
Oral Administration of Probiotics (Bacillus subtilis and Lactobacillus plantarum) in Nile Tilapia (Oreochromis niloticus) Vaccinated and Challenged with Streptococcus agalactiae

 , , , , , ,
, , , , , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Inoculum of Streptococcus Agalactiae
2.2. Vaccine Preparation
2.3. In Vivo Infection Assay
2.3.1. Control and Experimental Diets
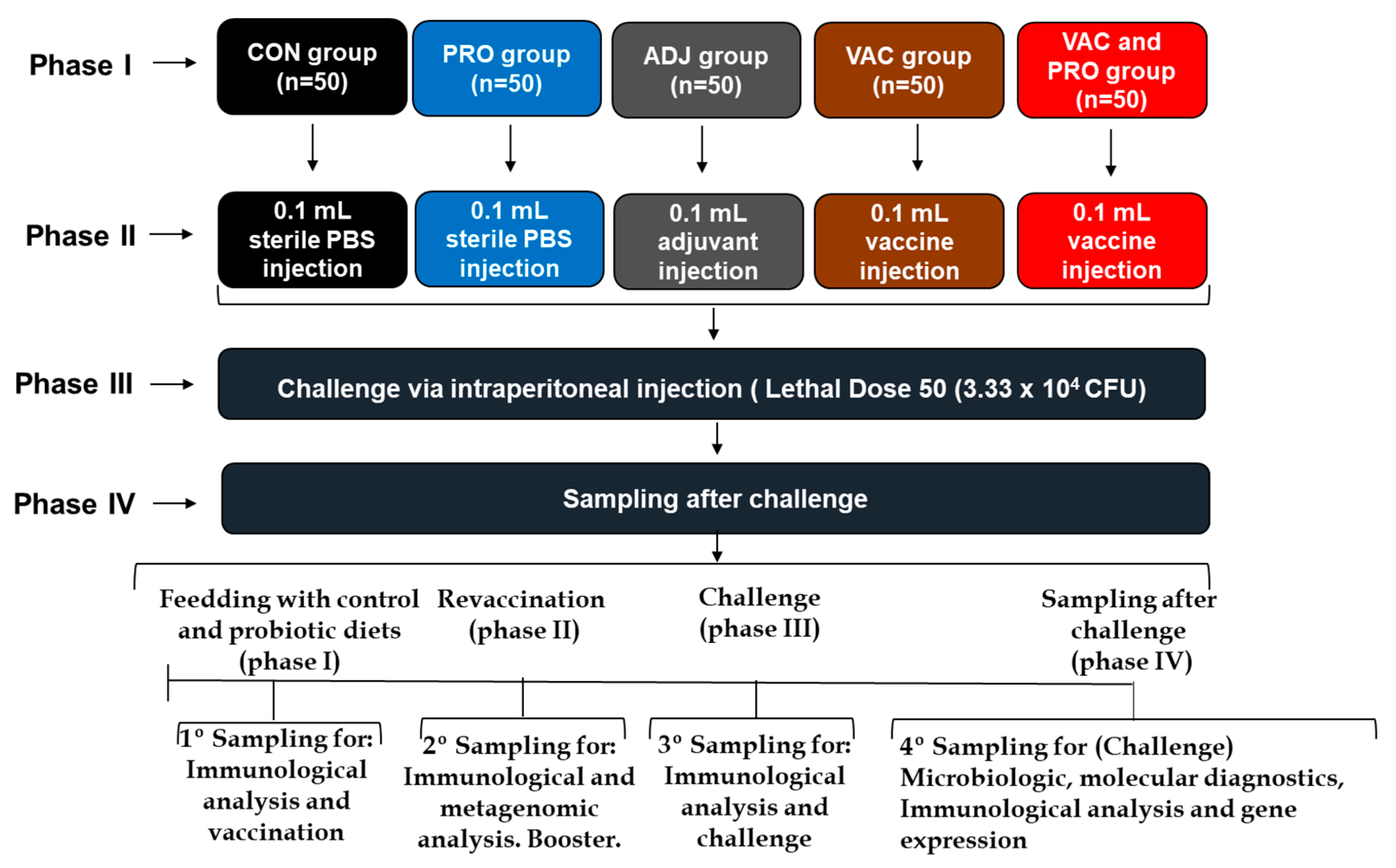
2.3.2. Experimental Design
2.4. Microbiological and Molecular Diagnostics
2.5. Biochemical Analysis
2.5.1. Lysozyme Activity
2.5.2. Phagocyte Respiratory Burst Activity
2.5.3. Assessment of the Specific S. agalactiae Antibody Response (IgM)
2.5.4. RNA Isolation, cDNA Synthesis, and Quantitative Reverse-Transcription PCR (RTq-PCR)
2.5.5. DNA Extraction, PCR Amplification and 16S rRNA Sequencing
2.6. Statistical Analysis
3. Results
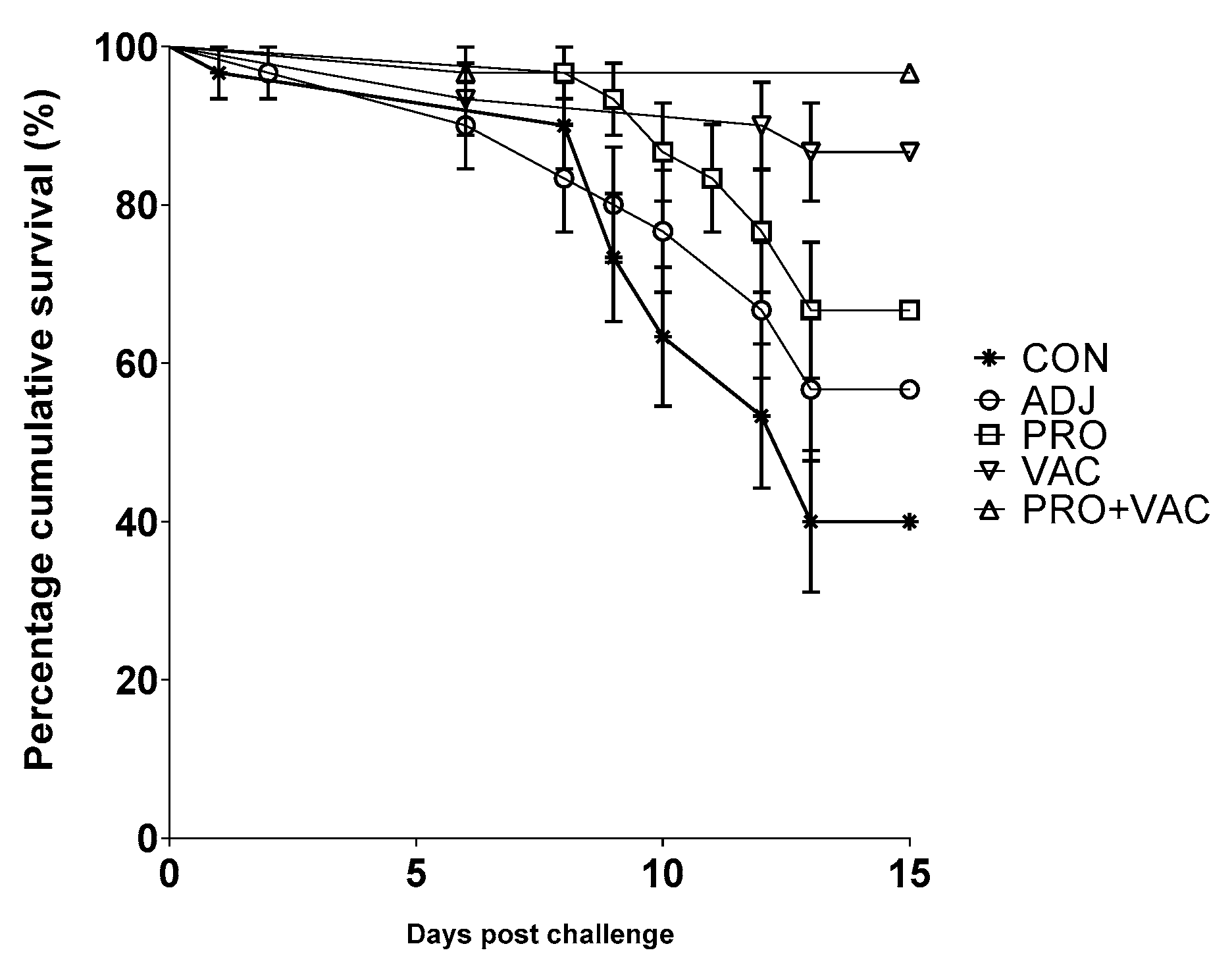
3.1. Vaccine Efficacy
3.2. S. agalactiae Isolation after Infection Challenge Test
3.3. Phagocyte Respiratory Burst and Lysozyme Activities
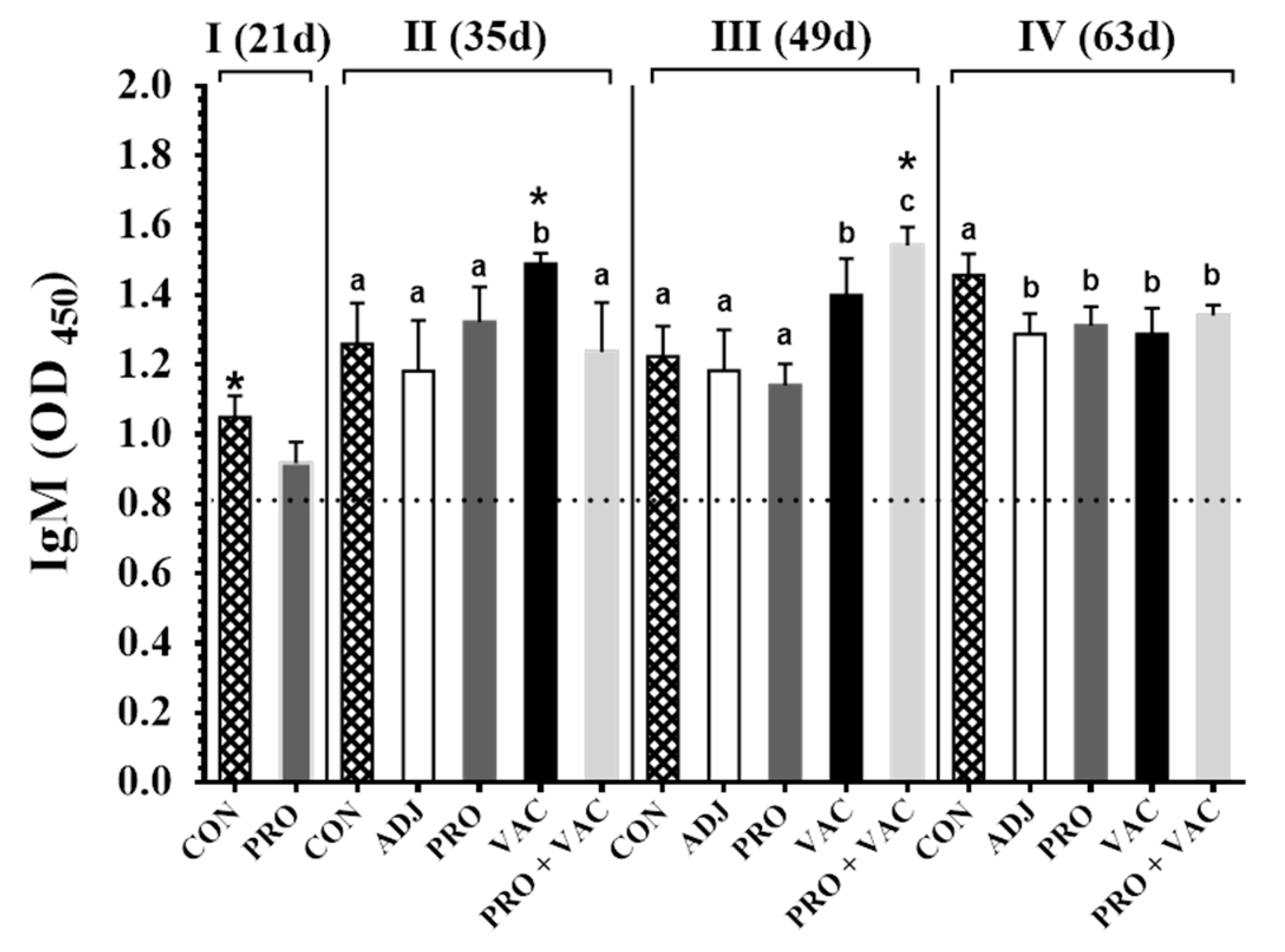
3.4. Quantification of IgM Produced after Probiotic Diet Feeding, Vaccination and Infection Challenge Test
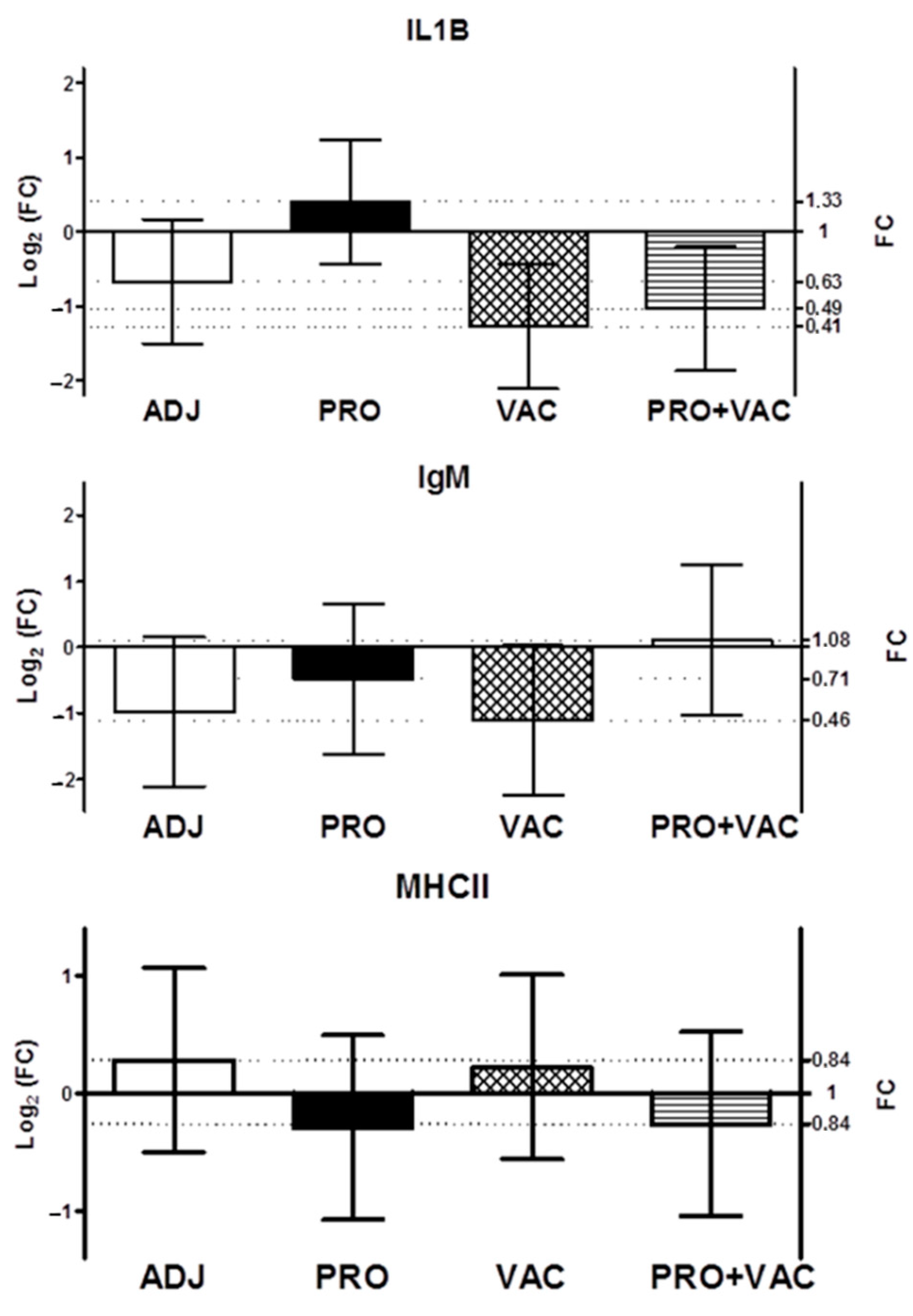
3.5. Gene Expression
3.5.1. Metagenomic Analyses
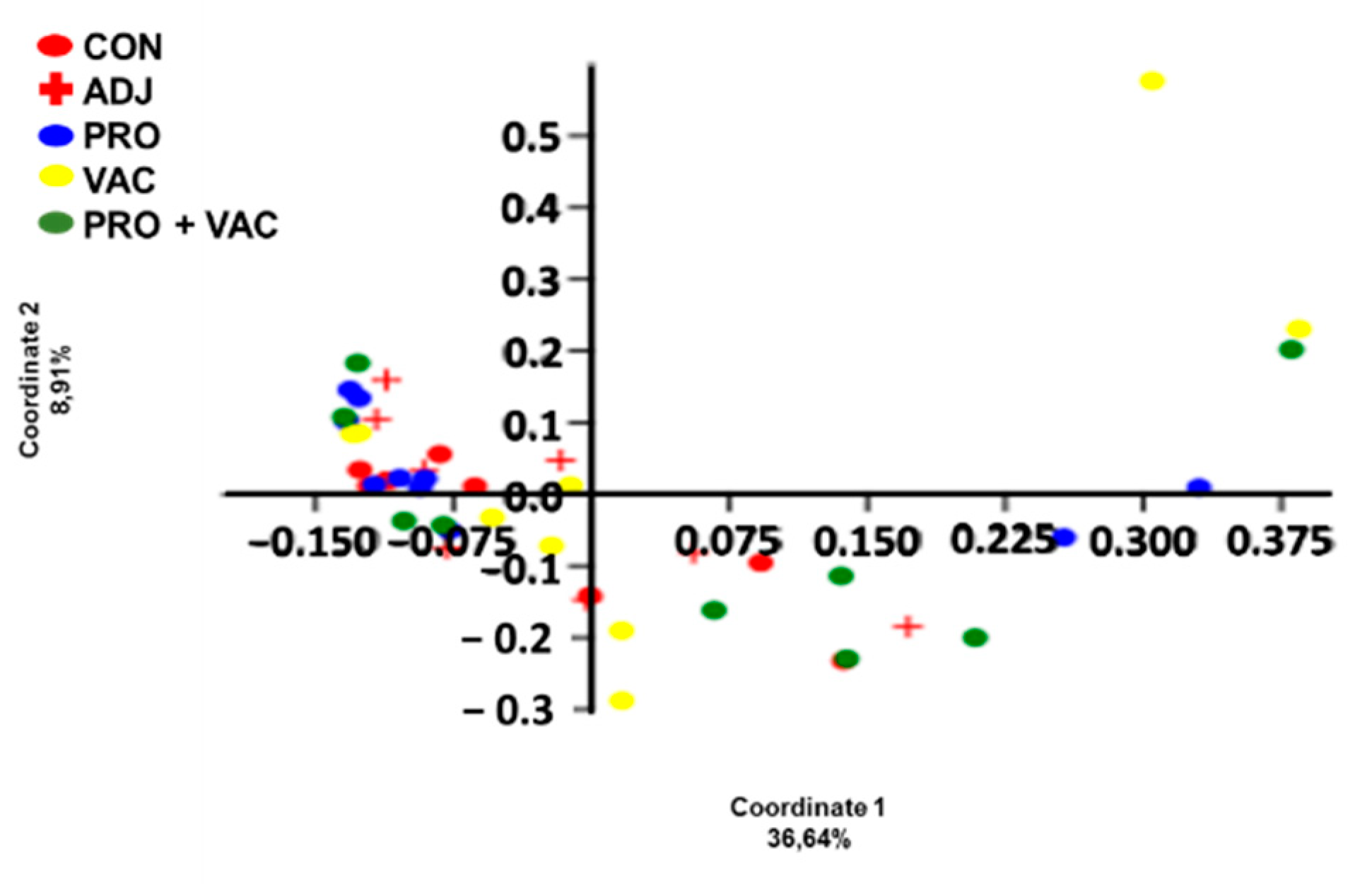
3.5.2. Alpha and Beta Diversity
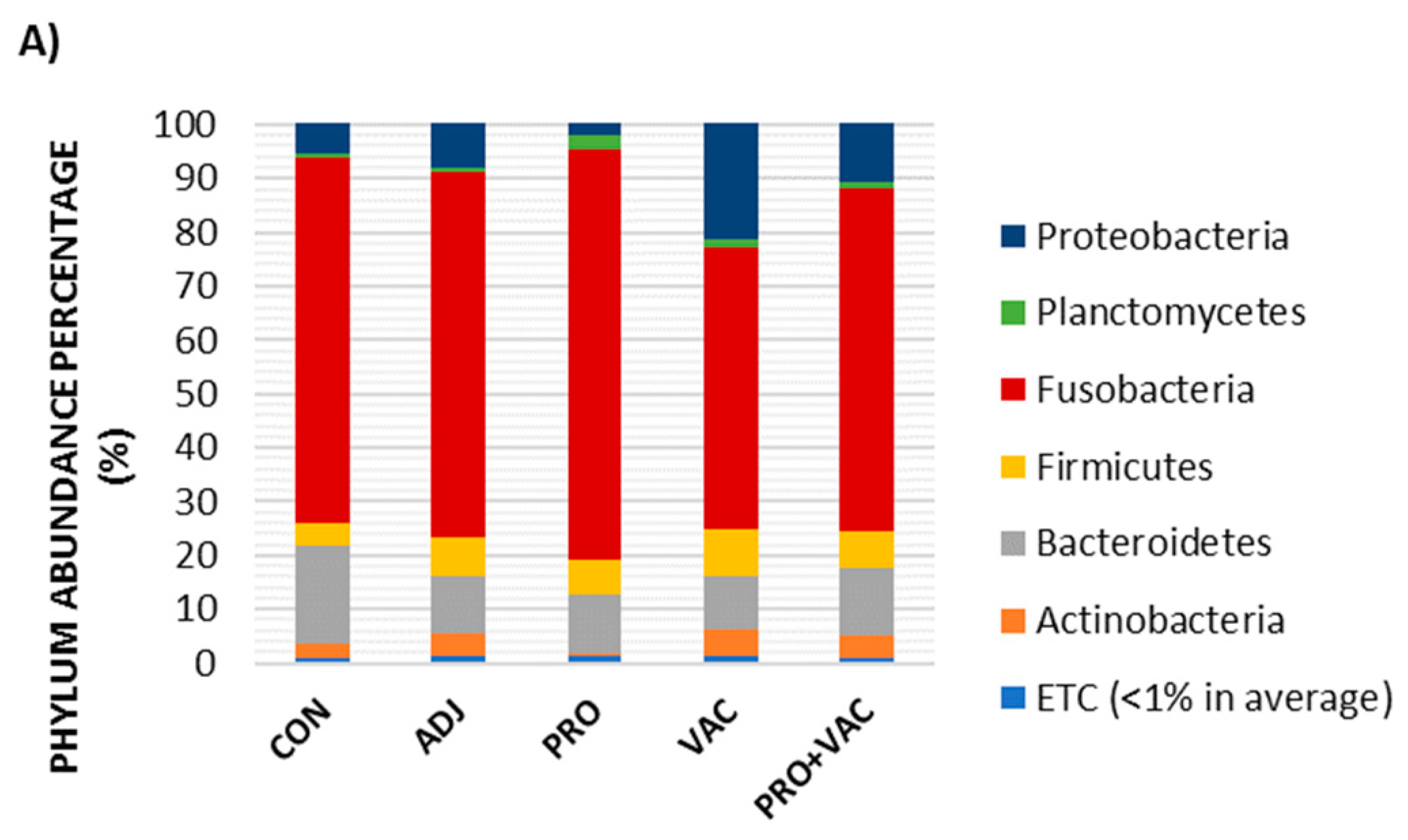
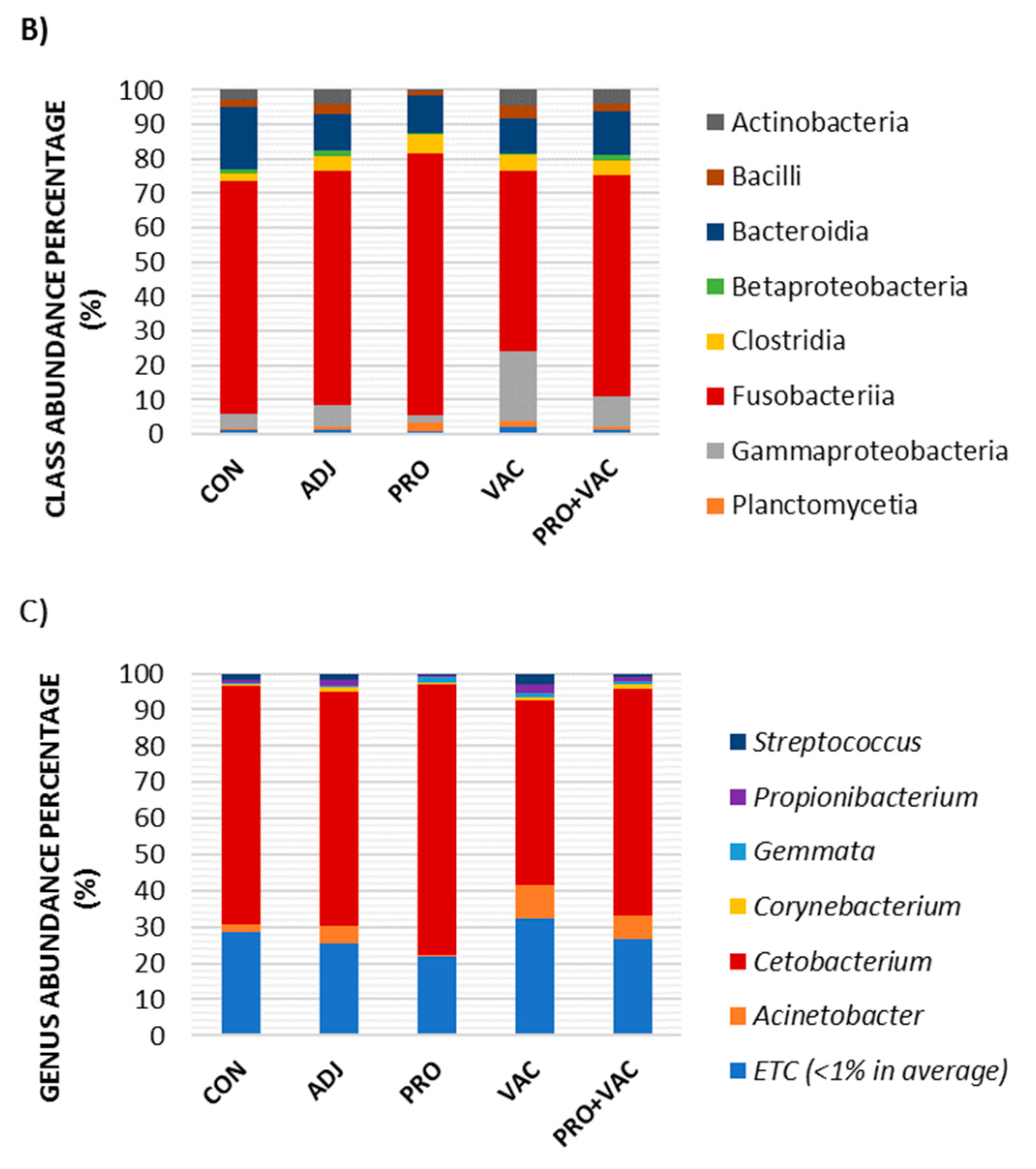
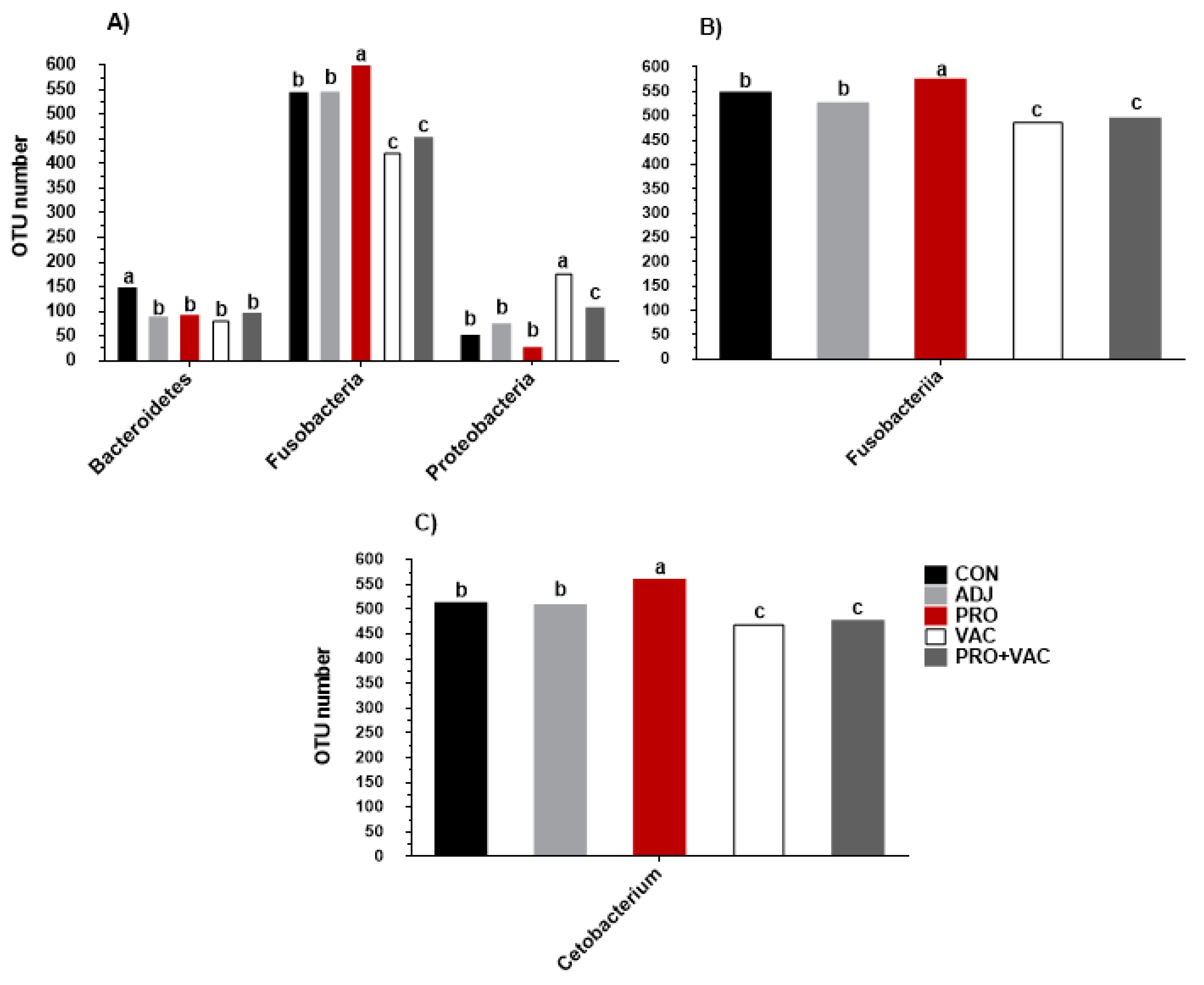
3.5.3. Gut Microbiological Profiles
4. Discussion
4.1. Experimental Infection with S. agalactiae
4.2. Relationships among the Gut Microbiological Profiles of Different Groups
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nadal, A.L.; Ikeda-Ohtsubo, W.; Sipkema, D.; Peggs, D.; McGurk, C.; Forlenza, M.; Wiegertjes, G.F.; Brugman, S. Feed, Microbiota, and Gut Immunity: Using the Zebrafish Model to Understand Fish Health. Front. Immunol. 2020, 11, 114. [Google Scholar] [CrossRef] [PubMed]
- Anuário Peixe BR da Piscicultura (Peixe BR Yearbook of Fish Farming); Fish Brazilian Association: Peixe, Brazil, 2020; pp. 1–136. Available online: https://www.peixebr.com.br/anuario2022/ (accessed on 17 July 2022).
- Brudeseth, B.E.; Wiulsrød, R.; Fredriksen, B.N.; Lindmo, K.; Løkling, K.E.; Bordevik, M.; Steine, N.; Klevan, A.; Gravningen, K. Status and future perspectives of vaccines for industrialised fin-fish farming. Fish Shellfish. Immunol. 2013, 35, 1759–1768. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.Y.; Chen, D.D.; Cui, Z.W.; Zhang, X.Y.; Zhou, Y.Y.; Guo, X.; Li, A.H.; Zhang, Y.A. Oral vaccination of tilapia against Streptococcus agalactiae using Bacillus subtilis spores expressing Sip. Fish Shellfish. Immunol. 2019, 86, 999–1008. [Google Scholar] [CrossRef] [PubMed]
