
Neuropeptide Y in Spotted Scat (Scatophagus Argus), Characterization and Functional Analysis towards Feed Intake Regulation

 ,
,  ,
,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Fish
2.2. Cloning and Sequence Analysis of the Npy Gene
2.3. Npy Expression Patterns in Different Spotted Scat Tissues, RNA Extraction and cDNA Synthesis
2.4. Npy Gene Localization in the Brain of the Spotted Scat via In Situ Hybridization
2.5. Post-Prandial Research and Long-Term Feeding, Fasting, and Refeeding of Spotted Scat
2.6. Npy Peptide Synthesis and In Vitro Incubation of the Spotted Scat
2.7. Npy Peptide In Vivo Injection of the Spotted Scat
2.8. Quantitative Real-Time PCR (qPCR)and Statistical Analysis
3. Results
3.1. Cloning and Sequence Analysis of the Npy Gene

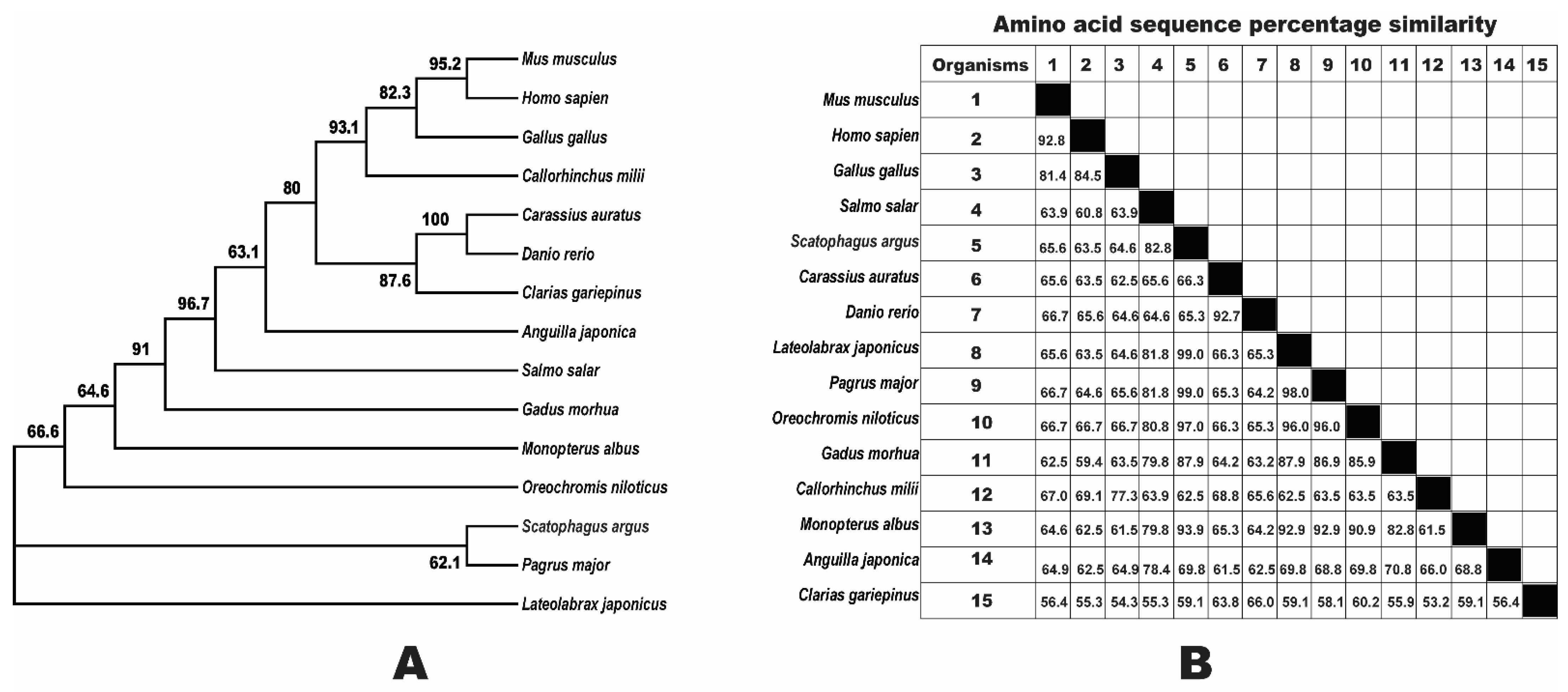
3.2. Phylogenetic Analysis of Npy
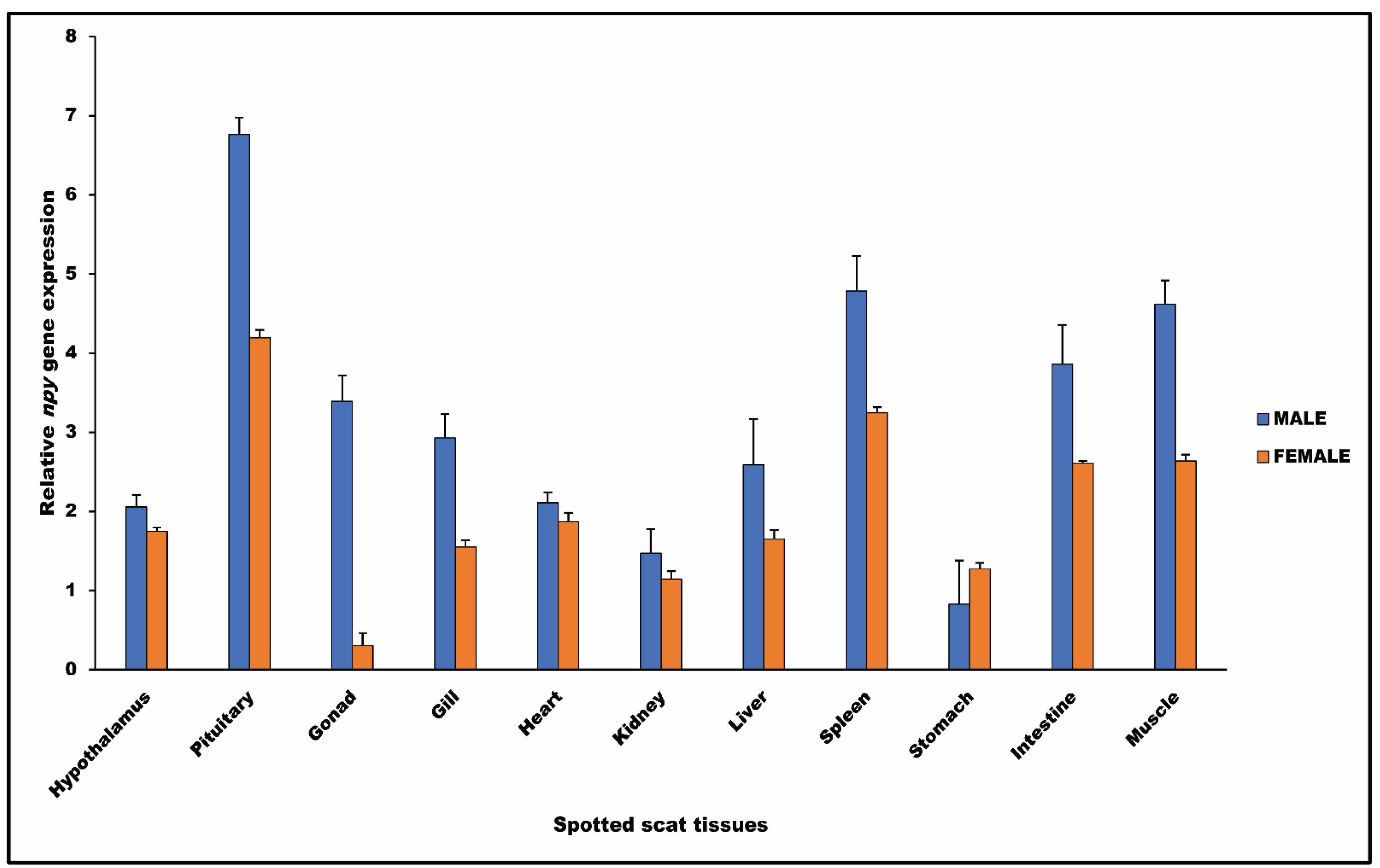
3.3. Npy Gene in Spotted Scat Tissues
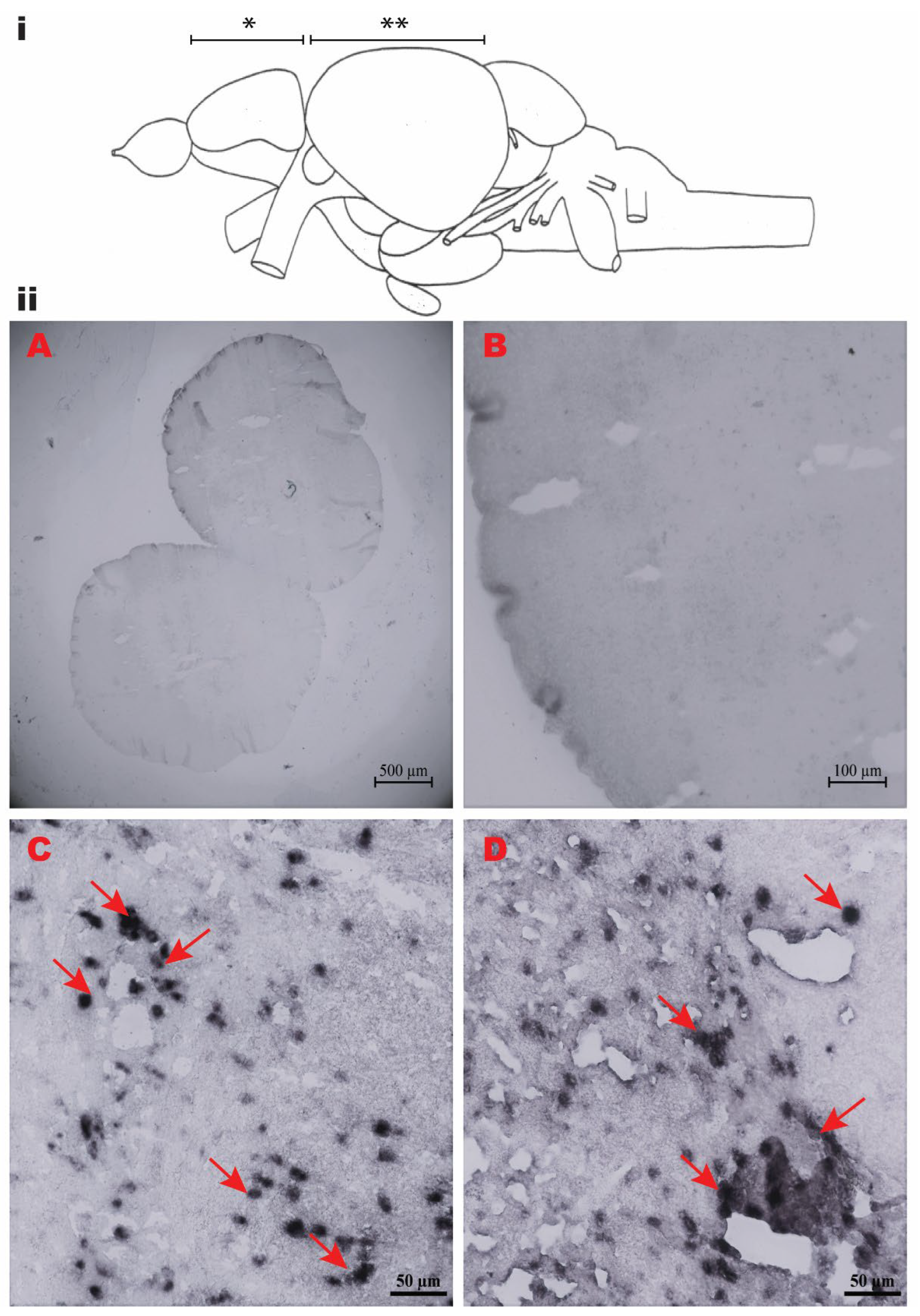
3.4. Localization of Npy mRNA in Spotted Scat Brain
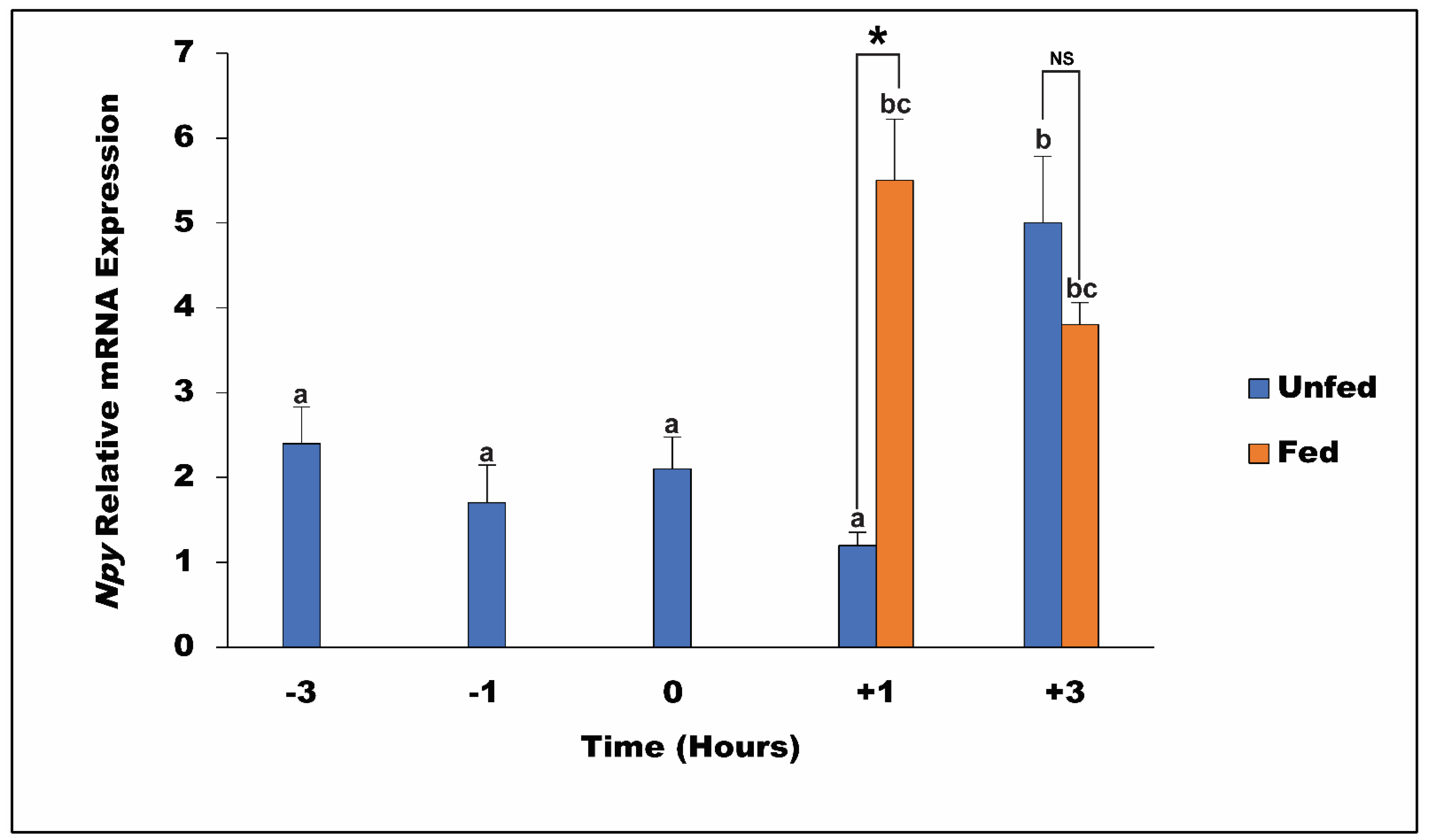
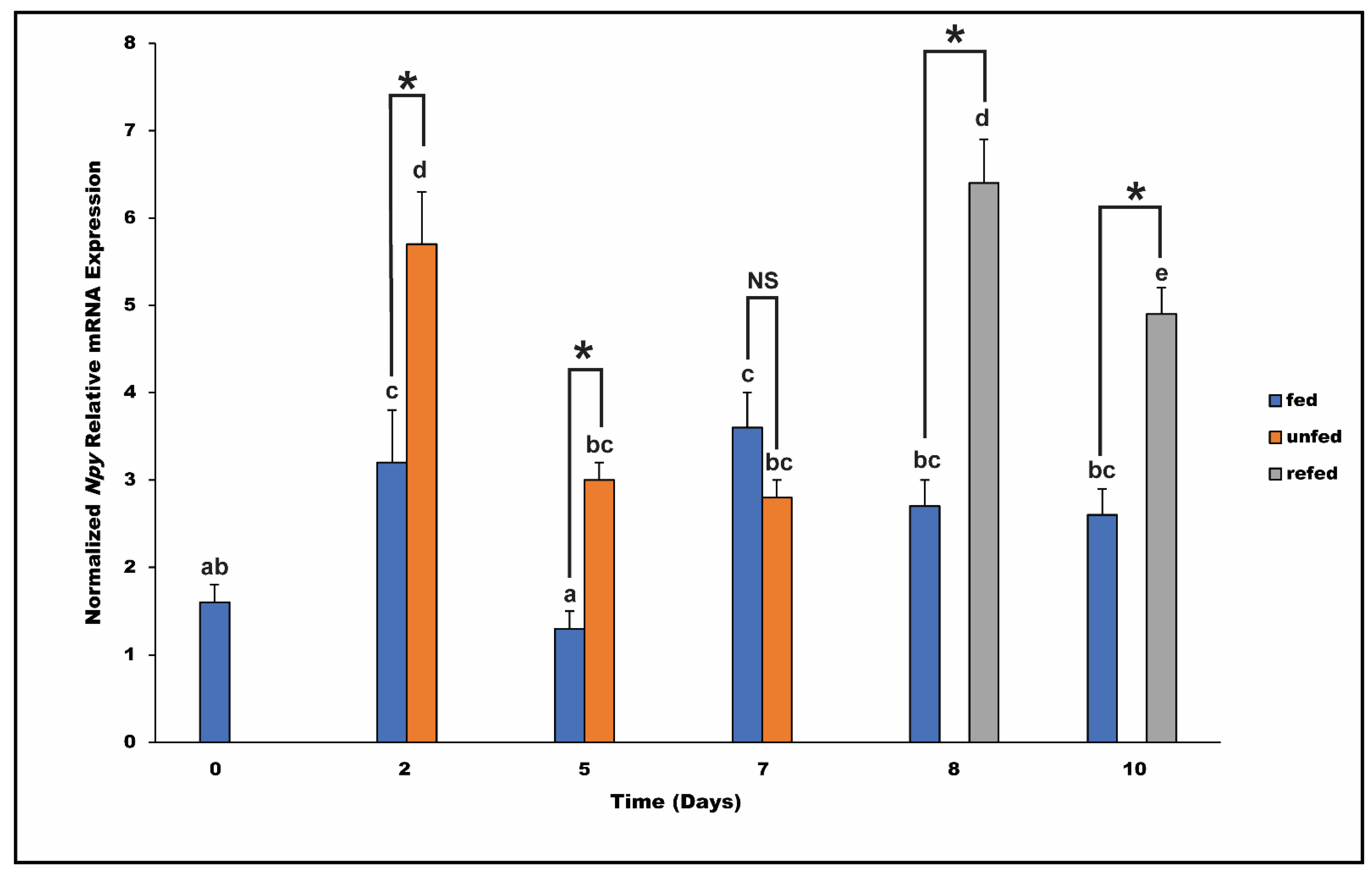
3.5. Pre and Post-Prandial and Long-Term Effects of Feeding, Fasting, and Refeeding on Npy mRNA Level in the Spotted Scat
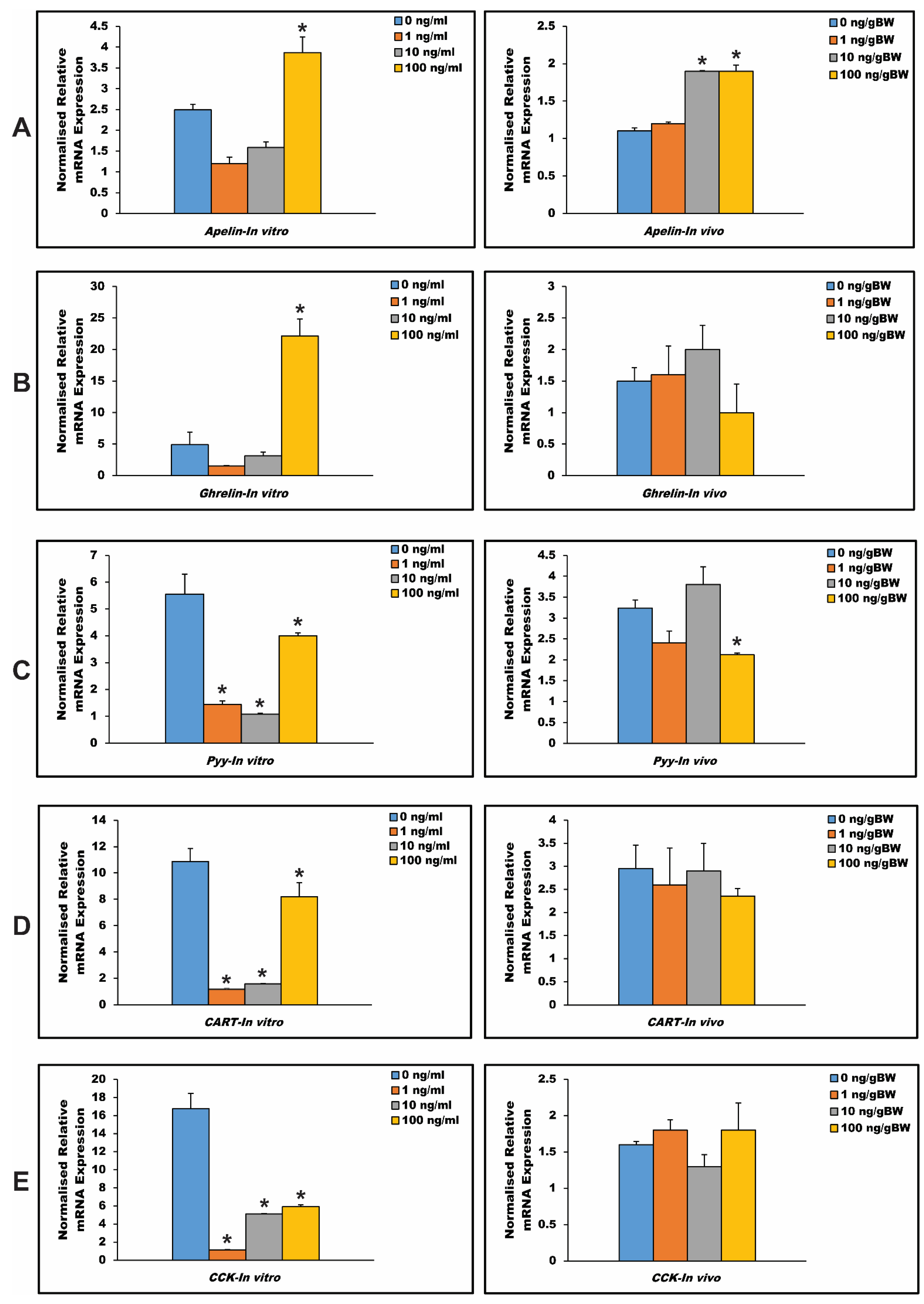
3.6. In Vitro and In Vivo Effects of Npy on Hypothalamus Expression of Other Appetite-Regulating Genes in Spotted Scat
4. Discussion
4.1. Cloning and Sequence Analysis of the Npy Gene
4.2. Npy Gene Expression in Spotted Scat Tissues
4.3. The Npy Gene in Spotted Scat Brain
4.4. Post-Prandial and Fasting and Refeeding Influence on Npy Expression in Spotted Scat Hypothalamus
4.5. The Appetite Regulatory Role of Npy via In Vitro and In Vivo Experimental Studies
4.6. Npy as Growth Control Peptide in Spotted Scat
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Amarasinghe, U.S.; Amarasinghe, M.D.; Nissanka, C. Investigation of the Negombo estuary (Sri Lanka) brush park fishery, with an emphasis on community-based management. Fish. Manag. Ecol. 2002, 9, 41–56. [Google Scholar] [CrossRef]
- Bambaradeniya, C.N.B.; Ekanayake, S.P.; Kekulandala, L.; Fernando, R.; Samarawickrama, V.A.P.; Priyadharshana, T.G.M. An assessment of the status of biodiversity in the Maduganga mangrove estuary. Occas. Pap. IUCN Sri Lanka 2002, 1, 49. [Google Scholar]
- Mathew, A.K. Studies on Some Aspects of Biology of Two Estuarine Fishes Megalops cyprinoides and Scatophagus argus. Ph.D. Thesis, Cochin University of Science and Technology, Kerala, India, 1988. [Google Scholar]
- Morgan, S. Scats: Personable, hardy garbage disposals for the brackish water aquarium. Trop. Fish Hobbyist 1983, 318, 65–69. [Google Scholar]
