
Comparative Transcriptome Analysis of Differentially Expressed Genes in the Testis and Ovary of Sea Urchin (Strongylocentrotus intermedius)

Abstract
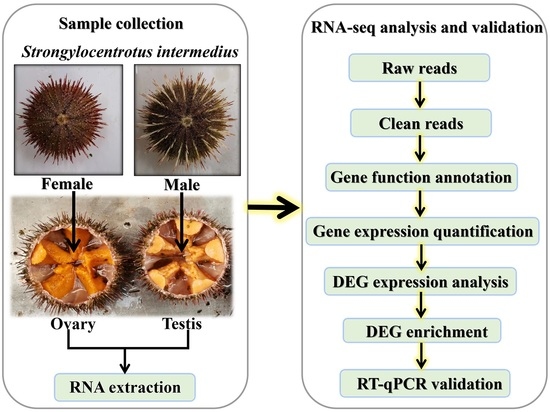
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Sample, RNA Preparation, and cDNA Library Construction
2.3. De Novo Transcriptome Assembly, Mapping and Annotation
2.4. Gene Expression Quantification and Differentially Expressed Gene Enrichment Analysis
2.5. RT-qPCR Validation
3. Results
3.1. Illumina Sequencing and De Novo Assembly
3.2. Annotation and Functional Classification
3.3. DEGs Identification and Enrichment Analysis
3.4. RT-qPCR Validation of DEGs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Sodergren, E.; Weinstock, G.M.; Davidson, E.H.; Cameron, R.A.; Gibbs, R.A.; Angerer, R.C.; Angerer, L.M.; Arnone, M.I.; Burgess, D.R.; Burke, R.D.; et al. The genome of the sea urchin Strongylocentrotus purpuratus. Science 2006, 314, 941–952. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.W.; Qin, L.; Zhou, D.Y.; Wu, H.T.; Jing, W.; Yang, J.F.; Li, D.M.; Dong, X.P.; Yoshiyuki, M. Extraction of lipid from sea urchin (Strongylocentrotus nudus) gonad by enzyme-assisted aqueous and supercritical carbon dioxide methods. Eur. Food Res. Techno. 2010, 230, 737–743. [Google Scholar] [CrossRef]
- Chen, Y.-D.; Chang, Y.-Q.; Wang, X.-L.; Qiu, X.-M.; Liu, Y. De novo assembly and analysis of tissue-specific transcriptomes revealed the tissue-specific genes and profile of immunity from Strongylocentrotus intermedius. Fish Shellfish Immunol. 2015, 46, 723–736. [Google Scholar] [CrossRef] [PubMed]
- Jia, Z.-Y.; Wang, Q.; Wu, K.-K.; Wei, Z.-L.; Zhou, Z.C.; Liu, X.-L. De novo transcriptome sequencing and comparative analysis to discover genes involved in ovarian maturity in Strongylocentrotus nudus. Comp. Biochem. Physiol. Part D Genom. Proteom. 2017, 23, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.-Y.; Pan, N.; Wu, J.-N.; Liu, S.; Liu, Z.; Chen, L. Analysis and evaluation of nutritional composition of sea urchin (Anthocidaris crassispina) gonad. Fish. Mod. 2017, 44, 50–55. (In Chinese) [Google Scholar]
- Zhao, C.; Sun, P.; Zhou, H.-S.; Tian, X.-F.; Feng, W.-P.; Chang, Y.-Q. Heritability and phenotypic correlations of gonad sweetness in the sea urchin Strongylocentrotus intermedius. Aquacult. Int. 2014, 22, 1737–1742. [Google Scholar] [CrossRef]
- Feng, W.-P.; Chang, Y.-Q.; Zhao, C.; Sun, P.; Wei, J.; Hou, S. Effects of inbreeding on growth, gametogenesis, gonad production, quality and MYP expression in the sea urchin Strongylocentrotus intermedius. Aquacult. Int. 2015, 23, 903–912. [Google Scholar] [CrossRef]
- Chen, G.-Q.; Xiang, W.-Z.; Lau, C.; Peng, J.; Qiu, J.W.; Chen, F.; Jiang, J. A comparative analysis of lipid and carotenoid composition of the gonads of Anthocidaris crassispina, Diadema setosum and Salmacis sphaeroides. Food Chem. 2010, 120, 973–977. [Google Scholar] [CrossRef]
- Murata, Y.; Yokoyama, M.; Unuma, T.; Sata, N.; Kuwahara, R.; Kaneniwa, M. Seasonal changes of bitterness and pulcherrimine content in gonads of green sea urchin Hemicentrotus pulcherrimus at Iwaki in Fukushima Prefecture. Fish. Sci. 2002, 68, 184–189. [Google Scholar] [CrossRef][Green Version]
