
Comparative Analysis of Intestinal Characteristics of Largemouth Bass (Micropterus salmoides) and Intestinal Flora with Different Growth Rates

Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Sample Collection
2.3. Intestinal Histological Analysis
2.4. Enzyme Activity Analysis
2.5. Total DNA Extraction and Biological Analysis of Intestinal Flora
2.6. Data Collection and Statistical Analysis
3. Results
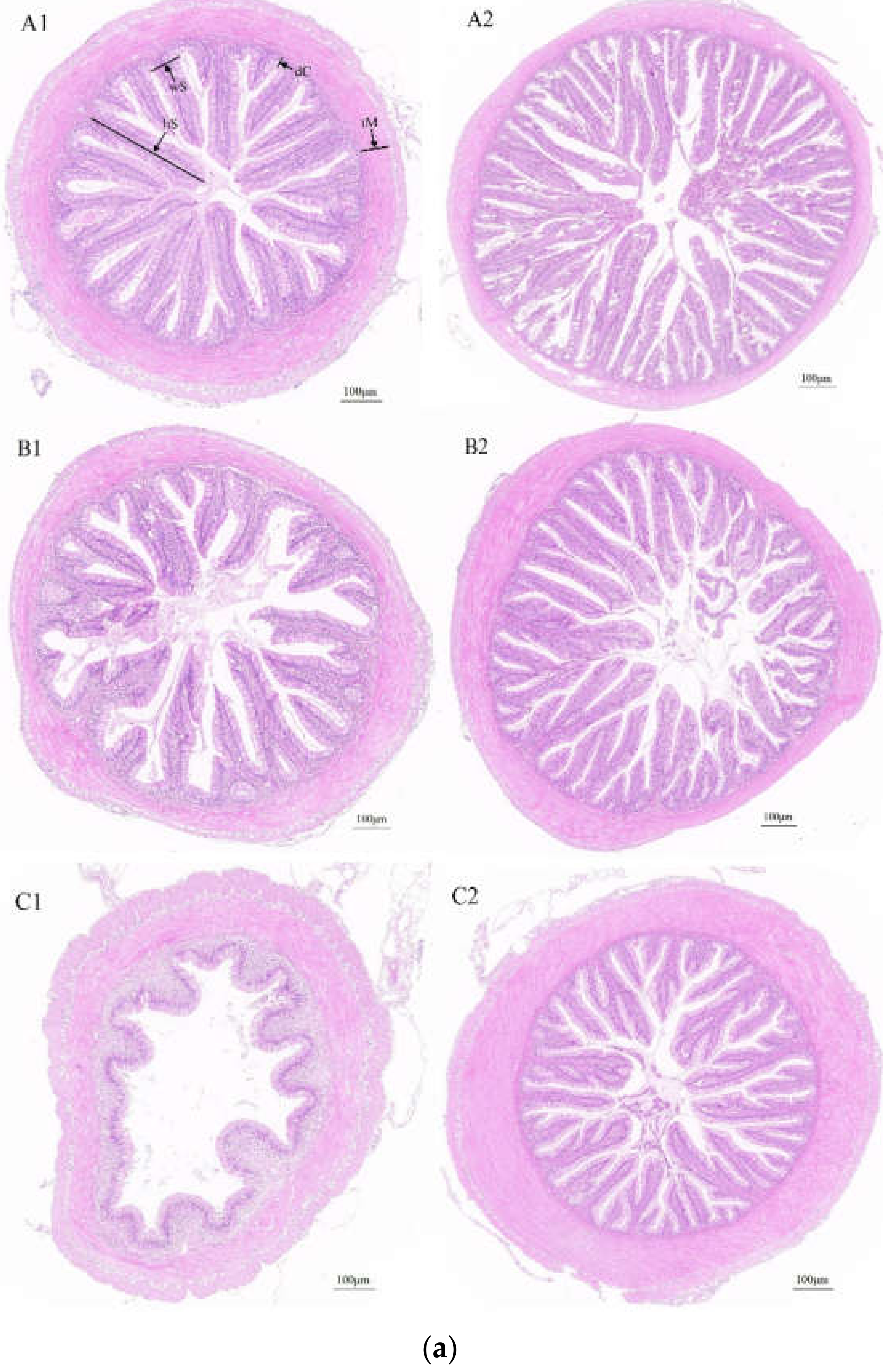
3.1. Intestinal Histomorphology of Juvenile Largemouth Bass with Different Growth Rates
3.1.1. Morphological Characteristics of the Foregut, Midgut and Hindgut
3.1.2. Determination of Morphological Indexes of the Foregut, Midgut and Hindgut
3.2. Intestinal Digestive Enzyme Activities of Juvenile Largemouth Bass with Different Growth Rates
3.3. Intestinal Microfloral Structure of Largemouth Bass with Different Growth Rates
3.3.1. Sequence Characteristics of the Intestinal Flora
3.3.2. Assessment of Intestinal Floral Differences
4. Discussion
4.1. Distinct Gut Characteristics of the Fast-Growing Group of Largemouth Bass
4.2. Differences in Intestinal Floral Levels between the FG Group and SG Group
4.3. Differences in Intestinal Flora at the Genus Level between the FG and SG Groups
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Adamek, D.; Rzepkowska, M.; Panagiotopoulou, H.; Ostaszewska, T.; Kolman, R. Morphological Differences of White Muscle Fibers and Genetic Diversity of Fast and Slow Growing Atlantic Sturgeons (Acipenser oxyrinchus). Turk. J. Fish. Aquat. Sci. 2017, 17, 959–966. [Google Scholar] [CrossRef]
- Mun, S.H.; You, J.H.; Oh, H.J.; Lee, C.H.; Baek, H.J.; Lee, Y.D.; Kwon, J.Y. Expression Patterns of Growth Related Genes in Juvenile Red Spotted Grouper (Epinephelus akaara) with Different Growth Performance after Size Grading. Dev. Reprod. 2019, 23, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Alm, G. Connection between Maturity, Size and Age in Fishes; Institute of Freshwater Research Report; Institute of Freshwater Research: Stockholm, Sweden, 1959. [Google Scholar]
- Tamayo, D.; Ibarrola, I.; Urrutia, M.B.; Navarro, E. The Physiological Basis for Inter-Individual Growth Variability in the Spat of Clams (Ruditapes philippinarum). Aquaculture 2011, 321, 113–120. [Google Scholar] [CrossRef]
- Valente, L.M.P.; Moutou, K.A.; Conceicao, L.E.C. What Determines Growth Potential and Juvenile Quality of Farmed Fish Species? Rev. Aquac. 2013, 5, S168–S193. [Google Scholar] [CrossRef]
- Xie, F.; Ai, Q.; Mai, K.; Wei, X.; Ma, H. The Optimal Feeding Frequency of Large Yellow Croaker (Pseudosciaena Crocea, Richardson) Larvae. Aquaculture 2011, 311, 162–167. [Google Scholar] [CrossRef]
