1. Introduction
Dosidicus gigas belongs to Ommastrephidae [
1] and is a cephalopod species distributed in the eastern Pacific Ocean from the Gulf of Alaska to southern Chile [
2,
3,
4,
5].
D. gigas can vertically migrate up to 1200 m and feed in deep, middle, and surface waters [
6,
7,
8]. It is preyed upon by other cephalopods and mammals while feeding on other invertebrates and fishes [
3].
D. gigas is therefore considered a trophic carrier linking spatially relatively independent marine ecosystems and plays an important role in the marine ecosystems in the eastern Pacific Ocean [
9]. The feeding habits of marine organisms are important in the material and energy flow analysis of marine ecosystems and provide the basis for better conservation and management of fishery resources [
10]. Therefore, it is necessary to explore the feeding habits of
D. gigas in marine ecosystems.
The traditional stomach content analysis method is widely used to study the feeding ecology of marine organisms [
8,
11,
12,
13]. However, it is only applicable to a short feeding period of 24 h, and it is difficult to identify decayed components in stomachs [
14]. In marine ecosystems, the majority of polyunsaturated fatty acids in organisms come from food supply because of their limited synthetic ability [
15,
16,
17]. Therefore, fatty acid biomarkers are widely used in studies on feeding transitions and trophic relationships [
18]. Some signature fatty acids can be reflected in the fatty acid composition of high-trophic animals through feeding activities [
16]. For example, C18:2n6 and C18:3n3 are signature fatty acid markers for terrestrial organic food sources [
19], C20:4n6 indicates zoobenthos [
20], and C22:6n3 (DHA) could indicate dinoflagellates [
21]. Gong et al. [
22] investigated the dietary habits of
D. gigas using fatty acids in muscle tissue and found that the spatial variability of fatty acid profiles was mainly ascribed to the different contents of C16:0, C18:2n6, C20:4n6, and DHA in different areas. Chen et al. [
23] used fatty acid analysis to study the reproductive input strategy of female
D. gigas and found that the energy of
D. gigas during the reproductive period was mainly converted from food intake, whereas the reused muscle energy reserve was limited. Similarly, Quispe et al. [
24] suggested that the digestive gland had the highest mean proportion of fatty acids in three tissues (digestive gland, gonad, and mantle), which are obtained through food and stored in their organs as bioenergetic fuel and may then be used for the subsequent process of migration and reproduction in oceanic waters. Meanwhile, Quispe et al. [
25] investigated the variability of fatty acid profiles between the digestive gland, gonad, and mantle muscle of squids and the prey in the stomach contents and found that
D. gigas may present an energy optimization strategy during the cold season (austral winter), characterized by the intake of prey with a high energy content and rich in polyunsaturated fatty acids.
The population structure of
D. gigas is complicated [
26]. Previous studies have been conducted to distinguish the population of
D. gigas based on geographic area and body length at maturity. On the one hand, Liu et al. [
27] used inductively coupled plasma mass spectrometry (ICP-MS) to determine the trace elements in statoliths of
D. gigas sampled outside the EEZ waters of Chile, Peru, and Costa Rica. They suggested that the spatial differences in trace elements of statolith can be used to separate geographic populations of
D. gigas and found that there were at least two geographic populations in the northern and southern parts of the eastern Pacific. Sandoval-Castellanos et al. [
28] analyzed randomly amplified polymorphic DNA (RAPD) data of
D. gigas from eight eastern Pacific sites and found that a genetic structure was detected that divided the populations into northern and southern locations. On the other hand, it was suggested that sub-populations of this species can be identified based on the size of mature individuals [
29,
30,
31,
32,
33,
34]. Three groups of length-at-maturity were distinguished on the basis of the mantle length (ML) of adult males and females: a small-sized group (mantle length: 130 to 260 mm for adult males and 140 to 340 mm for adult females), a medium-sized group (mantle length: 240 to 420 mm for adult males and 280 to 600 mm for adult females), and a large-sized group (mantle length: >400 mm for adult males and >550 mm for adult females) [
35,
36]. However, the feeding habits and coexistence mechanism of the three groups are currently unclear. Therefore, we attempted to explore the variability of feeding strategies and trophic niche among these three groups using fatty acids in the muscle tissue of
D. gigas.
Here, we used fatty acid analysis to investigate the variability of feeding strategies and trophic niche among these three groups off the Peruvian exclusive economic zone. As the fatty acids of a heterotrophic organism effectively reflect those in its diet [
37], this study was designed to (1) determine the differences in fatty acid composition and trophic niche among three groups, (2) explore the variability of fatty acid composition between sexes, and (3) evaluate the feeding strategy of three groups of
D. gigas. This study provides the basis for understanding the variability of feeding strategy and coexistence mechanism among groups and sexes.
