
Effect of Ultrafine Bubbles on Various Stocking Density of Striped Catfish Larviculture in Recirculating Aquaculture System


Abstract
1. Introduction
2. Materials and Methods
2.1. Larvae Description and Experimental Design
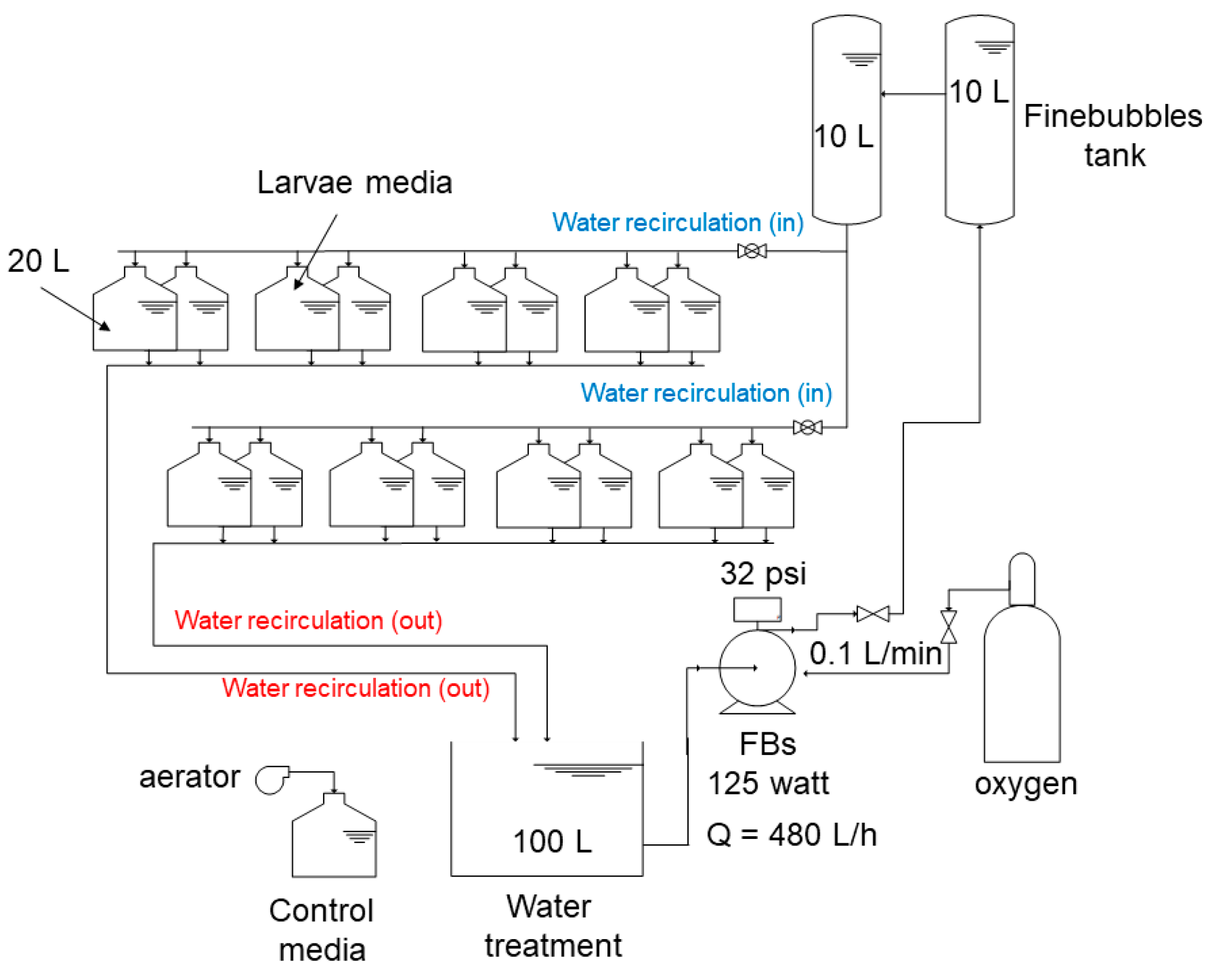
2.2. RAS Design and Operation
2.3. Ultrafine Bubbles and Water Quality Analysis
2.4. Larvae Performances
2.4.1. Percent Volume of the Yolk Sac
- V = yolk sac volume (mm3)
- L = yolk sac length (mm)
- H = yolk sac height (mm)
2.4.2. Yolk Sac Absorption Rate
- g = yolk sac absorption rate (%)
- V0 = early volume of the yolk sac (mm3)
- Vt = final volume of the yolk sac at 36 hah (mm3)
- t – i = time (hours)
2.4.3. Length Growth Rate
- α = length growth rate (%)
- Lt = average length of fish at the end of the sampling period (mm)
- L0 = average length of fish at the beginning of the sampling period (mm)
- t – i = time (hours)
2.4.4. Yolk Sac Utilization Efficiency
- EP = yolk sac utilization rate (%)
- α = length growth rate (%)
- g = yolk sac absorption rate (%)
2.4.5. Survival Rate
- SR = survival rate (%)
- Nt−n = total fish at end exogenous stage (fish)
- Nt−0 = total fish at the initial exogenous stage (fish)
2.4.6. Gross Feeding Efficiency
- Wf = final weight of exogenous stage (mg)
- Wi = initial weight of exogenous stage (mg)
- Fi = feed intake of artemia nauplii (mg)
2.4.7. Specific Growth Rate
- Wf = average final weight of fish (mg)
- Wi = average initial weight of fish (mg)
- t = exogenous stage periods (5 days)
- Lf = final length of exogenous stage (mm)
- Li = initial length of exogenous stage (mm)
- t = exogenous stage periods (5 days)
- Wbio.f = average final weight of biomass (g)
- Wbio.i = average initial weight of biomass (g)
- t = exogenous stage periods (5 days)
2.5. Statistical Analysis
2.6. Water Balance Analysis (WBA)
2.6.1. Oxygen Production
2.6.2. Ammonia Production at Endogenous and Exogenous Stages
2.6.3. The Oxygen Demand and Reserved Oxygen
3. Results and Discussion
3.1. Ultrafine Bubbles Characteristics and Water Quality
3.2. Larvae Performances at Endogenous Stage
3.3. Larvae Performances at Exogenous Stage
3.4. Water Balance Analysis (WBA)
3.4.1. Oxygen Production