- Sivan, G.; Venketesvaran, K.; Radhakrishnan, C.K. Biological and biochemical properties of Scatophagus argus venom. Toxicon 2007, 50, 563–571. [Google Scholar] [CrossRef]
- Sivan, G.; Radhakrishnan, C.K. Food, feeding habits and biochemical composition of Scatophagus argus. Turkish J. Fish. Aquat. Sci. 2011, 11, 603–608. [Google Scholar]
- Hajisamae, S.; Yeesin, P.; Chaimongkol, S. Habitat utilization by fishes in a shallow, semi-enclosed estuarine bay in southern Gulf of Thailand. Estuar. Coast. Shelf Sci. 2006, 68, 647–655. [Google Scholar] [CrossRef]
- Gandhi, V. Studies on the food and feeding habits of cultivable butterfish Scatophagus argus (Cuv. and Val.). J. Mar. Biol. Assoc. India 2002, 44, 115–121. [Google Scholar]
- Sivan, G.; Venketasvaran, K.; Radhakrishnan, C.K. Characterization of biological activity of Scatophagus argus venom. Toxicon 2010, 56, 914–925. [Google Scholar] [CrossRef]
- Yoshimura, K.; Yamane, T.; Utsugi, K.; Kohno, H. Seasonal occurrence and abundance of the spotted scat, Scatophagus argus, in surf zones and rivers of. La mer 2003, 41, 82–85. [Google Scholar]
- Gupta, S.; Banerjee, S. Ornamental fish trade in West Bengal. Fish. Chimes 2008, 28, 25–27. [Google Scholar]
- Gupta, S.; Banerjee, S. Indigenous Ornamental Fish Trade of West Bengal; Narendra Publishing House: Delhi, India, 2014; ISBN 9382471383. [Google Scholar]
- Kehoe, A.S.; Volkoff, H. Cloning and characterization of neuropeptide Y (NPY) and cocaine and amphetamine regulated transcript (CART) in Atlantic cod (Gadus morhua). Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2007, 146, 451–461. [Google Scholar] [CrossRef] [PubMed]
- Volkoff, H. The Neuroendocrine Regulation of Food Intake in Fish: A Review of Current Knowledge. Front. Neurosci. 2016, 10, 540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, R.; Zhou, C.; Yuan, D.; Wang, T.; Lin, F.; Chen, H.; Wu, H.; Xin, Z.; Yang, S.; Wang, Y.; et al. Characterization, tissue distribution and regulation of neuropeptideY in Schizothorax prenanti. J. Fish Biol. 2014, 85, 278–291. [Google Scholar] [CrossRef] [PubMed]
- Hoyle, C.H. V Neuropeptide families and their receptors: Evolutionary perspectives. Brain Res. 1999, 848, 1–25. [Google Scholar] [CrossRef]
- Cerdá-Reverter, J.; Larhammar, D. Neuropeptide Y family of peptides: Structure, anatomical expression, function, and molecular evolution. Biochem. Cell Biol. 2000, 78, 371–392. [Google Scholar] [CrossRef]
- Kalra, S.P.; Dube, M.G.; Pu, S.; Xu, B.; Horvath, T.L.; Kalra, P.S. Interacting appetite-regulating pathways in the hypothalamic regulation of body weight. Endocr. Rev. 1999, 20, 68–100. [Google Scholar]