- Iregui, C.A.; Comas, J.; Vásquez, G.M.; Verján, N. Experimental early pathogenesis of Streptococcus agalactiae infection in red tilapia Oreochromis spp. J. Fish Dis. 2016, 39, 205–215. [Google Scholar] [CrossRef]
- Caruso, G. Antibiotic Resistance in Fish Farming Environments: A Global Concern. J. FisheriesSciences.com 2016, 10, 9–13. [Google Scholar]
- Liu, G.; Zhu, J.; Chen, K.; Gao, T.; Yao, H.; Liu, Y.; Zhang, W.; Lu, C. Development of Streptococcus agalactiae vaccines for tilapia. Dis. Aquat. Org. 2016, 122, 163–170. [Google Scholar] [CrossRef]
- Merrifield, D.L.; Dimitroglou, A.; Foey, A.; Davies, S.J.; Baker, R.T.M.; Bøgwald, J.; Castex, M.; Ringø, E. The current status and future focus of probiotic and prebiotic applications for salmonids. Aquaculture 2010, 302, 1–18. [Google Scholar] [CrossRef]
- Hai, N.v. Research findings from the use of probiotics in tilapia aquaculture: A review. Fish Shellfish. Immunol. 2015, 45, 592–597. [Google Scholar] [CrossRef]
- Kuebutornye, F.K.A.; Abarike, E.D.; Sakyi, M.E.; Lu, Y.; Wang, Z. Modulation of nutrient utilization, growth, and immunity of Nile tilapia, Oreochromis niloticus: The role of probiotics. Aquac. Int. 2020, 28, 277–291. [Google Scholar] [CrossRef]
- Guimarães, M.C.; Guimarães, A.I.C.d.; Natori, M.M.; Alarcon, M.F.F.; Dias, D.d.; Ishikawa, C.M.; Tapia-Paniagua, S.; Moriñigo, M.Á.; Moyano, F.J.; Tachibana, L. Oral administration of Bacillus subtilis and Lactobacillus plantarum modulates the gut microbiota and increases the amylase activity of Nile tilapia (Oreochromis niloticus). Aquac. Int. 2020, 29, 91–104. [Google Scholar] [CrossRef]
- Secombes, C.J.; Belmonte, R. Overview of the Fish Adaptive Immune System. In Fish Vaccines; Springer: Basel, Switzerland, 2016; pp. 35–52. [Google Scholar] [CrossRef]
- Derome, N.; Gauthier, J.; Boutin, S.; Llewellyn, M. Bacterial Opportunistic Pathogens of Fish. In The Rasputin Effect: When Commensals and Symbionts Become Parasitic, Advances in Environmental Microbiology; Hurst, C.J., Ed.; Springer International Publishing: Cham, Switzerland, 2016; pp. 81–108. [Google Scholar] [CrossRef]
- Dadar, M.; Dhama, K.; Vakharia, V.N.; Hoseinifar, S.H.; Karthik, K.; Tiwari, R.; Khandia, R.; Munjal, A.; Salgado-Miranda, C.; Joshi, S.K. Advances in Aquaculture Vaccines Against Fish Pathogens: Global Status and Current Trends. Rev. Fish. Sci. Aquac. 2017, 25, 184–217. [Google Scholar] [CrossRef]