3.4.2. Ammonia Production
3.4.3. The Oxygen Demand and Reserved Oxygen
3.4.4. Effect of Water Balance on the Striped Catfish (Pangasianodon hypophthalmus) Larviculture Performance
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- FAO. The State of World Fisheries and Aquaculture, Sustainability in Action; FAO: Rome, Italy, 2020. [Google Scholar]
- Belton, B.; Haque, M.M.; Little, D.C.; Sinh, L.X. Certifying Catfish in Vietnam and Bangladesh: Who Will Make the Grade and Will It Matter? Food Policy 2011, 36, 289–299. [Google Scholar] [CrossRef]
- Ali, H.; Haque, M.M.; Belton, B. Striped Catfish (Pangasianodon Hypophthalmus, Sauvage, 1878) Aquaculture in Bangladesh: An Overview. Aquac. Res. 2013, 44, 950–965. [Google Scholar] [CrossRef]
- Nainggolan, H.; Rahmantya, K.F.; Asianto, A.D.; Wibowo, D.; Wahyuni, T.; Zunianto, A.; Ksatrya, S.P.; Malika, R. Kelautan dan Perikanan dalam Angka Tahun 2018; Pusat Data; Statistik dan Informasi: Jakarta, Indonesia, 2018; pp. 1–386. [Google Scholar]
- Kementerian Kelautan dan Perikanan. Laporan Tahunan Kementerian Kelautan Dan Perikanan 2020; Kementerian Kelautan dan Perikanan: Jakarta, Indonesia, 2020; pp. 1–171. [Google Scholar]
- Fairchild, E.A.; Howell, W.H. Optimal Stocking Density for Juvenile Winter Flounder Pseudopleuronectes Americanus. J. World Aquac. Soc. 2001, 32, 300–308. [Google Scholar] [CrossRef]
- Subagja, J.; Slembrouck, J.; Hung, L.T.; Legendre, M. Larval Rearing of an Asian Catfish Pangasius Hypophthalmus (Siluroidei, Pangasiidae): Analysis of Precocious Mortality and Proposition of Appropriate Treatments. Aquat. Living Resour. 1999, 12, 37–44. [Google Scholar] [CrossRef]
- De Barros, I.B.A.; Villacorta-Correa, M.A.; Carvalho, T.B. Stocking Density and Water Temperature as Modulators of Aggressiveness, Survival and Zootechnical Performance in Matrinxã Larvae, Brycon Amazonicus. Aquaculture 2019, 502, 378–383. [Google Scholar] [CrossRef]
- Wajsbrot, N.; Gasith, A.; Krom, M.D.; Popper, D.M. Acute Toxicity of Ammonia to Juvenile Gilthead Seabream Sparus Aurata under Reduced Oxygen Levels. Aquaculture 1991, 92, 277–288. [Google Scholar] [CrossRef]