- Halford, J.C.G.; Cooper, G.D.; Dovey, T.M. The Pharmacology of Human Appetite Expression. Curr. Drug Targets 2004, 5, 221–240. [Google Scholar] [CrossRef]
- Sudhakumari, C.-C.; Anitha, A.; Murugananthkumar, R.; Tiwari, D.K.; Bhasker, D.; Senthilkumaran, B.; Dutta-Gupta, A. Cloning, localization and differential expression of Neuropeptide-Y during early brain development and gonadal recrudescence in the catfish, Clarias gariepinus. Gen. Comp. Endocrinol. 2017, 251, 54–65. [Google Scholar] [CrossRef]
- Tatemoto, K. Neuropeptide Y: History and overview. In Neuropeptide Y and Related Peptides; Springer: Berlin/Heidelberg, Germany, 2004; pp. 1–21. [Google Scholar]
- Tatemoto, K.; Carlquist, M.; Mutt, V. Neuropeptide Y—A novel brain peptide with structural similarities to peptide YY and pancreatic polypeptide. Nature 1982, 296, 659–660. [Google Scholar] [CrossRef]
- Assan, D.; Mustapha, U.F.; Chen, H.; Li, Z.; Peng, Y.; Li, G. The Roles of Neuropeptide Y (Npy) and Peptide YY (Pyy) in Teleost Food Intake: A Mini Review. Life 2021, 11, 547. [Google Scholar] [CrossRef]
- Murashita, K.; Kurokawa, T.; Ebbesson, L.O.E.; Stefansson, S.O.; Rønnestad, I. Characterization, tissue distribution, and regulation of agouti-related protein (AgRP), cocaine- and amphetamine-regulated transcript (CART) and neuropeptide Y (NPY) in Atlantic salmon (Salmo salar). Gen. Comp. Endocrinol. 2009, 162, 160–171. [Google Scholar] [CrossRef] [PubMed]
- Narnaware, Y.K.; Peyon, P.P.; Lin, X.; Peter, R.E. Regulation of food intake by neuropeptide Y in goldfish. Am. J. Physiol.—Regul. Integr. Comp. Physiol. 2000, 279, 1025–1034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yokobori, E.; Azuma, M.; Nishiguchi, R.; Kang, K.S.; Kamijo, M.; Uchiyama, M.; Matsuda, K. Neuropeptide Y Stimulates Food Intake in the Zebrafish, Danio rerio. J. Neuroendocrinol. 2012, 24, 766–773. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Liang, X.-F.; Yuan, X.; Li, J.; He, Y.; Fang, L.; Guo, X.; Liu, L.; Li, B.; Shen, D. Neuropeptide Y stimulates food intake and regulates metabolism in grass carp, Ctenopharyngodon idellus. Aquaculture 2013, 380–383, 52–61. [Google Scholar] [CrossRef]
- Kamijo, M.; Kojima, K.; Maruyama, K.; Konno, N.; Motohashi, E.; Ikegami, T.; Uchiyama, M.; Shioda, S.; Ando, H.; Matsuda, K. Neuropeptide Y in Tiger Puffer (Takifugu rubripes): Distribution, Cloning, Characterization, and mRNA Expression Responses to Prandial Condition. Zool. Sci. 2011, 28, 882–890. [Google Scholar] [CrossRef] [PubMed]
- Hosomi, N.; Furutani, T.; Takahashi, N.; Masumoto, T.; Fukada, H. Yellowtail neuropeptide Y: Molecular cloning, tissue distribution, and response to fasting. Fish. Sci. 2014, 80, 483–492. [Google Scholar] [CrossRef]