- Assefa, A.; Abunna, F. Maintenance of Fish Health in Aquaculture: Review of Epidemiological Approaches for Prevention and Control of Infectious Disease of Fish. Vet. Med. Int. 2018, 2018, 1–10. [Google Scholar] [CrossRef]
- Lazado, C.C.; Marlowe, C.; Caipang, A. Immunology Mucosal immunity and probiotics in fish. Fish Shellfish. Immunol. 2014, 39, 78–89. [Google Scholar] [CrossRef]
- Berridge, B.R.; Bercovier, H.; Frelier, P.F. Streptococcus agalactiae and Streptococcus difficile 16S-23S intergenic rDNA: Genetic homogeneity and species-specific PCR. Vet. Microbiol. 2001, 78, 165–173. [Google Scholar] [CrossRef]
- Mansor, N.N.; Nawi, M.F.; Yusoff, S.; Abdullah, S.; Saad, M. Determination of LD 50 for Streptococcus agalactiae and Staphylococcus aureus infections in tilapia. J. Vet. Malaysia 2011, 23, 22–27. [Google Scholar]
- Salvador, R.; Claudiano, G.d.; Loureiro, B.A.; Marcusso, P.F.; Eto, S.F.; Pilarski, F.; Toazza, C.S.; de Moraes, J.R.E.; de Moraes, F.R. Desempenho e hematologia de tilápias-do-nilo alimentadas com Saccharomyces cerevisiae e vacinadas contra Streptococcus agalactiae. Pesqui. Agropecu. Bras. 2013, 48, 892–898. [Google Scholar] [CrossRef]
- NRC. Nutrient Requirements of Fish and Shrimp; The National Academies Press: Washington, DC, USA, 2011. [Google Scholar] [CrossRef]
- Dias, D.C.; Leonardo, A.F.G.; Tachibana, L.; Corrêa, C.F.; Bordon, I.C.A.C.; Romagosa, E.; Ranzani-Paiva, M.J.T. Effect of incorporating probiotics into the diet of matrinxã (Brycon amazonicus) breeders. J. Appl. Ichthyol. 2012, 28, 40–45. [Google Scholar] [CrossRef]
- Tachibana, L.; Telli, G.S.; Dias, D.d.; Gonçalves, G.S.; Ishikawa, C.M.; Cavalcante, R.B.; Natori, M.M.; Hamed, S.b.; Ranzani-Paiva, M.J.T. Effect of feeding strategy of probiotic Enterococcus faecium on growth performance, hematologic, biochemical parameters and non-specific immune response of Nile tilapia. Aquac. Rep. 2020, 16, 100277. [Google Scholar] [CrossRef]
- Amend, D.F. Potency testing of fish vaccines. Dev. Biol. Stand 1980, 49, 447–454. [Google Scholar]
- Sebastião, F.d.; Lemos, E.G.M.; Pilarski, F. Validation of absolute quantitative real-time PCR for the diagnosis of Streptococcus agalactiae in fish. J. Microbiol. Methods 2015, 119, 168–175. [Google Scholar] [CrossRef]
- Kim, D.H.; Austin, B. Innate immune responses in rainbow trout (Oncorhynchus mykiss, Walbaum) induced by probiotics. Fish Shellfish Immunol. 2006, 21, 513–524. [Google Scholar] [CrossRef]
- Lie, Ø.; Evensen, Ø.; Sørensen, A.; Frøysadal, E. Study on lysozyme activity in some fish species. Dis. Aquat. Org. 1989, 6, 1–5. [Google Scholar] [CrossRef]