- Ghisaura, S.; Loi, B.; Biosa, G.; Baroli, M.; Pagnozzi, D.; Roggio, T.; Uzzau, S.; Anedda, R.; Addis, M. Proteomic changes occurring along gonad maturation in the edible sea urchin Paracentrotus lividus. J. Proteomics 2016, 144, 63–72. [Google Scholar] [CrossRef]
- Zhou, X.; Zhou, D.-Y.; Lu, T.; Liu, Z.-Y.; Zhao, Q.; Liu, Y.-X.; Hu, X.-P.; Zhang, J.-H.; Shasidi, F. Characterization of lipids in three species of sea urchin. Food Chem. 2018, 241, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Dincer, T.; Cakli, S. Chemical composition and biometrical measurements of the Turkish sea urchin (Paracentrotus lividus, lamarck, 1816). Crit. Rev. Food Sci. Nutr. 2007, 47, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Komata, Y. Study of the extractives of ‘‘Uni” IV. Taste of each component in the extractives. Bull. Jpn. Soc. Fish. Sci. 1964, 30, 749–756. [Google Scholar] [CrossRef]
- Osako, K.; Kiriyama, T.; Ruttanapornvareesakul, Y. Free amino acid compositions of the gonad of the wild and cultured sea urchins Anthocidaris crassispina. Aquacult. Sci. 2006, 54, 301–304. [Google Scholar]
- Inomata, E.; Murata, Y.; Matsui, T.; Agatsuma, Y. Gonadal production and quality in the sea urchin Mesocentrotus nudus fed a high-protein concentrated red alga Pyropia yezoensis. Aquaculture 2015, 454, 184–191. [Google Scholar] [CrossRef]
- Ayyagari, A.; Babu, K. Nutrient composition and antioxidant activity of gonads of sea urchin Stomopneustes variolaris. Food Chem. 2016, 197, 597–602. [Google Scholar]
- Li, L.; Yang, D.; Qi, S.-B.; Zuo, R.-T.; Wang, H.; Ding, J.; Chang, Y.-Q. Changes in lipids and fatty acids in gonads of sea urchin Strongylocentrotus intermedius before and after reproduction. J. Dalian Ocean. Univ. 2018, 33, 423–429. (In Chinese) [Google Scholar]
- Ding, J.; Chang, Y.Q.; Hao, Z.L.; Zhang, B. Comparative studies on urchin gonad fatty acid composition and β-carotene content in north China sea section. J. Agric. Sci. Technol. 2011, 13, 122–128. (In Chinese) [Google Scholar]
- Zhao, C.; Zhang, W.-J.; Chang, Y.Q.; Liu, P. Test and gonad characteristics in different genders of cultivated sea urchins (Strongylocentrotus intermedius, agassiz): First insight into sexual identification. Afr. J. Biotechnol. 2010, 9, 7560–7563. [Google Scholar]
- Phillips, K.; Bremer, P.; Silcock, P.; Hamid, N.; Delahunty, C.; Barker, M.; Kissick, J. Effect of gender, diet and storage time on the physical properties and sensory quality of sea urchin (Evechinus chloroticus) gonads. Aquaculture 2009, 288, 205–215. [Google Scholar] [CrossRef]
- Kong, D.-R.; Cui, J.; Wang, Z.-C.; Sun, H.; Hu, Z.-W.; Li, X.; Qiu, X.-M.; Jiang, C.; Liu, H.-Y.; Zhang, T.; et al. The regulatory networks conferred by IFN-γ in the kidney of Takifugu rubripes. Int. J. Agric. Biol. 2018, 20, 2189–2195. [Google Scholar]
- Cui, J.; Liu, S.-K.; Zhang, B.; Wang, H.-D.; Sun, H.-J.; Song, S.-H.; Qiu, X.-M.; Liu, Y.; Wang, X.-L.; Jiang, Z.-Q.; et al. Transciptome analysis of the gill and swimbladder of Takifugu rubripes by RNA-Seq. PLoS ONE 2014, 9, e85505. [Google Scholar] [CrossRef] [PubMed]
