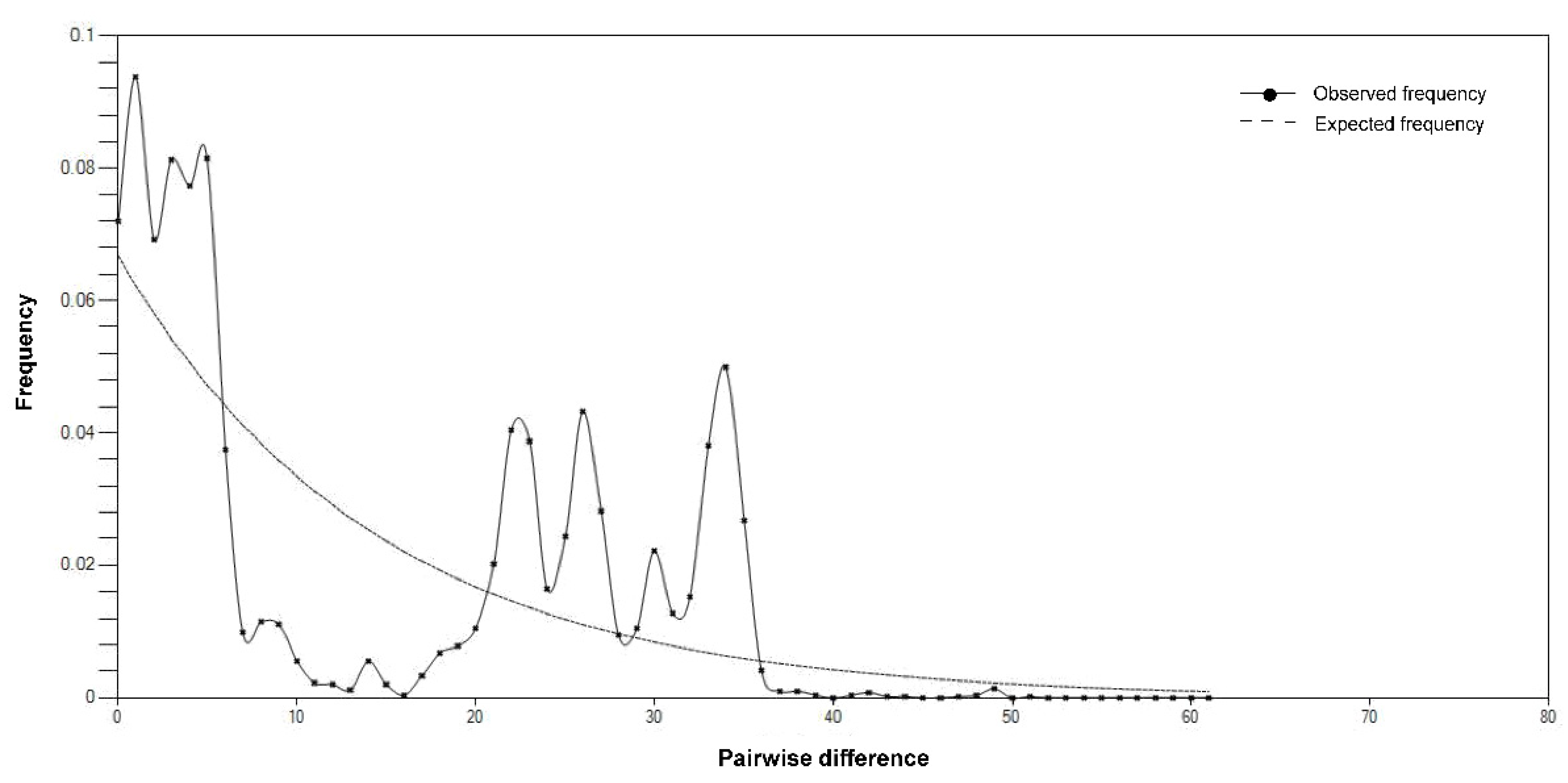
3.3. Genetic Diversity and Structure
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, Y.X. The genetic characterization and population genetic diversity of Schizothorax kozlovi (Nikolsky). Ph.D. Thesis, Sichuan Agricultural University, Ya’an, China, 2013. [Google Scholar]
- Wu, J.M.; Zhao, H.T.; Miao, Z.G.; Chen, Y.X.; Zhang, F.T.; Wang, J.W. Status and conservation of fish resources in the Chishui River. Biodiv. Sci. 2010, 18, 162–168. [Google Scholar]
- Wu, L. The Fishes of Guizhou; Guizhou People’s Press: Guiyang, China, 1989; p. 191. [Google Scholar]
- Yue, P.Q. Fauna Sinica: Osteichthyes–Cypriniformes III; Science Press: Beijing, China, 2000; pp. 327–328. [Google Scholar]
- Chen, Y.X.; Luo, Q.S. Study on reproductive ecology and biology of Schizothorax kozlovi: V propagation population and reproductive behavior. J. Bijie Teach. Coll. 1997, 1, 1–5. [Google Scholar]
- Zou, X.J. Study on Karyotype and Genetic Diversity in the Population of Schizothorax (Racoma) kozlovi. Master’s Thesis, Guizhou University, Guiyang, China, 2009. [Google Scholar]
- Campana, S.E.; Neilson, J.D. Microstructure of fish otoliths. Can. J. Fish. Aquat. Sci. 1985, 42, 1014–1032. [Google Scholar] [CrossRef]
- Gao, Y.W.; Beamish, R.J. Isotopic composition of otoliths as a chemical tracer in population identification of sockeye salmon (Oncorhynchus nerka). Can. J. Fish. Aquat. Sci. 1999, 56, 2062–2068. [Google Scholar] [CrossRef]
- Bastow, T.P.; Jackson, G.; Edmonds, J.S. Elevated salinity and isotopic composition of fish otolith carbonate: Stock delineation of pink snapper, Pagrus auratus, in Shark Bay, Western Australia. Mar. Biol. 2002, 141, 801–806. [Google Scholar] [CrossRef]
- Dufour, E.; Patterson, W.P.; Höök, T.; Rutherford, E.S. Early life history of Lake Michigan alewives (Alosa pseudoharengus) inferred from intra-otolith stable isotope ratios. Can. J. Fish. Aquat. Sci. 2005, 62, 2362–2370. [Google Scholar] [CrossRef][Green Version]
- Huxham, M.; Kimani, E.; Newton, J.; Augley, J. Stable isotope records from otoliths as tracers of fish migration in a mangrove system. J. Fish. Biol. 2007, 70, 1554–1567. [Google Scholar] [CrossRef]
- Campana, S.E. Chemistry and composition of fish otoliths: Pathways, mechanisms and applications. Mar. Ecol. Prog. Ser. 1999, 188, 263–297. [Google Scholar] [CrossRef]