- Ji, W.; Ping, H.-C.; Wei, K.-J.; Zhang, G.-R.; Shi, Z.-C.; Yang, R.-B.; Zou, G.-W.; Wang, W.-M. Ghrelin, neuropeptide Y (NPY) and cholecystokinin (CCK) in blunt snout bream (Megalobrama amblycephala): cDNA cloning, tissue distribution and mRNA expression changes responding to fasting and refeeding. Gen. Comp. Endocrinol. 2015, 223, 108–119. [Google Scholar] [CrossRef]
- Mercer, R.; Chee, M.; Colmers, W. The role of NPY in hypothalamic mediated food intake. Front. Neuroendocrinol. 2011, 32, 398–415. [Google Scholar] [CrossRef]
- Volkoff, H. The role of neuropeptide Y, orexins, cocaine and amphetamine-related transcript, cholecystokinin, amylin and leptin in the regulation of feeding in fish. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2006, 144, 325–331. [Google Scholar] [CrossRef]
- Mustapha, U.F.; Jiang, D.-N.; Liang, Z.-H.; Gu, H.-T.; Yang, W.; Chen, H.-P.; Deng, S.-P.; Wu, T.-L.; Tian, C.-X.; Zhu, C.-H.; et al. Male-specific Dmrt1 is a candidate sex determination gene in spotted scat (Scatophagus argus). Aquaculture 2018, 495, 351–358. [Google Scholar] [CrossRef]
- Carpio, Y.; Acosta, J.; Morales, A.; Herrera, F.; González, L.J.; Estrada, M.P. Cloning, expression and growth promoting action of Red tilapia (Oreochromis sp.) neuropeptide Y. Peptides 2006, 27, 710–718. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Wen, Z.Y.; Zou, Y.C.; Qin, C.J.; Wang, J.; Yuan, D.Y.; Li, R. Molecular cloning, tissue distribution, and effect of fasting and refeeding on the expression of neuropeptide Y in Channa argus. Gen. Comp. Endocrinol. 2018, 259, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Lei, L.; Yuan, D.; Zheng, Z.; Zhu, C.; Luo, H.; Ye, H.; Li, D.; Wang, J.; Li, B.; et al. Cloning, expression profiling, and effects of fasting status on neuropeptide Y in Schizothorax davidi. J. Food Biochem. 2019, 43, e12892. [Google Scholar] [CrossRef] [PubMed]
- Campos, V.F.; Collares, T.; Deschamps, J.C.; Seixas, F.K.; Dellagostin, O.A.; Lanes, C.F.C.; Sandrini, J.; Marins, L.F.; Okamoto, M.; Sampaio, L.A. Identification, tissue distribution and evaluation of brain neuropeptide Y gene expression in the Brazilian flounder Paralichthys orbignyanus. J. Biosci. 2010, 35, 405–413. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Li, W.; Lin, H. cDNA cloning and mRNA expression of neuropeptide Y in orange spotted grouper, Epinephelus coioides. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2005, 142, 79–89. [Google Scholar] [CrossRef]
- Liang, X.-F.; Li, G.-Z.; Yao, W.; Cheong, L.-W.; Liao, W.-Q. Molecular characterization of neuropeptide Y gene in Chinese perch, an acanthomorph fish. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2007, 148, 55–64. [Google Scholar] [CrossRef]