- Slembrouck, J.; Baras, E.; Subagja, J.; Hung, L.T.; Legendre, M. Survival, Growth and Food Conversion of Cultured Larvae of Pangasianodon Hypophthalmus, Depending on Feeding Level, Prey Density and Fish Density. Aquaculture 2009, 294, 52–59. [Google Scholar] [CrossRef]
- Xiao, R.; Wei, Y.; An, D.; Li, D.; Ta, X.; Wu, Y.; Ren, Q. A Review on the Research Status and Development Trend of Equipment in Water Treatment Processes of Recirculating Aquaculture Systems. Rev. Aquac. 2019, 11, 863–895. [Google Scholar] [CrossRef]
- Xiao, G.; Cheng, X.; Xie, J.; Zhu, D. Assessment of Aeration Plug-Flow Devices Used with Recirculating Aquaculture Systems on the Growth of Tilapia Oreochromis Niloticus. Aquac. Eng. 2020, 91, 102116. [Google Scholar] [CrossRef]
- Thorarensen, H.; Gústavsson, A.; Gunnarsson, S.; Árnason, J.; Steinarsson, A.; Björnsdóttir, R.; Imsland, A.K.D. The Effect of Oxygen Saturation on the Growth and Feed Conversion of Juvenile Atlantic Cod (Gadus morhua L.). Aquaculture 2017, 475, 24–28. [Google Scholar] [CrossRef]
- Subhan, U.; Iskandar; Zahidah; Joni, I.M. Detection of Reserve Oxygen Potential in the Present of Fine Bubbles and Its Ammonia Removal for Aquaculture Effluent. Mater. Sci. Forum 2021, 1044, 103–111. [Google Scholar] [CrossRef]
- Patel, A.K.; Singhania, R.R.; Chen, C.W.; Tseng, Y.S.; Kuo, C.H.; Wu, C.H.; Dong, C. Di Advances in Micro- and Nano Bubbles Technology for Application in Biochemical Processes. Environ. Technol. Innov. 2021, 23, 101729. [Google Scholar] [CrossRef]
- Subhan, U.; Muthukannan, V.; Azhary, S.Y.; Mulhadi, M.F.; Rochima, E.; Panatarani, C.; Joni, I.M. Development and Performance Evaluation of Air Fine Bubbles on Water Quality of Thai Catfish Rearing. AIP Conf. Proc. 2018, 1927, 0300431–0300435. [Google Scholar] [CrossRef]
- Stejskal, V.; Matousek, J.; Sebesta, R.; Nowosad, J.; Sikora, M.; Kucharczyk, D. Stocking Density Effect on Survival and Growth of Early Life Stages of Maraena Whitefish, Coregonus Maraena (Actinopterygii: Salmoniformes: Salmonidae). Acta Ichthyol. Piscat. 2021, 51, 139–144. [Google Scholar] [CrossRef]
- SNI: 01-6483.4; Produksi Benih Ikan Patin Siam (Pangasius Hyphthalmus) Kelas Benih Sebar. Badan Standar Nasional: Jakarta, Indonesia, 2000; pp. 1–10.