- Solomon, C.T.; Weber, P.K.; Cech, J.J.; Ingram, B.L.; Conrad, M.E.; Machavaram, M.V.; Pogodina, A.R.; Franklin, R.L. Experimental determination of the sources of otolith carbon and associated isotopic fractionation. Can. J. Fish. Aquat. Sci. 2006, 63, 79–89. [Google Scholar] [CrossRef]
- Gao, Y.W.; Joner, S.H.; Bargmann, G.G. Stable isotopic composition of otoliths in identification of spawning stocks of Pacific herring (Clupea pallasi) in Puget Sound. Can. J. Fish. Aquat. Sci. 2001, 58, 2113–2120. [Google Scholar] [CrossRef]
- Nelson, C.S.; Northcote, T.G.; Hendy, C.H. Potential use of oxygen and carbon isotopic composition of otoliths to identify migratory and non-migratory stocks of the New Zealand common smelt: A pilot study. N. Z. J. Mar. Freshwater Res. 1989, 23, 337–344. [Google Scholar] [CrossRef]
- Edmonds, J.S.; Fletcher, W.J. Stock discrimination of pilchards Sardinops sagax by stable isotope ratio analysis of otolith carbonate. Mar. Ecol. Prog. Ser. 1997, 152, 241–247. [Google Scholar] [CrossRef][Green Version]
- Campana, S.E. Otolith science entering the 21st century. Mar. Freshw. Res. 2005, 56, 485–495. [Google Scholar] [CrossRef]
- Gao, Y.W.; Dettman, D.L.; Piner, K.R.; Wallace, F.R. Isotopic correlation (δ18O and δ13C) of otoliths in identification of groundfish stocks. Trans. Am. Fish. Soc. 2010, 139, 491–501. [Google Scholar] [CrossRef]
- Newman, S.J.; Pember, M.B.; Rome, B.M.; Mitsopoulos, G.E.A.; Skepper, C.L.; Allsop, Q.; Saunders, T.; Ballagh, A.C.; Van Herwerden, L.; Garrett, R.N.; et al. Stock structure of blue threadfin Eleutheronema tetradactylum across northern Australia as inferred from stable isotopes in sagittal otolith carbonate. Fish. Manag. Ecol. 2011, 18, 246–257. [Google Scholar] [CrossRef]
- Shen, J.; Gao, Y. Stable isotope analyses in otoliths of silver carp: A pilot study in identification of natal sources and stock differences. Environ. Biol. Fish. 2012, 95, 445–453. [Google Scholar] [CrossRef]
- Neves, A.; Vieira, A.R.; Sequeira, V.; Paiva, R.B.; Janeiro, A.I.; Gaspar, L.M.; Gordo, L.S. Otolith shape and isotopic ratio analyses as a tool to study Spondyliosoma cantharus population structure. Mar. Environ. Res. 2019, 143, 93–100. [Google Scholar] [CrossRef]
- Avise, J.C. Phylogeography: The History and Formation of Species; Harvard University Press: Cambridge, MA, USA, 2000. [Google Scholar]
- Grant, W.S.; Cheng, W. Incorporating deep and shallow components of genetic structure into the management of Alaskan red king crab. Evol. Appl. 2012, 5, 820–837. [Google Scholar] [CrossRef]