- Silverstein, J.T.; Breininger, J.; Baskin, D.G.; Plisetskaya, E.M. Neuropeptide Y-like gene expression in the salmon brain increases with fasting. Gen. Comp. Endocrinol. 1998, 110, 157–165. [Google Scholar] [CrossRef]
- Montpetit, C.J.; Chatalov, V.; Yuk, J.; Rasaratnam, I.; Youson, J.H. Expression of neuropeptide Y family peptides in the brain and gut during stages of the life cycle of a parasitic lamprey (Petromyzon marinus) and a nonparasitic lamprey (Ichthyomyzon gagei). Ann. N. Y. Acad. Sci. 2005, 1040, 140–149. [Google Scholar] [CrossRef]
- Leonard, J.B.K.; Waldbieser, G.C.; Silverstein, J.T. Neuropeptide Y Sequence and Messenger RNA Distribution in Channel Catfish (Ictalurus punctatus). Mar. Biotechnol. 2001, 3, 111–118. [Google Scholar] [CrossRef]
- Peng, C.; Gallin, W.; Peter, R.E.; Blomqvist, A.G.; Larhammar, D. Neuropeptide-Y gene expression in the goldfish brain: Distribution and regulation by ovarian steroids. Endocrinology 1994, 134, 1095–1103. [Google Scholar] [CrossRef]
- Gaikwad, A.; Biju, K.C.; Muthal, P.L.; Saha, S.; Subhedar, N. Role of neuropeptide Y in the regulation of gonadotropin releasing hormone system in the forebrain of Clarias batrachus (Linn.): Immunocytochemistry and high performance liquid chromatography-electrospray ionization-mass spectrometric analysis. Neuroscience 2005, 133, 267–279. [Google Scholar] [CrossRef] [PubMed]
- Sundström, G.; Larsson, T.A.; Brenner, S.; Venkatesh, B.; Larhammar, D.A.N. Ray-Fin Fish Tetraploidization Gave Rise to Pufferfish Duplicates of NPY and PYY, but Zebrafish NPY Duplicate Was Lost. Ann. N. Y. Acad. Sci. 2005, 1040, 476–478. [Google Scholar] [CrossRef] [PubMed]
- Xiang, H. Comparative aspects of the role of neuropeptide Y in the regulation of the vertebrate heart. Cardioscience 1994, 5, 209–213. [Google Scholar] [PubMed]
- Lei, L.; Deng, X.; Yuan, D.; Zheng, Z.; Zhu, C.; Luo, H.; Li, B.; Ye, H.; Zhou, C. Molecular Cloning and Function Characterization in Feeding of Neuropeptide Y in Gibel Carp, Carassius auratus gibelio. Pak. J. Zool. 2019, 51, 1017–1025. [Google Scholar] [CrossRef]
- Cerdá-Reverter, J.M.; Anglade, I.; Martínez-Rodríguez, G.; Mazurais, D.; Muñoz-Cueto, J.A.; Carrillo, M.; Kah, O.; Zanuy, S. Characterization of neuropeptide Y expression in the brain of a perciform fish, the sea bass (Dicentrarchus labrax). J. Chem. Neuroanat. 2000, 19, 197–210. [Google Scholar] [CrossRef]
- Gaikwad, A.; Biju, K.C.; Saha, S.G.; Subhedar, N. Neuropeptide Y in the olfactory system, forebrain and pituitary of the teleost, Clarias batrachus. J. Chem. Neuroanat. 2004, 27, 55–70. [Google Scholar] [CrossRef]