4. Discussion
Intraspecific feeding variation of pelagic predators allows them to make the best use of available food resources and improve adaptation to variable marine environments [
49]. This intraspecific variation in feeding is critical for the
Dosidicus gigas, which is widely distributed in the eastern Pacific Ocean [
36]. In the present study, we used the methodology of fatty acid analyses to evaluate the feeding strategies and coexistence mechanisms for large-, medium-, and small-sized groups of
Dosidicus gigas in Peruvian waters. We randomly selected a total of 108 specimens for the fatty acids analyses, with 39 individuals in the large-sized group, 43 individuals in the medium-sized group, and 26 individuals in the small-sized group. The sample size was comparable to that of previous studies using fatty acids as trophic biomarkers (e.g., 18 individuals of
Idioteuthis cordiformis used by Jackson et al. [
50]; 32 individuals of
D. gigas used by Quispe-Machaca et al. [
25]).
In this study, 28 fatty acids were detected in the samples of squids off the Peruvian EEZ, and the fatty acid composition was dominated by PUFAs (50.75–51.64%), followed by SFAs (34.72–36.73%) and MUFAs (11.64–14.53%). Our results were consistent with the findings of Gong et al. [
22] and Chen et al. [
23]. Moreover, a similar relative fatty acid composition in other cephalopod species was also reported. Lin et al. [
51] analyzed the fatty acid composition of the muscles, digestive glands, and ovaries of
D. gigas. They found that the fatty acid composition in all three tissues was dominated by PUFAs, and the relative content of MUFAs was the lowest. In addition, the fatty acid composition in Octopodidae and Sepiidae was also dominated by PUFAs, SFAs, and MUFAs, which had the lowest content [
52,
53]. It could be seen that the proportions of PUFAs, MUFAs, and SFAs in cephalopod muscle tissue were generally similar.
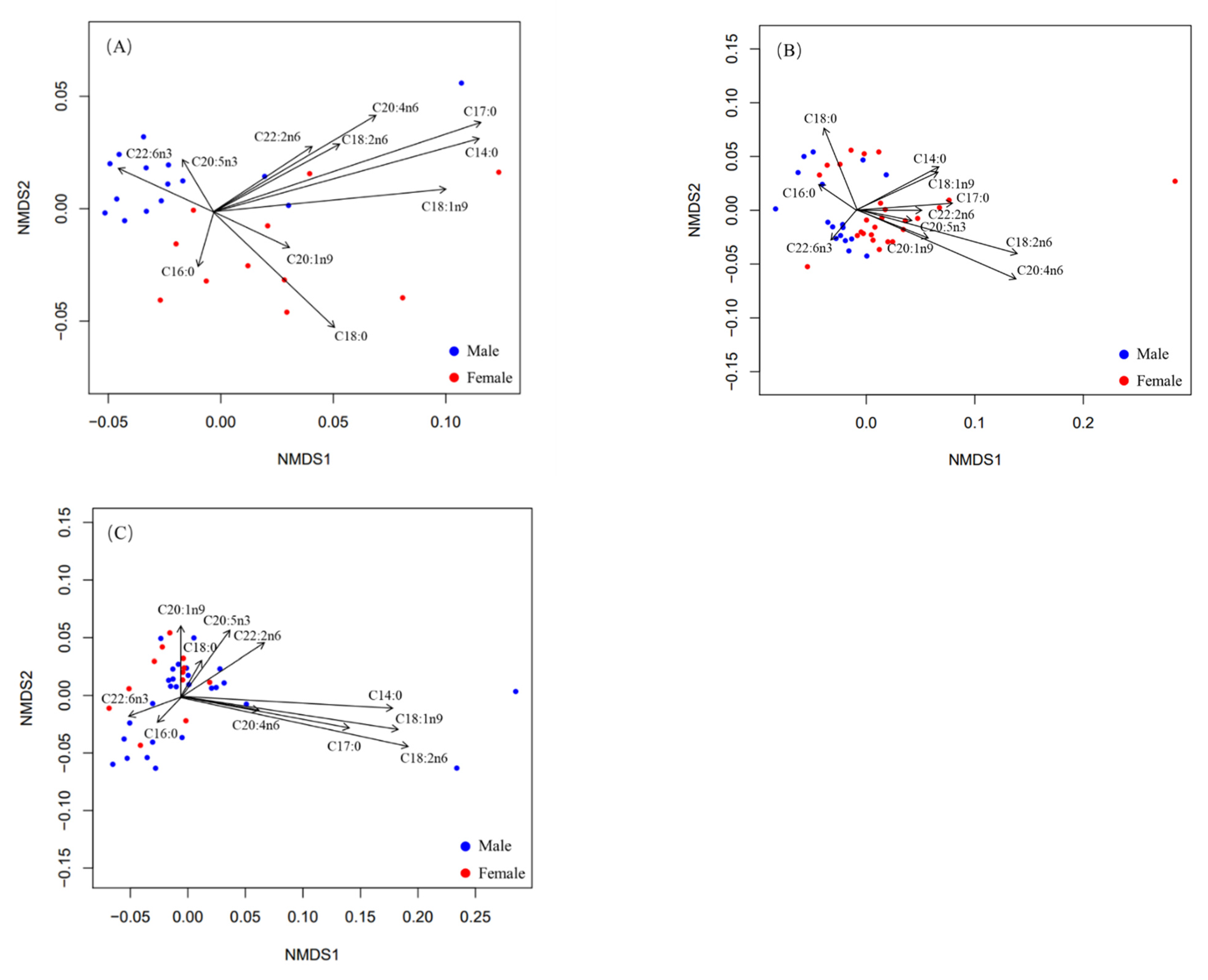
4.1. Intraspecific Sexual Variation
In marine ecosystems, male and female individuals of many species coexist by using different habitats or adopting different feeding behaviors [
54]. This coexistence strategy is often associated with sexual variability in size, feeding behavior, and nutrient requirements [
55].