- Aly, S.M.; Mohamed, M.F.; John, G. Effect of probiotics on the survival, growth and challenge infection in Tilapia nilotica (Oreochromis niloticus). Aquac. Res. 2008, 39, 647–656. [Google Scholar] [CrossRef]
- Clissa, P.B.; Orefice, D.P.; Colombini, M.; Guimarães, M.C.; Bach, E.E.; Dias, D.C.; Tachibana, L. Production of polyclonal antibodies anti-igm of nile tilapia, Oreochromis niloticus for standardization of immunoenzymatic assay. Ars Vet. 2021, 37, 3–4. [Google Scholar] [CrossRef]
- Liu, M.; Pan, J.; Ji, H.; Zhao, B.; Zhang, S. Vitellogenin mediates phagocytosis through interaction with FcγR. Mol. Immunol. 2011, 49, 211–218. [Google Scholar] [CrossRef]
- Wang, E.; Wang, K.; Chen, D.; Wang, J.; He, Y.; Long, B.; Yang, L.; Yang, Q.; Geng, Y.; Huang, X.; et al. Evaluation and selection of appropriate reference genes for real-time quantitative PCR analysis of gene expression in Nile tilapia (Oreochromis niloticus) during vaccination and infection. Int. J. Mol. Sci. 2015, 16, 9998–10015. [Google Scholar] [CrossRef]
- Pang, J.c.; Gao, F.y.; Lu, M.x.; Ye, X.; Zhu, H.p.; Ke, X.l. Major histocompatibility complex class IIA and IIB genes of Nile tilapia Oreochromis niloticus: Genomic structure, molecular polymorphism and expression patterns. Fish Shellfish. Immunol. 2013, 34, 486–496. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 408, 402–408. [Google Scholar] [CrossRef]
- Martínez, G.; Shaw, E.M.; Carrillo, M.; Zanuy, S. Protein salting-out method applied to genomic DNA isolation from fish whole blood. Biotechniques 1998, 24, 238–239. [Google Scholar] [CrossRef]
- Tapia-Paniagua, S.T.; Chabrillón, M.; Díaz-Rosales, P.; de la Banda, I.G.; Lobo, C.; Balebona, M.C.; Moriñigo, M.A. Intestinal Microbiota Diversity of the Flat Fish Solea senegalensis (Kaup, 1858) Following Probiotic Administration. Microb. Ecol. 2010, 60, 310–319. [Google Scholar] [CrossRef]
- Mc Murdie, P.J.; Holmes, S. Phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef]
- Tapia-Paniagua, S.T.; Fumanal, M.; Anguís, V.; Fernández-DÍaz, C.; Alarcón, F.J.; Moriñigo, M.A.; Balebona, M.C. Modulation of intestinal microbiota in Solea senegalensis fed low dietary level of Ulva ohnoi. Front. Microbiol. 2019, 10, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, E.L.; Meier, P. Nonparametric Estimation from Incomplete Observations. J. Am. Stat. Assoc. 1958, 53, 457–481. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. Past: Paleontological Statistics Software Package For Education And Data Analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Henriksson, P.J.G.; Rico, A.; Troell, M.; Klinger, D.H.; Buschmann, A.H.; Saksida, S.; Chadag, M.v.; Zhang, W. Unpacking factors influencing antimicrobial use in global aquaculture and their implication for management: A review from a systems perspective. Sustain. Sci. 2018, 13, 1105–1120. [Google Scholar] [CrossRef] [PubMed]