- Narnaware, Y.K.; Peter, R.E. Effects of food deprivation and refeeding on neuropeptide Y (NPY) mRNA levels in goldfish. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2001, 129, 633–637. [Google Scholar] [CrossRef]
- Volkoff, H.; Canosa, L.F.; Unniappan, S.; Cerdá-Reverter, J.M.; Bernier, N.J.; Kelly, S.P.; Peter, R.E. Neuropeptides and the control of food intake in fish. Gen. Comp. Endocrinol. 2005, 142, 3–19. [Google Scholar] [CrossRef]
- Chen, Y.; Shen, Y.; Pandit, N.P.; Fu, J.; Li, D.; Li, J. Molecular cloning, expression analysis, and potential food intake attenuation effect of peptide YY in grass carp (Ctenopharyngodon idellus). Gen. Comp. Endocrinol. 2013, 187, 66–73. [Google Scholar] [CrossRef]
- Gonzalez, R.; Unniappan, S. Molecular characterization, appetite regulatory effects and feeding related changes of peptide YY in goldfish. Gen. Comp. Endocrinol. 2010, 166, 273–279. [Google Scholar] [CrossRef]
- Valen, R.; Jordal, A.-E.; Murashita, K.; Rønnestad, I. Postprandial effects on appetite-related neuropeptide expression in the brain of Atlantic salmon, Salmo salar. Gen. Comp. Endocrinol. 2011, 171, 359–366. [Google Scholar] [CrossRef] [PubMed]
- Boujard, T.; Leatherland, J.F. Circadian rhythms and feeding time in fishes. Environ. Biol. Fishes 1992, 35, 109–131. [Google Scholar] [CrossRef]
- Silverstein, J.T.; Plisetskaya, E.M. The effects of NPY and insulin on food intake regulation in fish. Am. Zool. 2000, 40, 296–308. [Google Scholar] [CrossRef]
- Rønnestad, I.; Gomes, A.S.; Murashita, K.; Angotzi, R.; Jönsson, E.; Volkoff, H. Appetite-Controlling Endocrine Systems in Teleosts. Front. Endocrinol. 2017, 8, 73. [Google Scholar] [CrossRef] [Green Version]
- Blanco, A.M.; Bertucci, J.I.; Valenciano, A.I.; Delgado, M.J.; Unniappan, S. Ghrelin suppresses cholecystokinin (CCK), peptide YY (PYY) and glucagon-like peptide-1 (GLP-1) in the intestine, and attenuates the anorectic effects of CCK, PYY and GLP-1 in goldfish (Carassius auratus). Horm. Behav. 2017, 93, 62–71. [Google Scholar] [CrossRef]
- Miura, T.; Maruyama, K.; Shimakura, S.-I.; Kaiya, H.; Uchiyama, M.; Kangawa, K.; Shioda, S.; Matsuda, K. Regulation of food intake in the goldfish by interaction between ghrelin and orexin. Peptides 2007, 28, 1207–1213. [Google Scholar] [CrossRef]
- Yan, X.; Qin, C.; Yang, G.; Deng, D.; Yang, L.; Feng, J.; Mi, J.; Nie, G. The Regulatory Role of Apelin on the Appetite and Growth of Common Carp (Cyprinus carpio L.). Animals 2020, 10, 2163. [Google Scholar] [CrossRef]
- Penney, C.C.; Volkoff, H. Peripheral injections of cholecystokinin, apelin, ghrelin and orexin in cavefish (Astyanax fasciatus mexicanus): Effects on feeding and on the brain expression levels of tyrosine hydroxylase, mechanistic target of rapamycin and appetite-related hormones. Gen. Comp. Endocrinol. 2014, 196, 34–40. [Google Scholar] [CrossRef]