- Guerra, Á.; Roura, Á.; González, Á.F.; Pascual, S.; Cherel, Y.; Pérez-Losada, M. Morphological and genetic evidence that Octopus vulgaris Cuvier, 1797 inhabits Amsterdam and Saint Paul Islands (southern Indian Ocean). ICES J. Mar. Sci. 2010, 67, 1401–1407. [Google Scholar] [CrossRef]
- Kang, J.H.; Park, J.Y.; Choi, T.J. Genetic differentiation of octopuses from different habitats near the Korean peninsula and eastern China based on analysis of the mDNA cytochrome C oxidase 1 gene. Genet. Mol. Res. 2012, 11, 3988–3997. [Google Scholar] [CrossRef]
- Muhammad, F.; Chen, W.; Liu, L.; Gong, L.; Xun, D.; Shafi, M.; Lü, Z. Genetic structure of Amphioctopus fangsiao (Mollusca, Cephalopoda) in Chinese waters inferred from variation in three mtDNA genes (ATPase 6, ND2, and ND5). Hydrobiologia 2019, 838, 111–119. [Google Scholar] [CrossRef]
- Yu, D.; Zhang, Z.; Zhang, J.; Lin, P.C.; Xiong, S.R.; Tang, F.L.; Liu, H.Z. Genetic diversity and population demography of Schizothorax molesworthi from the Motuo area of lower reaches of the Yarlung Zangbo River and Lohit River. Acta Hydrobiol. Sin. 2019, 43, 923–930. [Google Scholar] [CrossRef]
- Dueñas-Romero, J.J.; Granados-Amores, J.; Palacios-Salgado, D.S.; Domínguez-Contreras, J.F.; Flores-Ortega, J.R.; García-Rodríguez, F.J. Diversity and population structure of Octopus hubbsorum in the Mexican Pacific inferred from mitochondrial DNA sequences. Mar. Freshw. Res. 2020, 72, 35–43. [Google Scholar] [CrossRef]
- Xiao, W.; Zhang, Y.; Liu, H. Molecular systematics of Xenocyprinae (Teleostei: Cyprinidae): Taxonomy, biogeography, and coevolution of a special group restricted in East Asia. Mol. Phylogenet. Evol. 2001, 18, 163–173. [Google Scholar] [CrossRef]
- Ward, R.D.; Zemlak, T.S.; Innes, B.H.; Last, P.R.; Hebert, P.D.N. DNA barcoding Australia’s fish species. Phil. Trans. R. Soc. B 2005, 360, 1847–1857. [Google Scholar] [CrossRef]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral. Ecol. 2001, 26, 32–46. [Google Scholar] [CrossRef]
- Anderson, M.J.; Gorley, R.N. PERMANOVA+ for PRIMER: Guide to Software and Statistical Methods; PRIMER-E: Plymouth, UK, 2007. [Google Scholar]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef]