- Heming, T.A.; Buddington, R.K. Yolk Absorption in Embryonic and Larval Fishes. Fish Physiol. 1988, 11, 407–446. [Google Scholar] [CrossRef]
- Watanabe, W.O.; Ellis, S.C.; Ellis, E.P.; Feeley, M.W. Temperature Effects on Eggs and Yolk Sac Larvae of the Summer Flounder at Different Salinities. N. Am. J. Aquac. 1999, 61, 267–277. [Google Scholar] [CrossRef]
- Cadorin, D.I.; da Silva, M.F.O.; Masagounder, K.; Fracalossi, D.M. Interaction of Feeding Frequency and Feeding Rate on Growth, Nutrient Utilization, and Plasma Metabolites of Juvenile Genetically Improved Farmed Nile Tilapia, Oreochromis Niloticus. J. World Aquac. Soc. 2022, 53, 500–515. [Google Scholar] [CrossRef]
- Huguenin, J.E.; Colt, J. Design and Operating Guide for Aquaculture Seawater Systems, 2nd ed.; Elsevier B.V.: Amsterdam, The Netherlands, 2002; Volume 33, ISBN 9789896540821. [Google Scholar]
- Sink, T.D.; Lochmann, R.T.; Pohlenz, C.; Buentello, A.; Gatlin, D. Effects of Dietary Protein Source and Protein-Lipid Source Interaction on Channel Catfish (Ictalurus Punctatus) Egg Biochemical Composition, Egg Production and Quality, and Fry Hatching Percentage and Performance. Aquaculture 2010, 298, 251–259. [Google Scholar] [CrossRef]
- Ebeling, J.M. Recirculating Acuaculture; Cayuga Aqua Ventures: Ithaca, NY, USA, 2010; ISBN 9780971264625. [Google Scholar]
- Emerson, K.; Russo, R.C.; Lund, R.E.; Thurston, R.V. Aqueous Ammonia Equilibrium Calculations: Effect of PH and Temperature. J. Fish. Res. Board Canada 1975, 32, 2379–2383. [Google Scholar] [CrossRef]
- Peykaran Mana, N.; Vahabzadeh, H.; Seidgar, M.; Hafezieh, M.; Pourali, H.R. Proximate Composition and Fatty Acids Profiles of Artemia Cysts, and Nauplii from Different Geographical Regions of Iran. Iran. J. Fish. Sci. 2014, 13, 761–775. [Google Scholar]
- Ownagh, E.; Agh, N.; Noori, F. Comparison of the Growth, Survival and Nutritional Value of Artemia Using Various Agricultural by-Products and Unicellular Algae Dunaliella Salina. Iran. J. Fish. Sci. 2015, 14, 358–368. [Google Scholar]
- Nowosad, J.; Żarski, D.; Biłas, M.; Dryl, K.; Krejszeff, S.; Kucharczyk, D. Dynamics of Ammonia Excretion in Juvenile Common Tench, Tinca tinca (L.), during Intensive Rearing under Controlled Conditions. Aquac. Int. 2013, 21, 629–637. [Google Scholar] [CrossRef]