In this study, we found that the fatty acid composition in female and male samples from the small-sized group differed significantly, and C18:1n9, C20:5n3, and C22:6n3 were identified as the main fatty acids, which were responsible for the above differences. Diet species may differ between male and female individuals in small groups. In the medium-sized group, the fatty acid composition also showed significant differences between males and females. Given that females in small- and medium-sized groups are slightly larger in body length than males, sexual differences in body size may have an impact on the feeding of squid, which in turn may lead to the differences in fatty acid composition. High levels of C20:4n6 suggested that females tend to consume more benthic organisms, making them frequently inhabit deeper waters. Therefore, their coexistence also benefits from sexual difference in habitats. However, the large-sized group showed no significant differences between males and females. It has been reported that individuals with large body length tend to consume more food to maintain their metabolism [
56]. Meanwhile, Liu et al. [
18] pointed out that, as body length increased,
D. gigas tended to adopt a specific feeding strategy. This feeding strategy may lead to the similarity in diet of males and females, which in turn affects the fatty acids composition.
Rossi et al. [
39] suggested that the higher DHA/EPA ratio corresponded to the higher trophic level. The DHA/EPA ratio in males was higher than in females in the three groups, so males likely fed on prey with higher trophic levels. However, Gong et al. [
54] explored immature
D. gigas through stable isotope analysis and found that the trophic level of females was higher than that of males and that females tended to feed on prey with a higher trophic level. Since all samples in our study were sexually mature individuals, DHA and EPA contents were strongly influenced by reproductive activity [
20,
57]. Therefore, a possible diet transition from the immature stage to the mature stage may exist in males and females and may be affected by reproductive activities. However, regardless of the changes in the contents of DHA and EPA in males and females during growth, the trophic levels of prey were different among different growth stages. Consequently, they can coexist in the same waters.