- Grabherr, M.; Haas, B.; Yassour, M.; Levin, J.; Thompson, D.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.-D.; et al. Full-length transcriptome assembly from RNA-seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef]
- Eddy, S. Profile hidden Markov models. Bioinformatics 1998, 14, 755–763. [Google Scholar] [CrossRef] [PubMed]
- Xie, C.; Mao, X.-Z.; Huang, J.-J.; Ding, Y.; Wu, J.-M.; Dong, S.; Kong, L.; Gao, G.; Li, C.-Y.; Wei, L.-P. Kobas 2.0: A web server for annotation and identification of enriched pathways and diseases. Nucleic Acids Res. 2011, 39, W316–W322. [Google Scholar] [CrossRef]
- Wang, Z.-C.; Qiu, X.-M.; Kong, D.-R.; Zhou, X.-X.; Guo, Z.-B.; Gao, C.-F.; Ma, S.; Hao, W.-W.; Jiang, Z.-Q.; Liu, S.-C.; et al. Comparative RNA-Seq analysis of differentially expressed genes in the testis and ovary of Takifugu rubripes. Comp. Biochem. Physiol. Part DGenom. Proteom. 2017, 22, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Dillies, M.; Rau, A.; Aubert, J.; Hennequet-Antier, C.; Jeanmougin, M.; Servant, N.; Keime, C.; Marot, G.; Castel, D.; Estelle, J.; et al. Comprehensive evaluation of normalization methods for Illumina high-throughput RNA sequencing data analysis. Brief. Bioinform. 2013, 14, 671–683. [Google Scholar] [CrossRef]
- Wang, F.-C.; Yang, J.; Wang, H.-B.; Xia, G.-L. Gonadotropin-regulated expressions of lanosterol 14α-demethylase, sterol Δ14-reductase and C-4 sterol methyl oxidase contribute to the accumulation of meiosis-activating sterol in rabbit gonads. Prostag. Oth. Lipid Mediat. 2010, 92, 25–32. [Google Scholar] [CrossRef]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome Biol. 2010, 11, 106. [Google Scholar] [CrossRef]
- Klopfenstein, D.V.; Zhang, L.; Pedersen, B.S.; Ramírez, F.; Vesztrocy, A.W.; Naldi, A.; Mungall, C.J.; Yunes, J.M.; Botvinnik, O.; Weigel, M.; et al. GOATOOLS: A Python library for Gene Ontology analyses. Sci. Rep. 2018, 18, 10872. [Google Scholar] [CrossRef]
- Lalitha, S. Primer Premier 5. Biotech. Softw. Internet Rep. 2000, 1, 270–272. [Google Scholar] [CrossRef]
- Livak, K.; Schmittgen, T. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I. Trinity: Reconstructing a full-length transcriptome without a genome from RNA-Seq data. Nat. Biotechnol. 2013, 29, 644. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.-W.; Cui, X.; Shen, X.-F.; Wang, L.-S.; Jiang, L.; Liu, H.-Y.; Liu, Y.; Liu, Q.; Jiang, C. De novo transcriptome analysis and differentially expressed genes in the ovary and testis of the Japanese mantis shrimp Oratosquilla oratoria by RNA-Seq. Comp. Biochem. Physiol. Part D Genom. Proteom. 2018, 26, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Mcbride, S.; Price, R.; Tom, P.; Lawrence, J.; Lawrence, A. Comparison of gonad quality factors: Color, hardness and resilience, of Strongylocentrotus franciscanus between sea urchins fed prepared feed or algal diets and sea urchins harvested from the northern California fishery. Aquaculture 2004, 233, 405–422. [Google Scholar] [CrossRef]
- Murata, Y.; Sata, N.; Yokoyama, M.; Kuwahara, R.; Oohara, I. Determination of a novel bitter amino acid, pulcherrimine, in the gonad of the green sea urchin Hemicentrotus pulcherrimus. Fish. Sci. 2001, 67, 341–345. [Google Scholar] [CrossRef]