- Wang, C.L.; Wang, X.D.; Xiao, S.S.; Bu, X.Y.; Chen, L.Q. T-2 Toxin in the Diet Suppresses Growth and Induces Immunotoxicity in Juvenile Chinese Mitten Crab (Eriocheir sinensis). Fish Shellfish Immunol. 2019, 97, 593–601. [Google Scholar] [CrossRef] [PubMed]
- Escaffre, A.M.; Kaushik, S.; Mambrini, M. Morphometric Evaluation of Changes in the Digestive Tract of Rainbow Trout (Oncorhynchus mykiss) due to Fish Meal Replacement with Soy Protein Concentrate. Aquaculture 2007, 273, 127–138. [Google Scholar] [CrossRef]
- Noaillac-Depeyre, J.; Gas, N. Structure and Function of the Intestinal Epithelial Cells in the Perch (Perca Fluviatillis L.). Anat. Rec. 1979, 195, 621–627. [Google Scholar] [CrossRef]
- Kashinskaya, E.N.; Suhanova, E.V.; Solov’Ev, M.M.; Izvekova, G.I.; Glupov, V.V. Diversity of Microbial Communities of the Intestinal Mucosa and Intestinal Contents of Fish from Lake Chany (Western Siberia). Inland Water Biol. 2014, 7, 172–177. [Google Scholar] [CrossRef]
- Comabella, Y.; Mendoza, R.; Aguilera, C.; Carrillo, O.; Hurtado, A.; García-Galano, T. Digestive Enzyme Activity During Early Larval Development of the Cuban Gar Atractosteus tristoechus. Fish Physiol. Biochem. 2006, 32, 147–157. [Google Scholar] [CrossRef]
- Kolkovski, S.; Tandler, A.; Kissil, G.W.; Gertler, A. The Effect of Dietary Exogenous Digestive Enzymes on Ingestion, Assimilation, Growth and Survival of Gilthead Seabream (Sparus Aurata, Sparidae, Linnaeus) Larvae. Fish Physiol. Biochem. 1993, 12, 203–209. [Google Scholar] [CrossRef]
- Du, Z.-Y.; Liu, Y.-J.; Tian, L.-X.; Wang, J.-T.; Wang, Y.; Liang, G.-Y. Effect of Dietary Lipid Level on Growth, Feed Utilization and Body Composition by Juvenile Grass Carp (Ctenopharyngodon idella). Aquac. Nutr. 2005, 11, 139–146. [Google Scholar] [CrossRef]
- Pei, Z.; Xie, S.; Lei, W.; Zhu, X.; Yang, Y.; Pei, Z.; Xie, S.; Lei, W.; Zhu, X.; Yang, Y. Comparative Study on the Effect of Dietary Lipid Level on Growth and Feed Utilization for Gibel Carp (Carassius auratus gibelio) and Chinese Longsnout Catfish (Leiocassis longirostris Gunther). Aquac. Nutr. 2015, 10, 209–216. [Google Scholar] [CrossRef]
- Chapagain, P.; Arivett, B.; Cleveland, B.M.; Walker, D.M.; Salem, M. Analysis of the Fecal Microbiota of Fast- and Slow-Growing Rainbow Trout (Oncorhynchus mykiss). BMC Genom. 2019, 20, 788. [Google Scholar] [CrossRef]
- Shi, Y.; Ma, D.Y.; Wei, Z.S. Revealing the Difference of Intestinal Microbiota Composition of Cultured European Eels (Anguilla anguilla) with Different Growth Rates. Isr. J. Aquac. 2020, 72, 4–12. [Google Scholar] [CrossRef]
- Guan, F.; Shen, L.; Zhou, X.; Chen, Z.; Yu, C.; Zhang, J.; Yuan, Y. Effects of Underwater and Semi-Aquatic Environments on Gut Tissue and Microbiota of the Mudskipper Boleophthalmus Pectinirostris. J. Comp. Physiol. B 2021, 191, 741–753. [Google Scholar] [CrossRef]