- Roman, M.R.; Brandt, S.B.; Houde, E.D.; Pierson, J.J. Interactive Effects of Hypoxia and Temperature on Coastal Pelagic Zooplankton and Fish. Front. Mar. Sci. 2019, 6, 139. [Google Scholar] [CrossRef]
- Palińska-Żarska, K.; Żarski, D.; Krejszeff, S.; Nowosad, J.; Biłas, M.; Trejchel, K.; Kucharczyk, D. Dynamics of Yolk Sac and Oil Droplet Utilization and Behavioural Aspects of Swim Bladder Inflation in Burbot, Lota lota L., Larvae during the First Days of Life, under Laboratory Conditions. Aquac. Int. 2014, 22, 13–27. [Google Scholar] [CrossRef]
- Biłas, M.; Zarski, D.; Palińska, K.; Wiszniewska, K.; Kupren, K.; Targońska, K.; Krejszeff, S.; Furgała-Selezniow, G.; Kucharczyk, D. Effect of Stocking Density in Relationship to Bottom Area Sonthe Growth and Survival of Common Barbel Barbus Barbus (L.) Larvae. Polish J. Nat. Sci. 2012, 27, 315–325. [Google Scholar]
- Żarski, D.; Targońska, K.; Krejszeff, S.; Kwiatkowski, M.; Kupren, K.; Kucharczyk, D. Influence of Stocking Density and Type of Feed on the Rearing of Crucian Carp, Carassius carassius (L.), Larvae under Controlled Conditions. Aquac. Int. 2011, 19, 1105–1117. [Google Scholar] [CrossRef]
- Kupren, K.; Zarski, D.; Krejszeff, S.; Kucharczyk, D.; Targońska, K. Effect of Stocking Density on Growth, Survival and Development of Asp Aspius aspius (L.), Ide Leuciscus idus (L.) and Chub Leuciscus cephalus (L.) Larvae during Initial Rearing under Laboratory Conditions. Ital. J. Anim. Sci. 2011, 10, 178–184. [Google Scholar] [CrossRef]
- Zhang, W.; Cao, Z.D.; Peng, J.L.; Chen, B.J.; Fu, S.J. The Effects of Dissolved Oxygen Level on the Metabolic Interaction between Digestion and Locomotion in Juvenile Southern Catfish (Silurus Meridionalis Chen). Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2010, 157, 212–219. [Google Scholar] [CrossRef] [PubMed]
- Purcell, J.; Grover, J. Predation and Food Limitation as Causes of Mortality in Larval Herring at a Spawning Ground in British Columbia. Mar. Ecol. Prog. Ser. 1990, 59, 55–61. [Google Scholar] [CrossRef]
- Ohnari, H. Today’s subjects of microbubble technology. Cem. Eng. Jpn. 2001, 71, 154–159. [Google Scholar] [CrossRef][Green Version]
- Ebina, K.; Shi, K.; Hirao, M.; Hashimoto, J.; Kawato, Y.; Kaneshiro, S.; Morimoto, T.; Koizumi, K.; Yoshikawa, H. Oxygen and Air Nanobubble Water Solution Promote the Growth of Plants, Fishes, and Mice. PLoS ONE 2013, 8, e65339. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Huang, K.; Huang, L.; Hua, Y.; Yu, K.; Liu, T. Effects of Dissolved Oxygen on the Growth Performance, Haematological Parameters, Antioxidant Responses and Apoptosis of Juvenile GIFT (Oreochromis Niloticus). Aquac. Res. 2020, 51, 3079–3090. [Google Scholar] [CrossRef]