- Xu, W.-D.; Han, P.; Wang, L.-M.; Zhou, Z.-C.; Han, J.-B. The amino acid compositions of gonads in sea urchin Strongylocentrotus nudus. J. Dalian Ocean. Univ. 2009, 24, 583–586. (In Chinese) [Google Scholar]
- Murata, Y.; Yamamoto, T.; Kaneniwa, M.; Kuwahara, R.; Yokoyama, M. Occurrence of bitter gonad in Hemicentrotus pulcherrimus. Nippon. Suisan Gakkaishi 1998, 64, 477–478. [Google Scholar] [CrossRef][Green Version]
- Li, Y.; Chen, G.-K.; Tong, X.-W.; Zhang, H.-T.; Liu, X.-G.; Liu, Y.-H.; Lu, F.-P. Construction of Escherichia coli strains producing L-serine from glucose. Biotechnol. Lett. 2012, 34, 1525–1530. [Google Scholar] [CrossRef]
- Deng, Y.-H.; Ran, W.; Man, S.-Q.; Li, X.-P.; Gao, H.-J.; Tang, T.; Tachibana, H.; Cheng, X.-J. Artemether exhibits amoebicidal activity against acanthamoeba castellanii through inhibition of the serine biosynthesis pathway. Antimicrob. Agents Chemother. 2015, 59, 4680–4688. [Google Scholar] [CrossRef]
- Basurko, M.J.; Marche, M.; Darriet, M.; Cassaigne, A. Phosphoserine aminotra-nsferase, the second step-catalyzing enzyme for serine biosynthesis. IUBMB Life 1999, 48, 525–529. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, Y.; Sakuraba, H.; Doi, K.; Ohshima, T. Molecular and functional characterization of D-3-phosphoglycerate dehydrogenase in the serine biosynthetic pathway of the hyperthermophilic archaeon Sulfolobus tokodaii. Arch. Biochem. Biophys. 2008, 470, 120–128. [Google Scholar] [CrossRef] [PubMed]
- MacFarlane, A.; Liu, X.-W.; Perry, C.; Flodby, P.; Allen, R.; Stabler, S.; Stover, P. Cytoplasmic serine hydroxymethyltransferase regulates the metabolic partitioning of methylenetetrahydrofolate but is not essential in mice. J. Biol. Chem. 2008, 283, 25846–25853. [Google Scholar] [CrossRef]
- Grant, G.; Hu, Z.-Q.; Xu, X.-L. Amino acid residue mutations uncouple cooperative effects in Escherichia coli D-3-phosphoglycerate dehydrogenase. J. Bio. Chem. 2001, 276, 17844–17850. [Google Scholar] [CrossRef]
- Grant, G.; Xu, X.L.; Hu, Z. The relationship between effector binding and inhibition of activity in D-3-phosphoglycerate dehydrogenase. Protein Sci. 1999, 8, 2501–2505. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Grant, G. Transient kinetic analysis of L-serine Interaction with Escherichia coli D-3-phosphoglycerate dehydrogenase containing amino acid mutations in the hinge regions. Biochemistry 2011, 50, 2900–2906. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Dey, S.; Grant, G.; Sacchettini, J. Crystal structure of mycobacterium tuberculosis D-3-phosphoglycerate dehydrogenase: Extreme asymmetry in a tetramer of identical subunits. J. Bio. Chem. 2005, 280, 14892–14899. [Google Scholar] [CrossRef]
- Zhang, W.; Zhang, M.; Gao, C.; Zhang, Y.-P.; Ge, Y.-S.; Guo, S.-T.; Guo, X.-T.; Zhou, Z.-K.; Liu, Q.-Y.; Zhang, Y.-X.; et al. Coupling between d-3-phosphoglycerate dehydrogenase and d-2-hydroxyglutarate dehydrogenase drives bacterial l-serine synthesis. Proc. Natl. Acad. Sci. USA 2017, 114, E7574–E7582. [Google Scholar] [CrossRef]