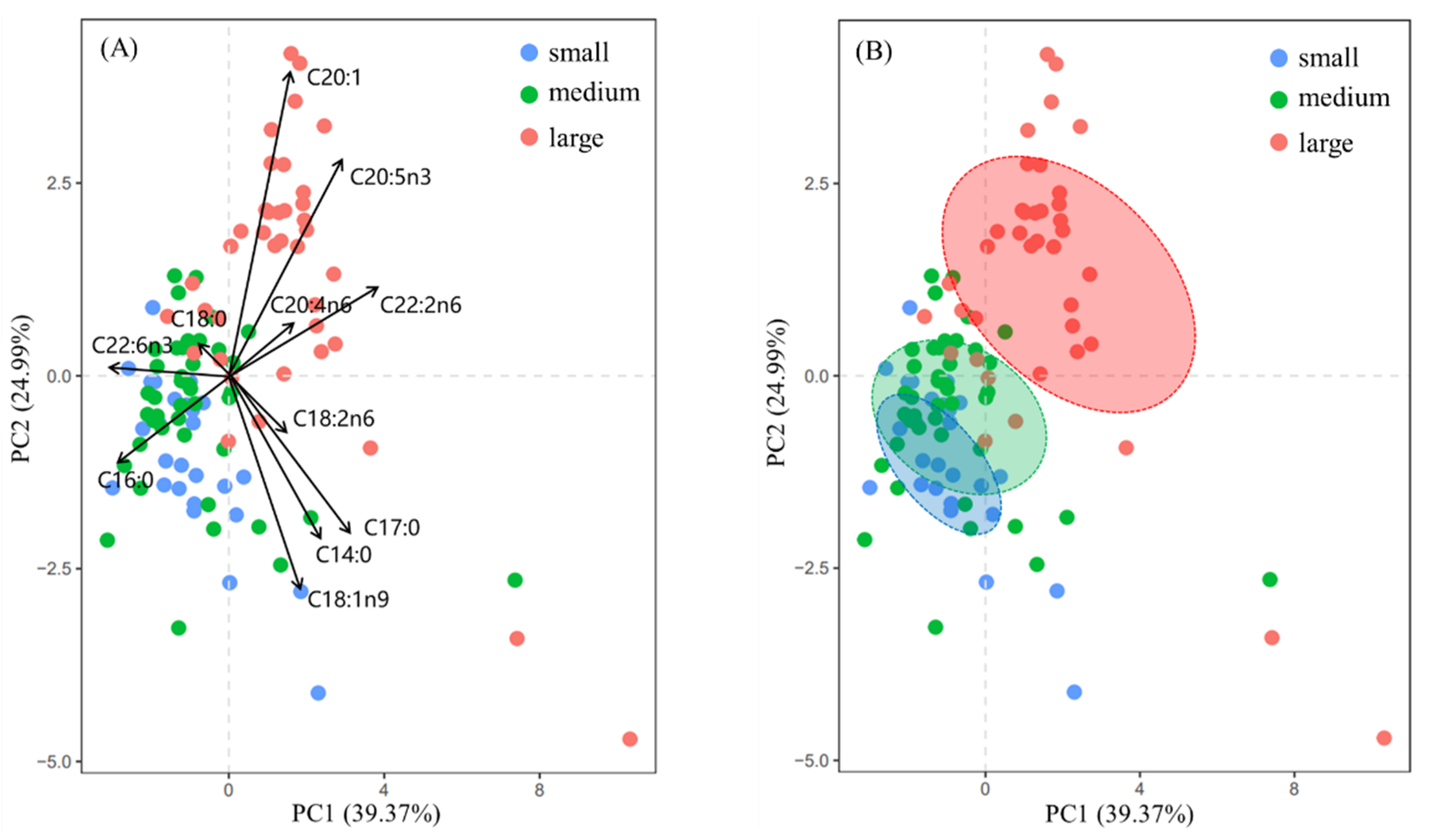
4.2. Interspecific Variation
PCA results showed that C16:0, C18:1n9, C20:1n9, C20:5n3, and C22:6n3 were the main fatty acids that were responsible for the differences in fatty acid composition among the small-, medium- and large-sized groups. The content of C16:0 in the large-sized group was significantly lower than that in the small- and medium-sized groups (
p < 0.05). Marine fatty acids come from their own synthesis or food supply, and cephalopods can extend and desaturate the carbon chain of C16:0 to synthesize C18:1n9 [
58]. However, the content of C18:1n9 showed no significant difference among small-, medium-, and large-sized groups (
p > 0.05), so most of C18:1n9 in the muscle tissue of the large-sized group probably originated from feeding activities.