- Romano, N.; Scapigliati, G.; Abelli, L. Water Oxygen Content Affects Distribution of t and b Lymphocytes in Lymphoid Tissues of Farmed Sea Bass (Dicentrarchus Labrax). Fishes 2017, 2, 16. [Google Scholar] [CrossRef]
- Burggren, W.; Mwalukoma, A. Respiration during Chronic Hypoxia and Hyperoxia in Larval and Adult Bullfrogs (Rana Catesbeiana). I. Morphological Responses of Lungs, Skin and Gills. J. Exp. Biol. 1983, 105, 191–203. [Google Scholar] [CrossRef] [PubMed]
- Nimura, Y.; Miah, M.I.S. Nitrogen Excretion by Artemia Franciscana. Nippon. Suisan Gakkaishi Jpn. Ed. 1991, 57, 837–844. [Google Scholar] [CrossRef]
- Gross, A.; Boyd, C.E.; Wood, C.W. Nitrogen Transformations and Balance in Channel Catfish Ponds. Aquac. Eng. 2000, 24, 1–14. [Google Scholar] [CrossRef]
- Brune, D.E.; Schwartz, G.; Eversole, A.G.; Collier, J.A.; Schwedler, T.E. Intensification of Pond Aquaculture and High Rate Photosynthetic Systems. Aquac. Eng. 2003, 28, 65–86. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Value |
|---|---|
| Bubbles size distribution means (nm) | 482.9 ± 38.3 |
| Polydispersity Index | 1.342 |
| Zeta potential (mV) | − 23.6 ± 8.20 |
| Electrophoretic mobility (cm2/Vs) | − 0.000183 ± 0.00006 |
| Dissolved oxygen (%sat.) | 255.31 ± 3.90 |
| Temperature (°C) | 26.2 ± 0.15 |
| Water Quality | Endogenous Stage | Exogenous Stage |
|---|---|---|
| Dissolved oxygen (% sat.) | 128.97 ± 6.69 | 128.73 ± 6.17 |
| Temperature (°C) | 26.08 ± 0.19 | 27 ± 0.10 |
| pH | 7.60 ± 0.10 | 7.60 ± 0.10 |
| Total Ammonia Nitrogen (mg/L) | 0.25 ± 0.34 | 0.25 ± 0.34 |
| Ammonia nitrogen, NH3-N (mg/L) | 0.006 ± 0.00 | 0.007 ± 0.00 |
| Ammonium nitrogen, NH4+-N (mg/L) | 0.244 ± 0.00 | 0.243 ± 0.00 |
| Nitrite, NO2-N (mg/L) | 0.0 | 1.75 ± 2.82 |
| Nitrate, NO3-N (mg/L) | 5 ± 0.05 | 3.50 ± 2.60 |
| Tank | Recirculating Flow Rate (Q) | Oxygen Saturation | Concentration (CDO) | Oxygen Production (Poxygen) |
|---|---|---|---|---|
| (L/h) | (Sat%) | (mg/L) | (mg/h) | |
| Ultrafine bubbles tank | 480 | 255.31 | 19.11 | 9172.8 |
| Water treatment tank (endogenous stage) | 480 | 128.97 | 9.46 | 4540.8 |
| Water treatment tank (exogenous stage) | 480 | 128.73 | 9.44 | 4531.2 |