- Kaneko, K.; Matsumoto, H.; Shirai, T.; Kamei, M.; Okazaki, E.; Osako, K. Seasonal variations in free amino acid composition and taste aspects of black sea urchin, Diadema setosum, gonad. Food Sci. Technol. Res. 2012, 18, 835–842. [Google Scholar] [CrossRef][Green Version]
- Woods, C.; James, P.; Moss, G.; Wright, J.; Siikavuopio, S. A comparison of the effect of urchin size and diet on gonad yield and quality in the sea urchin. Aquac. Int. 2008, 16, 49–68. [Google Scholar] [CrossRef]
- Azad, A.; Pearce, C.; Mckinley, R. Effects of diet and temperature on ingestion, absorption, assimilation, gonad yield, and gonad quality of the purple sea urchin (Strongylocentrotus purpuratus). Aquaculture 2011, 317, 187–196. [Google Scholar] [CrossRef]
- 52 Wilson, P.; Gardner, S.; Lambie, N.; Commans, S.; Crowther, D. Characterization of the human patatin-like phospholipase family. J. Lipid R. 2006, 47, 1940–1949. [Google Scholar] [CrossRef] [PubMed]
- Rotman, Y.; Koh, C.; Zmuda, J.; Kleiner, D.; Liang, T.; Nash, C. The association of genetic variability in patatin-like phospholipase domain-containing protein 3 (PNPLA3) with histological severity of nonalcoholic fatty liver disease. Hepatology 2010, 52, 894–903. [Google Scholar] [CrossRef] [PubMed]
- Viganò, M.; Valenti, L.; Lampertico, P.; Facchetti, F.; Motta, B.; D’Ambrosio, R.; Romagnoli, S.; Dongiovanni, P.; Donati, B.; Fargion, S.; et al. Patatin-like phospholipase domain-containing 3 I148M affects liver steatosis in patients with chronic hepatitis B. Hepatology 2013, 58, 1245–1252. [Google Scholar] [CrossRef]
- Chang, Y.-Q.; Zhang, W.-J.; Ding, J.; Shi, S.-B.; Zhao, C.; Zhang, B. Comparison of gonad traits among families and between genders of the sea urchin Strongylocentrotus intermedius. J. Fish. China 2010, 34, 1080–1088. [Google Scholar]
- Tong, S.-Y.; Chen, W.; You, X.-C.; Liu, H.-L. Study on lipid and fatty acids composition of three kinds of echinoidea’s gonad. J. Fish. China 1998, 22, 247–252. (In Chinese) [Google Scholar]
- Hathaway, R. Conversion of estradiol-17β by sperm preparations of sea urchins and oysters. Gen. Comp. Endocrinol. 1965, 5, 504–508. [Google Scholar] [CrossRef]
- Wang, C.; Croll, R. Effects of sex steroids on gonadal development and gender determination in the sea scallop, Placopectenmagellanicus. Aquaculture 2004, 238, 483–498. [Google Scholar] [CrossRef]
- Kiyomoto, M.; Kikuchi, A.; Morinaga, S.; Unuma, T.; Yokota, Y. Exogastrulation and interference with the expression of major yolk protein by estrogens administered to sea urchins. Cell Biol. Toxicol. 2008, 24, 611–620. [Google Scholar] [CrossRef]
- Leenders, F.; Adamski, J.; Husen, B.; Thole, H.; Jungblut, P. Molecular cloning and amino acid sequence of the porcine 17beta-estradiol dehydrogenase. Eur. J. Biochem. 1994, 222, 221–227. [Google Scholar] [CrossRef]
- Ohno, S.; Nishikawa, K.; Honda, Y.; Nakajin, S. Expression in E. coli and tissue distribution of the human homologue of the mouse Ke 6 gene, 17beta-hydroxysteroid dehydrogenase type 8. Mol. Cell Biochemi. 2008, 309, 209–215. [Google Scholar] [CrossRef]