- Zhang, D.; Gao, F.; Jakovlić, I.; Zou, H.; Zhang, J.; Li, W.X.; Wang, G.T. PhyloSuite: An integrated and scalable desktop platform for streamlined molecular sequence data management and evolutionary phylogenetics studies. Mol. Ecol. Resour. 2020, 20, 348–355. [Google Scholar] [CrossRef]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP v6: DNA Sequence Polymorphism Analysis of Large Data Sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Bandelt, H.J.; Forster, P.; Röhl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef]
- Leigh, J.W.; Bryant, D. POPART: Full-feature software for haplotype network construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]
- Excoffier, L.; Lischer, H.E.L. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef] [PubMed]
- Corander, J.; Cheng, L.; Marttinen, P.; Sirén, J.; Tang, J. BAPS: Bayesian Analysis of Population Structure Version 6.0; Department of Mathematics and Statistics, University of Helsinki: Helsinki, Finland, 2013; p. 14. [Google Scholar]
- Grant, W.A.S.; Bowen, B.W. Shallow population histories in deep evolutionary lineages of marine fishes: Insights from sardines and anchovies and lessons for conservation. J. Hered. 1998, 89, 415–426. [Google Scholar] [CrossRef]
- Dai, Y.G.; Zou, X.J.; Xiao, H. Genetic diversity of the mtDNA D-loop in the population of Schizothorax kozlovi from the Wujiang River. Sichuan J. Zool. 2010, 29, 505–509. [Google Scholar] [CrossRef]
- Balloux, F.; Lugon-Moulin, N. The estimation of population differentiation with microsatellite markers. Mol. Ecol. 2002, 11, 155–165. [Google Scholar] [CrossRef]
- Moritz, C. Defining ‘evolutionarily significant units’ for conservation. Trends Ecol. Evol. 1994, 9, 373–375. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | Sample Year | SL (mm) | Otolith Mass (mg) | Estimated Age Range (Years) | Sampling Numbers |
|---|---|---|---|---|---|
| GT | 2017, 2019 | 278.3 (11.3) | 5.47 (0.61) | 4–6 | 23, 7 (4:3) |
| SG | 2016, 2017, 2018 | 172.0 (9.3) | 2.18 (0.27) | 1–3 | 45, 12 (5:7) |
| SJ | 2017, 2018 | 244.9 (16.6) | 3.98 (0.34) | 3–6 | 18, 12 (6:6) |
| JP | 2016 | 266.8 (6.2) | 3.45 (0.24) | 4–6 | 13, 12 (2:10) |
| Method | Source | d.f. | Sums of Squares | Mean Squares | F | p |
|---|---|---|---|---|---|---|
| ANCOVA (δ13C) | Locations | 3 | 32.635 | 10.878 | 44.182 | 0.000 |
| Otolith mass | 1 | 0.725 | 0.725 | 2.943 | 0.094 | |
| Residual | 38 | 9.356 | 0.246 | |||
| Total | 43 | 42.000 | ||||
| ANCOVA (δ18O) | Locations | 3 | 29.621 | 9.874 | 30.485 | 0.000 |
| Otolith mass | 1 | 0.137 | 0.137 | 0.423 | 0.519 | |
| Residual | 38 | 12.308 | 0.324 | |||
| Total | 43 | 42.000 | ||||
| PERMANOVA | Locations | 3 | 65,551.787 | 21,850.596 | 2425.994 | 0.001 |