- Valenti, W.C.; Barros, H.P.; Moraes-Valenti, P.; Bueno, G.W.; Cavalli, R.O. Aquaculture in Brazil: Past, present and future. Aquac. Rep. 2021, 19, 100611. [Google Scholar] [CrossRef]
- Zhang, D.; Gao, Y.; Li, Q.; Ke, X.; Liu, Z.; Lu, M.; Shi, C. An effective live attenuated vaccine against Streptococcus agalactiae infection in farmed Nile tilapia (Oreochromis niloticus): Live attenuated vaccine for Nile tilapia. Fish Shellfish. Immunol. 2020, 98, 853–859. [Google Scholar] [CrossRef]
- Uribe, C.; Folch, H.; Enriquez, R.; Moran, G. Innate and adaptive immunity in teleost fish: A review. Veterinární Med. 2011, 56, 486–503. [Google Scholar] [CrossRef]
- Aly, S.M.; al Zohairy, M.A.; Rahmani, A.H.; Fathi, M.; Atti, N.M.A. Trials to improve the response of Oreochromis niloticus to Aeromonas hydrophila vaccine using immunostimulants (garlic, Echinacea) and probiotics. Afr. J. Biotechnol. 2016, 15, 989–994. [Google Scholar] [CrossRef]
- Venkatalakshmi, S.; Ebanasar, J. Immunostimulatory effect of Lactobacillus sporogenes on the nonspecific defense mechanisms of Oreochromis mossambicus (Peters). Int. J. Fish. Aquat. Stud. 2015, 2, 362–369. [Google Scholar]
- Pérez-Sánchez, T.; Balcázar, J.L.; Merrifield, D.L.; Carnevali, O.; Gioacchini, G.; de Blas, I.; Ruiz-Zarzuela, I. Expression of immune-related genes in rainbow trout (Oncorhynchus mykiss) induced by probiotic bacteria during Lactococcus garvieae infection. Fish Shellfish. Immunol. 2011, 31, 196–201. [Google Scholar] [CrossRef]
- Shahin, K.; Shinn, A.P.; Metselaar, M.; Ramirez-Paredes, J.G.; Monaghan, S.J.; Thompson, K.D.; Hoare, R.; Adams, A. Efficacy of an Inactivated Whole-Cell Injection Vaccine for Nile Tilapia, Oreochromis niloticus (L.), against Multiple Isolates of Francisella noatunensis Subsp. orientalis from Diverse Geographical Regions; Elsevier Ltd.: Amsterdam, The Netherlands, 2019. [Google Scholar] [CrossRef]
- Marcusso, P.F.; Salvador, R.; Marinho-Neto, F.d. Infecção por Streptococcus agalactiae em tilápias do Nilo (Oreochromis niloticus). Rev. Ciências Agroveterinárias 2017, 16, 165–169. [Google Scholar] [CrossRef][Green Version]
- Ramesh, D. Effects of potential probiotic Bacillus subtilis KADR1 and its subcellular components on immune responses and disease resistance in Labeo rohita. Aquac. Res. 2017, 49, 1–11. [Google Scholar] [CrossRef]
- Hu, J.; Wang, C.; Ye, L.; Yang, W.; Huang, H.; Meng, F.; Shi, S.; Ding, Z. Anti-tumour immune effect of oral administration of Lactobacillus plantarum to CT26 tumour-bearing mice. J. Biosci. 2015, 40, 269–279. [Google Scholar] [CrossRef]