- Rotinen, M.; Celay, J.; Alonso, M.; Arrazola, A.; Encio, I.; Villar, J. Estradiol induces type 8 17beta-hydroxysteroid dehydrogenase expression: Crosstalk between estrogen receptor alpha and C/EBPbeta. J. Endocrinol. 2009, 200, 85–92. [Google Scholar] [CrossRef] [PubMed]
- 63 Smuc, T.; Rizner, T. Expression of 17beta-hydroxysteroid dehydrogenases and other estrogen-metabolizing enzymes in different cancer cell lines. Chem. Biol. Interact. 2009, 178, 228–233. [Google Scholar] [CrossRef] [PubMed]
- 64 Liu, J.-G.; Zhang, Z.-F.; Ma, X.-S.; Liang, S.-S.; Yang, D.-D. Characteristics of 17β-hydroxysteroid dehydrogenase 8 and its potential role in gonad of Zhikong scallop Chlamys farreri. J. Steroid. Biochem. Mol. Biol. 2014, 141, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Motohara, K.; Tashiro, H.; Taura, Y.; Ohba, T.; Katabuchi, H. Immunohistochemical analysis of 17β-hydroxysteroid dehydrogenase isozymes in human ovarian surface epithelium and epithelial ovarian carcinoma. Med. Mol. Morphol. 2010, 43, 197–203. [Google Scholar] [CrossRef]
- Hilborn, E.; Stål, O.; Jansson, A. Estrogen and androgen-converting enzymes 17β-hydroxysteroid dehydrogenase and their involvement in cancer: With a special focus on 17β-hydroxysteroid dehydrogenase type 1, 2, and breast cancer. Oncotarget 2017, 8, 30552–30562. [Google Scholar] [CrossRef]
- Chen, Z.-J.; Kastaniotis, A.; Miinalainen, I.; Rajaram, V.; Wierenga, R.; Hiltunen, J. 17β-hydroxysteroid dehydrogenase type 8 and carbonyl reductase type 4 assemble as a ketoacyl reductase of human mitochondrial FAS. FASEB J. 2009, 23, 3682–3691. [Google Scholar] [CrossRef]
- Jin, Q.; Ren, Y.; Wang, M.; Suraneni, P.; Crispino, J.; Fan, J.; Huang, Z. Novel function of faxdc2 in megakaryopoiesis. Blood Cancer J. 2016, 6, 478. [Google Scholar] [CrossRef]
- Machlus, K.; Italiano, J. The incredible journey: From megakaryocyte development to platelet formation. J. Cell Biol. 2013, 201, 785–796. [Google Scholar] [CrossRef]
- Parra, M.; Rubilar, T.; Latorre, M.; Epherra, L.; Gil, D. Nutrient allocation in the gonads of the sea urchin Arbacia dufresnii in different stages of gonadal development. Invertebr. Reprod. Dev. 2014, 59, 26–36. [Google Scholar] [CrossRef]
- Bard, M.; Bruner, D.; Pierson, C.; Lees, N.; Biermann, B.; Frye, L.; Koegel, C.; Barbuch, R. Cloning and characterization of ERG25, the Saccharomyces cerevisiae gene encoding C-4 sterol methyl oxidase. Proc. Natl. Acad. Sci. USA 1966, 93, 186–190. [Google Scholar] [CrossRef]
- Yao, J.N.; Liao, N.Q.; Li, H.M. Characterization and bioinformatics analysis of c-4 sterol methyl oxidase from Monascus purpureus. Appl. Mech. Mater. 2014, 522–524, 247–250. [Google Scholar] [CrossRef]
- Pollier, J.; Vancaester, E.; Kuzhiumparambil, U.; Vickers, C.; Klaas, V.K.; Goossens, A.; Fabris, M.; Kuzhiumparambil, U. A widespread alternative squalene epoxidase participates in eukaryote steroid biosynthesis. Nat. Microbiol. 2019, 4, 226–233. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene ID | Primer Sequences (5′–3′) |
|---|---|---|
| 18sRNA | F:GCGTTTAGCCAGGCGAGAT; R:GGAGGAGGTTCAGCGGATTA | |
| SHMT | DN77177_c3_g1 | F: CCCTGTATGGCATTGACT; R:GCCTTAGCAGCATTTGGT |
| PGDH | DN73867_c0_g1 | F:TCAGCAGCACTTAGGGTATT; R:TGTCATCAAAGCAGGCAC |