- Li, Y.Y.; Chen, X.; Li, S.Y.; Li, P.; Li, Y.H.; Song, T.Y. Influence of factors related to the intestinal digestion and absorption on growth of cultured large yellow croaker Pseudosciaena crocea. J. Dalian Ocean. Univ. 2015, 30, 5. [Google Scholar]
- Bayne, B.L. Physiological Components of Growth Differences between Individual Oysters (Crassostrea gigas) and a Comparison with Saccostrea Commercialis. Physiol. Biochem. Zool. 1999, 72, 705–713. [Google Scholar] [CrossRef]
- Ibarrola, I.; Hilton, Z.; Ragg, N. Physiological Basis of Inter-Population, Inter-Familiar and Intra-Familiar Differences in Growth Rate in the Green-Lipped Mussel Perna Canaliculus. Aquaculture 2017, 479, 544–555. [Google Scholar] [CrossRef]
- Adamek-Urbańska, D.; Kasprzak, R.; Tyszkiewicz, M.; Fisher, K.; Dbrowski, K. Negative Effects of Artificial Diets on Growth and the Digestive Tract of 1-month-old Redhead Cichlid (Vieja melanura, Günther, 1862). Aquac. Res. 2021, 52, 4890–4896. [Google Scholar] [CrossRef]
- Goodlad, R.A.; Ratcliffe, B.; Lee, C.Y.; Wright, N.A. Dietary Fibre and the Gastrointestinal Tract: Differing Trophic Effects on Muscle and Mucosa of the Stomach, Small Intestine and Colon. Eur. J. Clin. Nutr. 1995, 49 (Suppl. S3), S178–S181. [Google Scholar] [PubMed]
- Austin, B. The Bacterial Microflora of Fish. Sci. World J. 2014, 2, 558–572. [Google Scholar] [CrossRef][Green Version]
- Wang, A.R.; Ran, C.; Ringø, E.; Zhou, Z.G. Progress in Fish Gastrointestinal Microbiota Research. Rev. Aquac. 2018, 10, 1753–5123. [Google Scholar] [CrossRef]
- Moon, C.D.; Wayne, Y.; Maclean, P.H.; Cookson, A.L.; Bermingham, E.N. Metagenomic Insights into the Roles of Proteobacteria in the Gastrointestinal Microbiomes of Healthy Dogs and Cats. MicrobiologyOpen 2018, 7, e00677. [Google Scholar] [CrossRef] [PubMed]
- Bereded, N.K.; Abebe, G.B.; Fanta, S.W.; Curto, M.; Domig, K.J. The Impact of Sampling Season and Catching Site (Wild and Aquaculture) on Gut Microbiota Composition and Diversity of Nile Tilapia (Oreochromis niloticus). Biology 2021, 10, 180. [Google Scholar] [CrossRef]
- Semova, I.; Carten, J.; Stombaugh, J.; Mackey, L.; Knight, R.; Farber, S.; Rawls, J.F. Microbiota Regulate Intestinal Absorption and Metabolism of Fatty Acids in the Zebrafish. Cell Host Microbe 2012, 12, 277–288. [Google Scholar] [CrossRef] [PubMed]
- Anandan, R.; Dhanasekaran, D.; Manogaran, G.P. An Introduction to Actinobacteria. In Actinobacteria—Basics and Biotechnological Applications; IntechOpen: London, UK, 2016; pp. 4–37. [Google Scholar]
- Fan, L.; Li, Q.X. Characteristics of Intestinal Microbiota in the Pacific White Shrimp Litopenaeus vannamei Differing Growth Performances in the Marine Cultured Environment. Aquaculture 2019, 50, 450–461. [Google Scholar] [CrossRef]