- Yukgehnaish, K.; Kumar, P.; Sivachandran, P.; Marimuthu, K.; Arshad, A.; Paray, B.A.; Arockiaraj, J. Gut microbiota metagenomics in aquaculture: Factors influencing gut microbiome and its physiological role in fish. Rev. Aquac. 2020, 12, 1903–1927. [Google Scholar] [CrossRef]
- Puvanasundram, P.; Chong, C.M.; Sabri, S.; Yusoff, M.S.; Karim, M. Multi-strain probiotics: Functions, effectiveness and formulations for aquaculture applications. Aquac. Rep. 2021, 21, 100905. [Google Scholar] [CrossRef]
- Eichmiller, J.J.; Hamilton, M.J.; Staley, C.; Sadowsky, M.J.; Sorensen, P.W. Environment shapes the fecal microbiome of invasive carp species. Microbiome 2016, 4, 1–13. [Google Scholar] [CrossRef]
- Li, M.; Li, L.; Huang, T.; Liu, Y.; Lei, A.; Ma, C.; Chen, F.; Chen, M. Effects of attenuated S. agalactiae strain YM001 on intestinal microbiota of Tilapia are recoverable. Front. Microbiol. 2019, 10, 1–14. [Google Scholar] [CrossRef]
- Liu, H.; Guo, X.; Gooneratne, R.; Lai, R.; Zeng, C.; Zhan, F. The gut microbiome and degradation enzyme activity of wild freshwater fishes influenced by their trophic levels. Sci. Rep. 2016, 6, 24340. [Google Scholar] [CrossRef]
- Yi, Y.; Zhang, Z.; Zhao, F.; Liu, H.; Yu, L.; Zha, J.; Wang, G. Probiotic potential of Bacillus velezensis JW: Antimicrobial activity against fish pathogenic bacteria and immune enhancement effects on Carassius auratus. Fish Shellfish. Immunol. 2018, 78, 322–330. [Google Scholar] [CrossRef]
- Standen, B.T.; Rodiles, A.; Peggs, D.L.; Davies, S.J.; Santos, G.A.; Merrifield, D.L. Modulation of the intestinal microbiota and morphology of tilapia, Oreochromis niloticus, following the application of a multi-species probiotic. Appl. Microbiol. Biotechnol. 2015, 99, 8403–8417. [Google Scholar] [CrossRef]
- De Rodrigáñez, M.A.S.; DÍaz-Rosales, P.; Chabrillón, M.; Smidt, H.; Arijo, S.; León-Rubio, J.M.; Alarcón, F.J.; Balebona, M.C.; Moriñigo, M.A.; Cara, J.B.; et al. Effect of dietary administration of probiotics on growth and intestine functionality of juvenile Senegalese solea (Solea senegalensis, Kaup 1858). Aquac. Nutr. 2009, 15, 177–185. [Google Scholar] [CrossRef]
- Tsuchiya, C.; Sakata, T.; Sugita, H. Novel ecological niche of Cetobacterium somerae, an anaerobic bacterium in the intestinal tracts of freshwater fish. Lett. Appl. Microbiol. 2008, 46, 43–48. [Google Scholar] [CrossRef]