| Residual | 39 | 351.268 | 9.007 | |||
| Total | 42 | 65903.055 | 1 |
| Locations | Number of Haplotypes | Haplotypes | Haplotype Diversity | Nucleotide Diversity |
|---|---|---|---|---|
| GT | 12 | Hap1-Hap12 | 0.913 (±0.040) | 0.00708 (±0.00127) |
| SG | 23 | Hap1, Hap6-Hap7, Hap11, Hap13-Hap31 | 0.932 (±0.024) | 0.00871 (±0.00124) |
| SJ | 14 | Hap1, Hap3-Hap4, Hap6, Hap9, Hap14, Hap25, Hap28-Hap29, Hap32-Hap36 | 0.967 (±0.030) | 0.00795 (±0.00081) |
| JP | 7 | Hap1, Hap6-Hap7, Hap12, Hap25, Hap28, Hap37 | 0.846 (±0.085) | 0.00361 (±0.00199) |
| Total | 37 | Hap1-Hap37 | 0.928 (±0.017) | 0.00778 (±0.00078) |
| Category | Source of Variation | d.f. | Sum of Squares | Variance Components | Percentage of Variation (%) |
|---|---|---|---|---|---|
| Location | Among locations | 3 | 38.714 | 0.23704 | 3.06 |
| Within locations | 95 | 713.862 | 7.51433 | 96.94 | |
| Total | 98 | 752.862 | 7.75137 | ||
| Lineage | Among lineages | 1 | 388.875 | 10.04411 | 72.82 |
| Within lineages | 97 | 363.701 | 3.74949 | 27.18 | |
| Total | 98 | 752.576 | 13.79360 |
| Locations | GT | SG | SJ | JP |
|---|---|---|---|---|
| GT | 0.214 | 0.398 | 0.122 | |
| SG | 0.010 | 0.126 | 0.112 | |
| SJ | −0.009 | 0.028 | 0.017 | |
| JP | 0.055 | 0.040 | 0.159 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, Y.; Gong, J.; Wu, X.; Zhu, Y.; Yang, D. Population Structure of Wild Schizothorax kozlovi in the Upper Yangtze River Based on mtDNA and Stable Isotopes, and Their Relationship with Ambient Temperature. Fishes 2022, 7, 292. https://doi.org/10.3390/fishes7050292
He Y, Gong J, Wu X, Zhu Y, Yang D. Population Structure of Wild Schizothorax kozlovi in the Upper Yangtze River Based on mtDNA and Stable Isotopes, and Their Relationship with Ambient Temperature. Fishes. 2022; 7(5):292. https://doi.org/10.3390/fishes7050292
Chicago/Turabian StyleHe, Yongfeng, Jinling Gong, Xingbing Wu, Yongjiu Zhu, and Deguo Yang. 2022. "Population Structure of Wild Schizothorax kozlovi in the Upper Yangtze River Based on mtDNA and Stable Isotopes, and Their Relationship with Ambient Temperature" Fishes 7, no. 5: 292. https://doi.org/10.3390/fishes7050292
APA StyleHe, Y., Gong, J., Wu, X., Zhu, Y., & Yang, D. (2022). Population Structure of Wild Schizothorax kozlovi in the Upper Yangtze River Based on mtDNA and Stable Isotopes, and Their Relationship with Ambient Temperature. Fishes, 7(5), 292. https://doi.org/10.3390/fishes7050292