- Matsuda, K. Recent Advances in the Regulation of Feeding Behavior by Neuropeptides in Fish. Ann. N. Y. Acad. Sci. 2009, 1163, 241–250. [Google Scholar] [CrossRef]
- Volkoff, H.; Wyatt, J.L. Apelin in goldfish (Carassius auratus): Cloning, distribution and role in appetite regulation. Peptides 2009, 30, 1434–1440. [Google Scholar] [CrossRef]
- Yan, X.; Qin, C.; Deng, D.; Yang, G.; Feng, J.; Lu, R.; Yang, L.; Meng, X.; Cao, X.; Nie, G. Apelin and apj in common carp (Cyprinus carpio L.): Molecular characterization, tissue expression and responses to periprandial changes and fasting–refeeding. Aquac. Res. 2020, 51, 1012–1025. [Google Scholar] [CrossRef]
- Lee, D.K.; Cheng, R.; Nguyen, T.; Fan, T.; Kariyawasam, A.P.; Liu, Y.; Osmond, D.H.; George, S.R.; O’Dowd, B.F. Characterization of apelin, the ligand for the APJ receptor. J. Neurochem. 2000, 74, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Volkoff, H. In vitro assessment of interactions between appetite-regulating peptides in brain of goldfish (Carassius auratus). Peptides 2014, 61, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Diz-Chaves, Y. Ghrelin, appetite regulation, and food reward: Interaction with chronic stress. Int. J. Pept. 2011, 2011, 898450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Volkoff, H.; Peter, R.E. Effects of CART peptides on food consumption, feeding and associated behaviors in the goldfish, Carassius auratus: Actions on neuropeptide Y- and orexin A-induced feeding. Brain Res. 2000, 887, 125–133. [Google Scholar] [CrossRef]
- Volkoff, H.; Joy Eykelbosh, A.; Ector Peter, R. Role of leptin in the control of feeding of goldfish Carassius auratus: Interactions with cholecystokinin, neuropeptide Y and orexin A, and modulation by fasting. Brain Res. 2003, 972, 90–109. [Google Scholar] [CrossRef]
- Batterham, R.L.; Cowley, M.A.; Small, C.J.; Herzog, H.; Cohen, M.A.; Dakin, C.L.; Wren, A.M.; Brynes, A.E.; Low, M.J.; Ghatei, M.A. Gut hormone PYY 3-36 physiologically inhibits food intake. Nature 2002, 418, 650–654. [Google Scholar] [CrossRef]
- Kuz’mina, V.V. The Regulatory Mechanisms of Feeding Behavior in Fish. J. Evol. Biochem. Physiol. 2019, 55, 1–13. [Google Scholar] [CrossRef]
- Volkoff, H.; Hoskins, L.J.; Tuziak, S.M. Influence of intrinsic signals and environmental cues on the endocrine control of feeding in fish: Potential application in aquaculture. Gen. Comp. Endocrinol. 2010, 167, 352–359. [Google Scholar] [CrossRef]
- Bertucci, J.I.; Blanco, A.M.; Sundarrajan, L.; Rajeswari, J.J.; Velasco, C.; Unniappan, S. Nutrient regulation of endocrine factors influencing feeding and growth in fish. Front. Endocrinol. 2019, 10, 83. [Google Scholar] [CrossRef] [Green Version]