- Hao, K.; Wu, Z.Q.; Li, D.L.; Yu, X.B.; Wang, G.X.; Ling, F. Effects of Dietary Administration of Shewanella xiamenensis A-1, Aeromonas veronii A-7, and Bacillus subtilis, Single or Combined, on the Grass Carp (Ctenopharyngodon idella) Intestinal Microbiota. Probiotics Antimicrob. Proteins 2017, 9, 386–396. [Google Scholar] [CrossRef]
- Hoseinifar, S.H.; van Doan, H.; Dadar, M.; Ringø, E.; Harikrishnan, R. Feed Additives, Gut Microbiota, and Health in Finfish Aquaculture. In Microbial Communities in Aquaculture Ecosystems; Springer International Publishing: Cham, Switzerland, 2019; pp. 121–142. [Google Scholar] [CrossRef]
- Gómez, G.D.; Balcázar, J.L. A review on the interactions between gut microbiota and innate immunity of fish. FEMS Immunol. Med. Microbiol. 2008, 52, 145–154. [Google Scholar] [CrossRef]
- Soltani, M.; Kane, A.; Pakzad, K. Effect of the probiotic, Lactobacillus plantarum on growth performance and haematological indices of rainbow trout (Oncorhynchus mykiss) immunized with bivalent streptococcosis/lactococcosis vaccine. Iran. J. Fish. Sci. 2019, 18, 283–295. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient (g.100 g−1) | Control Diet | Probiotic Diet |
|---|---|---|
| Wheat meal | 14.18 | 14.18 |
| Corn | 21.79 | 21.79 |
| Poultry by-product meal | 14.11 | 14.11 |
| Bovine hemoglobin | 13.00 | 13.00 |
| Meat and bone meal | 12.00 | 12.00 |
| Soybean meal | 10.00 | 10.00 |
| Middling rice | 6.00 | 6.00 |
| Rice meal | 4.00 | 4.00 |
| Corn gluten meal | 1.66 | 1.66 |
| Soybean oil | 1.50 | 1.50 |
| Vitamin and mineral supplement (Premix) 1 | 0.50 | 0.50 |
| Probiotics (L. plantarum and B. subtilis) | 0.00 | 0.02 |
| Salt (NaCl) | 0.50 | 0.50 |
| DL-Methionine | 0.20 | 0.20 |
| Vitamin C (35%) | 0.20 | 0.20 |
| Mycotoxin adsorbent (Mycofix® FUM) | 0.10 | 0.10 |
| Antifungal (MOLD-NIL™ MC DRY) | 0.10 | 0.10 |
| Choline chloride (70%) | 0.10 | 0.10 |
| Antioxidant (OXY-NIL™ RX DRY) | 0.05 | 0.05 |
| Calculated chemical composition (%) | ||
| Moisture | 9.36 | 9.36 |
| Crude protein | 36.00 | 36.00 |
| Total lipid | 9.18 | 9.18 |
| Fiber | 3.02 | 3.02 |
| Starch | 25.00 | 25.00 |
| Ash | 8.97 | 8.97 |
| Calcium | 2.36 | 2.36 |
| Phosphorus | 1.40 | 1.40 |
| Digestible energy (kcal kg−1) | 3100 | 3100 |
| Digestible protein | 30.93 | 30.93 |
| Total lysine | 2.70 | 2.70 |
| Total methionine + cysteine | 0.95 | 0.95 |
| Tryptophan | 0.39 | 0.39 |
| Threonine | 1.35 | 1.35 |
| Treatments | Days Post Infection | RPL | |||||||
|---|---|---|---|---|---|---|---|---|---|
| 2 | 3 | 4 | 5 | 6 | 7 | 9 | 14 | ||
| Control (CON) | 4 | 7 | 7 | 10 | 15 | 17 | 17 | 17 | - |
| Adjuvant (ADJ) | 4 | 7 | 7 | 8 | 8 | 10 | 10 | 13 | 28% |
| Probiotics (PRO) | 2 | 4 | 5 | 7 | 8 | 9 | 9 | 9 | 44% |
| Vaccine (VAC) | 1 | 2 | 2 | 2 | 2 | 2 | 4 | 4 | 78% |