| Each larva media | 30 | 166.03 | 12.18 | 365.4 |
| Term | Stock Density Treatment | ||||
|---|---|---|---|---|---|
| D40C | D80 | D100 | D120 | ||
| Endogenous Stage | |||||
| Lt0 (mm) | Length of larvae at initial period (0 h) in endogenous stage | 3.31 ± 0.21 | 3.31 ± 0.21 | 3.31 ± 0.21 | 3.31 ± 0.21 |
| Survival rate (%) | The survival rate of each stock density at endogenous stage | 100 | 100 | 100 | 100 |
| Ave.Lrate (%) | Average lengths growth rate (Figure 4b) | 2.00 ± 0.09 | 2.02 ± 0.13 | 2.03 ± 0.15 | 1.98 ± 0.08 |
| Feedeff. (%) | Average yolk sac efficiency (Figure 4c) | 50.63 ± 2.82 | 52.03 ± 4.75 | 53.00 ± 3.35 | 50.04 ± 3.21 |
| Wt0 (g), (Equation (12)) | Weight of larvae at initial period | 0.00021 | 0.00021 | 0.00021 | 0.00021 |
| Lt36 (mm), (Equation (13)) | Length of larvae at end endogenous stage (36 hah) | 5.69 | 5.72 | 5.73 | 5.67 |
| Wt36 (g), (Equation (12)) | Weight of larvae at the end of the endogenous phase (36 hah) | 0.00115 | 0.00117 | 0.00118 | 0.00114 |
| Wfeed (g), (Equation (15)) | Weight of the yolk sac | 1.37 | 2.69 | 3.33 | 4.09 |
| Exogenous Stage | |||||
| Lt0 (mm) | Length of larvae at initial period (0 h) in exogenous stage | 8.38 ± 0.33 | 7.72 ± 0.15 | 7.70 ± 0.14 | 7.87 ± 0.35 |
| Survival rate (%) | The survival rate of each stock density at endogenous stage (Figure 5) | 26.79 ± 2.74 | 38.27 ± 0.59 | 42.12 ± 1.88 | 41.38 ± 5.69 |
| Ave.Wrate (%) | Average specific growth weight rate per hour for each stock density (Figure 6) | 1.41 ± 0.08 | 1.30 ± 0.05 | 1.34 ± 0.05 | 1.25 ± 0.06 |
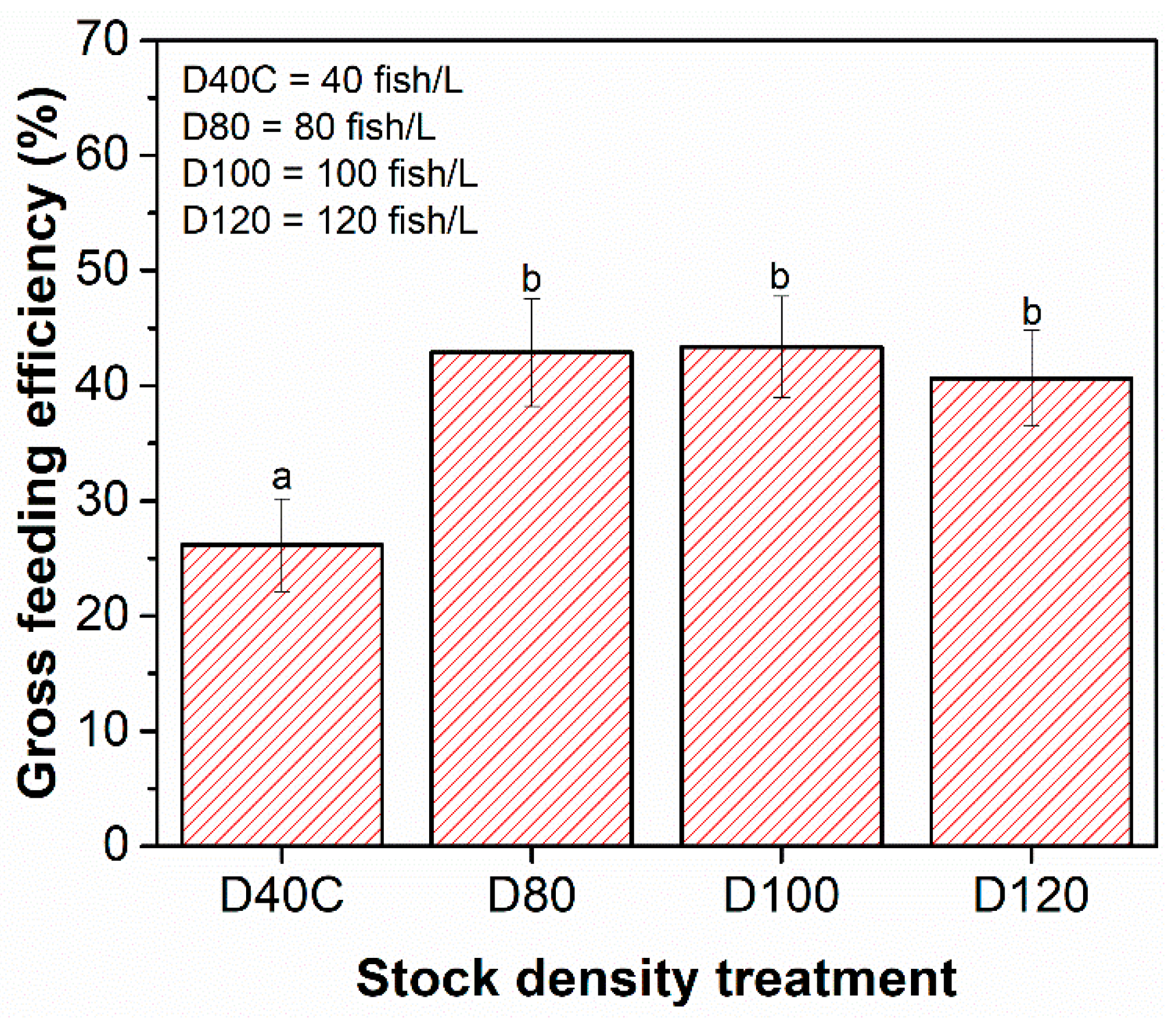
| Feedeff. (%) | Average Gross Feeding Efficiency (GFE) of larvae for each stock density (Figure 8) | 26.17 ± 4.01 | 42.87 ± 4.67 | 43.36 ± 4.40 | 40.65 ± 4.18 |
| Wt0 (g), (Equation (12)) | Weight of larvae at initial period in exogenous stage | 0.00384 | 0.00297 | 0.00295 | 0.00316 |
| Wt120 (g), (Equation (20)) | Weight of larvae at the end of the exogenous stage (120 hah) | 0.01035 | 0.00761 | 0.00771 | 0.00825 |
| Wfeed (g), (Equation (15)) | Weight of the external feed (artemia nauplii) | 6.36 | 8.15 | 11.23 | 15.11 |
| Stages | Stock Density Treatment | Recirculating Flow Rate (Q) | Concentration of Unionized Ammonia (Cun-N) | Production of Unionized Ammonia (Pun-N) | |
|---|---|---|---|---|---|
| (L/h) | (mg/L) | (µg/L) | (mg/h) | ||