| PNPLA7 | DN82957_c4_g2 | F:CCAGAGGCATCAACTTCG; R:AAACAAGCCATTCCAGACCC |
| FAXDC2 | DN68245_c4_g1 | F:AATAGAAGCCAAACTCCTCG; R:GGTGCTCGCTAACCAGACAG |
| C4MO | DN69430_c0_g1 | F:CCGCTTACCTTACTCAAT; R:AGCAATCCACCTGTATCT |
| Gstt1 | DN56592_c2_g2 | F:TGGATAGAGTGAAGGCTTGT; R:TGTTGGCTGAAACCTGATGT |
| MAN2B1 | DN80637_c2_g1 | F:TTCTGGAGCGGGTATTTCGT; R:GTAGCCTGATTGACTGGGAG |
| RARS | DN83813_c9_g2 | F:ACGCAACACGAAGGGCAGTC; R:GTAGCCTGATTGACTGGGAG |
| Adprh | DN77147_c2_g2 | F:GCATCAGATCCCACATAC; R:AGCAGGACCAAATACCAT |
| HSP83 | DN71588_c2_g2 | F:TCTCACCCTCATTGACACCG; R:TCCAACACCAAATTGACCGA |
| CHDH | DN79960_c0_g2 | F:CGGACCGAAAGATAACAC; R:GTTGATGGAGGAAGAGCC |
| 17β-HSD8 | DN62887_c3_g3 | F:GACAAGAGCCCTGAAGAC; R:TTGAGACGATTGGTGGTT |
| Samples | Raw Reads | Clean Reads | Q30% | Clean Bases (bp) | GC Content(%) | Mapped Reads |
|---|---|---|---|---|---|---|
| Male gonad | 43,797,146 | 41,959,814 | 94.97 | 6,174,663,478 | 41.64 | 13,462,822 |
| Female gonad | 56,222,782 | 54,537,612 | 93.61 | 7,972,005,581 | 42.76 | 18,319,817 |
| Length (bp) | Number of Transcripts (Percent) | Number of Unigenes (Percent) |
|---|---|---|
| 500 | 54,965 (43%) | 46,236 (54%) |
| 501–1000 | 32,733 (25%) | 18,254 (21%) |
| 1001–1500 | 17,592 (14%) | 7980 (9%) |
| 1501–2000 | 9645 (7%) | 4603 (5%) |
| 2001–2500 | 5464 (4%) | 2918 (3%) |
| 2501–3000 | 3206 (2%) | 1898 (2%) |
| 3001–3500 | 1843 (1%) | 1181 (1%) |
| 3501–4000 | 1181 (1%) | 841 (1%) |
| 4001–4500 | 723 (1%) | 524 (1%) |
| >4500 | 1627 (1%) | 1310 (2%) |
| Gene Name | Description | FPKM Testis | FPKM Ovary | Log2 Fold Change |
|---|---|---|---|---|
| PNPLA7 | patatin-like phospholipase domain-containing protein 7 | 35.258851 | 7.297912 | 2.2724 |
| 17β-HSD8 | estradiol 17-beta-dehydrogenase 8 | 0 | 39.011632 | −6.2858 |
| C4MO | C-4 methylsterol oxidase-like | 18.693844 | 54.036744 | −1.5314 |
| FAXDC2 | fatty acid hydroxylase domain-containing protein 2 | 169.64164 | 64.822629 | 1.3879 |
| PGDH | D-3-phosphoglycerate dehydrogenase | 7.7170316 | 36.972509 | −2.2603 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gou, P.; Wang, Z.; Yang, J.; Wang, X.; Qiu, X. Comparative Transcriptome Analysis of Differentially Expressed Genes in the Testis and Ovary of Sea Urchin (Strongylocentrotus intermedius). Fishes 2022, 7, 152. https://doi.org/10.3390/fishes7040152
Gou P, Wang Z, Yang J, Wang X, Qiu X. Comparative Transcriptome Analysis of Differentially Expressed Genes in the Testis and Ovary of Sea Urchin (Strongylocentrotus intermedius). Fishes. 2022; 7(4):152. https://doi.org/10.3390/fishes7040152
Chicago/Turabian StyleGou, Panpan, Zhicheng Wang, Jin Yang, Xiuli Wang, and Xuemei Qiu. 2022. "Comparative Transcriptome Analysis of Differentially Expressed Genes in the Testis and Ovary of Sea Urchin (Strongylocentrotus intermedius)" Fishes 7, no. 4: 152. https://doi.org/10.3390/fishes7040152
APA StyleGou, P., Wang, Z., Yang, J., Wang, X., & Qiu, X. (2022). Comparative Transcriptome Analysis of Differentially Expressed Genes in the Testis and Ovary of Sea Urchin (Strongylocentrotus intermedius). Fishes, 7(4), 152. https://doi.org/10.3390/fishes7040152