Since the fatty acid composition of organisms largely depends on the fatty acid profile of their food [
59] and signature fatty acids indicate feeding habits, fatty acid composition and content can be used to indicate food sources. According to the available fatty acid marker system (
Table 2), C18:1n9 in marine organisms is mostly from zooplankton [
39], and therefore, the large-sized group of
D. gigas may feed on more zooplankton or other species that feed on zooplankton. In addition, the content of C20:1n9 in the muscle tissue in the large-sized group was significantly higher than that in the small- and medium-sized groups, suggesting that more copepods or other species that feed on copepods may be consumed by the large-sized group [
60,
61,
62,
63]. Cecilia et al. [
63] found that
Trachurus murphyi significantly exceeded other species in the stomach of
D. gigas in terms of frequency, quantity, and weight. Given that the main prey of
Trachurus murphyi includes copepods and zooplankton [
60,
61], we inferred that C18:1n9 and C20:1n9 in
D. gigas in our study may be derived from
Trachurus murphyi, and the large-sized population had a more significant feeding preference for
Trachurus murphyi. Similarly, the content of C20:4n6 in the muscle tissue of the large-sized group was also significantly higher than that in small- and medium-sized groups, indicating that more benthic organisms may be preyed on by the large-sized group. The oxygen consumption of squids in normal metabolism is negatively correlated with body length, and the larger body size of squids corresponds to lower oxygen consumption [
56]. As a result, the large-sized group can remain at greater depths for longer periods of time due to relatively low oxygen consumption, thereby feeding on prey that lives in deeper water.
In our study, the trophic niches of the small- and medium-sized groups overlapped highly, which may reveal a similarity in their use of food or habitat resources. In contrast, the standard ellipse of the large-sized group was clearly distinguished from that of the small- and medium-sized groups (
Figure 3). Tentacles and beaks, as the feeding apparatus, influence the feeding and resource utilization of
D. gigas [
49]. Firstly, the longer tentacles and larger beaks of large-sized groups allow them to feed on larger prey, thus broadening their trophic niche. Secondly, large-sized groups have larger feeding organs and feed on larger prey with a higher trophic level, so that stable food supplements can be obtained by small- and medium-sized groups. Thirdly, the contents of C18:2n6 and C18:3n3 indicate the terrestrial organic source of marine organisms, such as estuarine/coastal macroalgae [
19,
43]. In our study, the sum of the C18:2n6 and C18:3n3 in the large-sized group was slightly higher than in other groups, suggesting frequent feeding nearshore where the terrestrial input exists. The results were in agreement with the point of Gong et al. [
49]. According to the report by Gong et al. [
49], both pelagic and nearshore food resources were utilized by
D. gigas. Therefore, compared with the small- and medium-sized groups, the large-sized group may migrate to the sea area closer to the coast for feeding. In other words, the differences in food sources and habitats may contribute to the stable survival of different groups in the same waters. In addition, the relative oxygen consumption of
D. gigas is negatively correlated with body length [
56], so larger individuals can migrate to deeper waters to feed. In our study, the content of C20:4n6 indicating the food origin of zoobenthos was highest in the large group, followed by the medium group and lowest in the small group. The large-sized group can migrate more easily to deeper locations. As a result, both the horizontal and vertical migration areas of the large-sized groups were larger than those of the medium- and small-sized groups, thus occupying the largest area of trophic niches. However, the ratio of DHA/EPA indicating trophic level was highest in the small-sized group and lowest in the large-sized group. Previous studies indicated that DHA and EPA, which are essential for the structure and function of biological membranes, may be significantly altered by factors unrelated to feeding (reproductive activities) [
57,
64]. The samples in this study were all sexually mature individuals, suggesting that reproductive activities may lead to abnormal changes in DHA/EPA among different groups.
5. Conclusions
This study focused on a comparison of fatty acids in the muscle tissue of small-, medium-, and large-sized groups of D. gigas off Peru and further explored the feeding habit and trophic niche of these three groups. There was no significant difference in fatty acid composition between the small and medium groups, and their trophic niche overlapped significantly. Meanwhile, they usually achieve coexistence through internal regulation, in which males likely feed on prey with higher trophic levels. We found that there was no significant difference in fatty acid composition between males and females in large groups. However, the fatty acid composition differed significantly between the large-sized group and other groups. The squids from the large-sized group may feed more frequently in nearshore and deep waters. In other words, they may coexist with other groups through a more specialized feeding strategy and a wider feeding space. However, the samples selected in this study were sexually mature individuals, and discriminant equations of hard tissue, such as beak, could be established for the three size groups. Once three size groups are distinguished by discriminant equations, it is possible to study early life history samples. In addition, multiple analysis may benefit from correction, especially in cases with a great deal of testing. Finally, there are some limitations of fatty acid analysis. Fatty acid analysis, for example, is only an indirect indication of food source and does not determine trophic level. In future studies, stable isotopes and gastric DNA barcoding technology can be combined to improve the feeding ecology of the three populations from multiple dimensions.


 ,
, 




{kind=link}
{kind=link}
{kind=link}