| Endogenous | D40C | 30 | 0.0005 | 0.5 | 0.014 |
| D80 | 30 | 0.0009 | 0.9 | 0.027 | |
| D100 | 30 | 0.0011 | 1.1 | 0.034 | |
| D120 | 30 | 0.0014 | 1.4 | 0.041 | |
| Exogenous | D40C | 30 | 0.00006 | 0.06 | 0.002 |
| D80 | 30 | 0.00008 | 0.08 | 0.003 | |
| D100 | 30 | 0.00012 | 0.12 | 0.003 | |
| D120 | 30 | 0.00015 | 0.15 | 0.005 | |
| Stages | Stock Density Treatment | Recirculating Flow Rate | Oxygen Supply in Each Larvae Media | Measured Oxygen in Water Treatment Box | Oxygen Demand for Metabolism | Oxygen Demand for Nitrifying Bacteria | Oxygen Demand for Direct Oxidation Unionized Ammonia (NH3-N) | Reserved Oxygen | |
|---|---|---|---|---|---|---|---|---|---|
| (L/h) | (mg/h) | (mg/h) | (mg/h) | (mg/h) | (mg/h) | (mg/h) | (mg/L) | ||
| Endogenous | D40C | 30 | 365.48 | 283.80 | 9.49 | 4.55 | 1.62 | 66.02 | 2.20 |
| D80 | 30 | 365.48 | 283.80 | 18.71 | 8.98 | 3.18 | 50.81 | 1.69 | |
| D100 | 30 | 365.48 | 283.80 | 23.11 | 11.09 | 3.93 | 43.55 | 1.45 | |
| D120 | 30 | 365.48 | 283.80 | 28.43 | 13.65 | 4.84 | 34.77 | 1.16 | |
| Exogenous | D40C | 30 | 365.48 | 283.20 | 13.25 | 6.36 | 0.23 | 62.44 | 2.08 |
| D80 | 30 | 365.48 | 283.20 | 16.98 | 8.15 | 0.29 | 56.86 | 1.90 | |
| D100 | 30 | 365.48 | 283.20 | 23.39 | 11.23 | 0.40 | 47.26 | 1.58 | |
| D120 | 30 | 365.48 | 283.20 | 31.48 | 15.11 | 0.54 | 35.15 | 1.17 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Subhan, U.; Iskandar; Zahidah; Panatarani, C.; Joni, I.M. Effect of Ultrafine Bubbles on Various Stocking Density of Striped Catfish Larviculture in Recirculating Aquaculture System. Fishes 2022, 7, 190. https://doi.org/10.3390/fishes7040190
Subhan U, Iskandar, Zahidah, Panatarani C, Joni IM. Effect of Ultrafine Bubbles on Various Stocking Density of Striped Catfish Larviculture in Recirculating Aquaculture System. Fishes. 2022; 7(4):190. https://doi.org/10.3390/fishes7040190
Chicago/Turabian StyleSubhan, Ujang, Iskandar, Zahidah, Camellia Panatarani, and I Made Joni. 2022. "Effect of Ultrafine Bubbles on Various Stocking Density of Striped Catfish Larviculture in Recirculating Aquaculture System" Fishes 7, no. 4: 190. https://doi.org/10.3390/fishes7040190
APA StyleSubhan, U., Iskandar, Zahidah, Panatarani, C., & Joni, I. M. (2022). Effect of Ultrafine Bubbles on Various Stocking Density of Striped Catfish Larviculture in Recirculating Aquaculture System. Fishes, 7(4), 190. https://doi.org/10.3390/fishes7040190