| Probiotics + Vaccine (PRO + VAC) | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 94% |
| Alpha Diversity Index | |||
|---|---|---|---|
| Treatment | Chao1 | Shannon | Simpson |
| CON | 599.54 ± 23.14 | 4.79 ± 0.23 | 0.97± 0.01 |
| ADJ | 605.48 ± 52.24 | 4.79 ± 0.31 | 0.97 ± 0.01 |
| PRO | 598.97 ± 90.22 | 4.69 ± 0.25 | 0.97± 0.01 |
| VAC | 566.71 ± 130.22 | 4.59 ± 0.65 | 0.96 ± 0.05 |
| PRO + VAC | 572.08 ± 82.47 | 4.86 ± 0.38 | ± 0.01 |
| PERMANOVA | |||||
|---|---|---|---|---|---|
| CON | ADJ | PRO | VAC | PRO + VAC | |
| CON | - | 0.746 | 0.371 | 0.353 | 0.173 |
| ADJ | 0.746 | - | 0.906 | 0.737 | 0.48 |
| PRO | 0.371 | 0.906 | - | 0.477 | 0.28 |
| VAC | 0.353 | 0.737 | 0.477 | - | 0.847 |
| PRO + VAC | 0.173 | 0.48 | 0.28 | 0.847 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guimarães, M.C.; Cerezo, I.M.; Fernandez-Alarcon, M.F.; Natori, M.M.; Sato, L.Y.; Kato, C.A.T.; Moriñigo, M.A.; Tapia-Paniagua, S.; Dias, D.d.C.; Ishikawa, C.M.; et al. Oral Administration of Probiotics (Bacillus subtilis and Lactobacillus plantarum) in Nile Tilapia (Oreochromis niloticus) Vaccinated and Challenged with Streptococcus agalactiae. Fishes 2022, 7, 211. https://doi.org/10.3390/fishes7040211
Guimarães MC, Cerezo IM, Fernandez-Alarcon MF, Natori MM, Sato LY, Kato CAT, Moriñigo MA, Tapia-Paniagua S, Dias DdC, Ishikawa CM, et al. Oral Administration of Probiotics (Bacillus subtilis and Lactobacillus plantarum) in Nile Tilapia (Oreochromis niloticus) Vaccinated and Challenged with Streptococcus agalactiae. Fishes. 2022; 7(4):211. https://doi.org/10.3390/fishes7040211
Chicago/Turabian StyleGuimarães, Mateus Cardoso, Isabel M. Cerezo, Miguel Frederico Fernandez-Alarcon, Mariene Miyoko Natori, Luciana Yuri Sato, Camila A. T. Kato, Miguel Angel Moriñigo, Silvana Tapia-Paniagua, Danielle de Carla Dias, Carlos Massatoshi Ishikawa, and et al. 2022. "Oral Administration of Probiotics (Bacillus subtilis and Lactobacillus plantarum) in Nile Tilapia (Oreochromis niloticus) Vaccinated and Challenged with Streptococcus agalactiae" Fishes 7, no. 4: 211. https://doi.org/10.3390/fishes7040211
APA StyleGuimarães, M. C., Cerezo, I. M., Fernandez-Alarcon, M. F., Natori, M. M., Sato, L. Y., Kato, C. A. T., Moriñigo, M. A., Tapia-Paniagua, S., Dias, D. d. C., Ishikawa, C. M., Ranzani-Paiva, M. J. T., Cassiano, L. L., Bach, E. E., Clissa, P. B., Orefice, D. P., & Tachibana, L. (2022). Oral Administration of Probiotics (Bacillus subtilis and Lactobacillus plantarum) in Nile Tilapia (Oreochromis niloticus) Vaccinated and Challenged with Streptococcus agalactiae. Fishes, 7(4), 211. https://doi.org/10.3390/fishes7040211