- Conde-Sieira, M.; Soengas, J.L. Nutrient Sensing Systems in Fish: Impact on Food Intake Regulation and Energy Homeostasis. Front. Neurosci. 2017, 10, 603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peter, R.E.; Chang, J.P. Brain regulation of growth hormone secretion and food intake in fish. In Neural Regulation in the Vertebrate Endocrine System; Springer: Berlin/Heidelberg, Germany, 1999; pp. 55–67. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene and Primer Orientation | Primers Sequence (5′–3′) | Application |
|---|---|---|
| Npy-F Npy-R | GCAAACCTCAGCCTAACCCTCT ACTGTAAGCAGTTGTGACTGTAGC | ORF/ cloning |
| Npy-TD-F1 Npy-TD-R1 | ATACCCGGTGAAACCCGAGAAC GCGTGTCTGTGCTTTCCTTCAA | Tissue Distribution (Gene expression) |
| Npy-ish-F1 Npy-ish-R1 | taatacgactcactatagggATACCCGGTGAAACCCGAGAAC taatacgactcactatagggGCGTGTCTGTGCTTTCCTTCAA | In situ hybridization |
| β -Actin-F β -Actin-R | GAGAGGTTCCGTTGCCCAGAG ACGCCAACACTGTGCTGTCTG | House-keeping gene |
| Apelin-F Apelin-R | ATGAATGTGAAGATCCTGACG CTAGAACGGCATGGGTCCC | Npy in vitro and in vivo experiment analysis on other appetite-regulating genes |
| Ghrelin-F Ghrelin-R | CTCAGCCCTTCACAGAAACCTC ATCTCCTGCAACGCCACATCGT | |
| Pyy-F Pyy-R | GGAACACTGGCAGATGCCTACC TCTGTTGCTGTCGCCACCAA | |
| Cart-F Cart-R | CAGCGCCGCTGGAGCCCAAGTGT TTCTTTTCCCACAGAGGCAGTC | |
| Cck-F Cck-R | TGCTCGAAACAGCCTGAACCAG AAAGTCCATCCAGCCGAGGTAA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Assan, D.; Wang, Y.; Mustapha, U.F.; Ndandala, C.B.; Li, Z.; Li, G.-L.; Chen, H. Neuropeptide Y in Spotted Scat (Scatophagus Argus), Characterization and Functional Analysis towards Feed Intake Regulation. Fishes 2022, 7, 111. https://doi.org/10.3390/fishes7030111
Assan D, Wang Y, Mustapha UF, Ndandala CB, Li Z, Li G-L, Chen H. Neuropeptide Y in Spotted Scat (Scatophagus Argus), Characterization and Functional Analysis towards Feed Intake Regulation. Fishes. 2022; 7(3):111. https://doi.org/10.3390/fishes7030111
Chicago/Turabian StyleAssan, Daniel, Yaorong Wang, Umar Farouk Mustapha, Charles Brighton Ndandala, Zhiyuan Li, Guang-Li Li, and Huapu Chen. 2022. "Neuropeptide Y in Spotted Scat (Scatophagus Argus), Characterization and Functional Analysis towards Feed Intake Regulation" Fishes 7, no. 3: 111. https://doi.org/10.3390/fishes7030111
APA StyleAssan, D., Wang, Y., Mustapha, U. F., Ndandala, C. B., Li, Z., Li, G.-L., & Chen, H. (2022). Neuropeptide Y in Spotted Scat (Scatophagus Argus), Characterization and Functional Analysis towards Feed Intake Regulation. Fishes, 7(3), 111. https://doi.org/10.3390/fishes7030111