- Tan, X.; Sun, Z. Dietary Dandelion Extract Improved Growth Performance, Immunity, Intestinal Morphology and Microbiota Composition of Golden Pompano Trachinotus ovatus. Aquac. Rep. 2020, 18, 100491. [Google Scholar] [CrossRef]
- Kostic, A.D.; Gevers, D.; Pedamallu, C.S.; Michaud, M.; Meyerson, M. Genomic Analysis Identifies Association of Fusobacterium with Colorectal Carcinoma. Genome Res. 2012, 22, 292–298. [Google Scholar] [CrossRef]
- Bachrach, G.; Ianculovici, C.; Naor, R.; Weiss, E.I. Fluorescence Based Measurements of Fusobacterium Nucleatum Coaggregation and of Fusobacterial Attachment to Mammalian Cells. FEMS Microbiol. Lett. 2010, 248, 235–240. [Google Scholar] [CrossRef]
- Li, Y.D.; Zhou, F.L.; Tang, Y.P.; Huang, J.H.; Jiang, S.G. Variation in Bacterial Communities among Stress: Ensitive and Stress-Tolerant Black Tiger Shrimp (Penaeus monodon) Individuals. Aquac. Res. 2020, 52, 2146–2159. [Google Scholar] [CrossRef]
- Didier, R. Probiotics and Obesity: A Link? Nat. Rev. Microbiol. 2009, 7, 616. [Google Scholar]
- Itami, T.; Asano, M.; Tokushige, K.; Kubono, K.; Nakagawa, A.; Takeno, N.; Nishimura, H.; Maeda, M.; Kondo, M.; Takahashi, Y. Enhancement of Disease Resistance of Kuruma Shrimp, Penaeus Japonicus, after Oral Administration of Peptidoglycan Derived from Bifidobacterium Thermophilum. Aquaculture 1998, 164, 277–288. [Google Scholar] [CrossRef]
- Hsu, H.-Y.; Chang, F.-C.; Wang, Y.-B.; Chen, S.-H.; Lin, Y.-P.; Chung, Y.-L.; Han, Y.-S. Revealing the Compositions of the Intestinal Microbiota of Three Anguillid Eel Species Using 16S rDNA Sequencing. Aquac. Res. 2018, 49, 2404–2415. [Google Scholar] [CrossRef]
- Mao, L.; Zeng, C.X.; Jia, X.Q.; Zhai, S.W.; Li, Z.Q.; Ma, Y. The Composition and Structure of the Intestinal Microflora of Anguilla Marmorata at Different Growth Rates: A Deep Sequencing Study. J. Appl. Bacteriol. 2018, 126, 1340–1352. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Fold Length/μm | Fold Width/μm | Crypt Depth/μm | Goblet Cell Number/Cells/100 μm | Muscle Thickness/μm |
|---|---|---|---|---|---|
| Foregut (SG) | 243.097 ± 32.021 A | 88.822 ± 16.607 A | 25.216 ± 1.489 | 7.846 ± 1.461 A | 70.897 ± 2.624 A |
| Foregut (FG) | 711.945 ± 143.179 B | 121.795 ± 30.937 B | 24.501 ± 4.173 | 10.230 ± 2.888 B | 104.589 ± 14.392 B |
| Midgut (SG) | 262.311 ± 44.298 A | 99.601 ± 24.127 | 23.170 ± 4.491 | 7.851 ± 2.582 | 66.581 ± 17.173 A |
| Midgut (FG) | 479.917 ± 123.664 B | 95.035 ± 23.205 | 23.976 ± 4.426 | 8.481 ± 2.190 | 115.418 ± 42.527 B |
| Hindgut (SG) | 166.811 ± 36.217 A | 175.475 ± 16.175 A | 40.821 ± 5.996 A | 10.166 ± 3.714 A | 101.991 ± 14.217 A |
| Hindgut (FG) | 422.190 ± 87.901 B | 93.080 ± 20.008 B | 25.139 ± 4.500 B | 13.222 ± 2.414 B | 189.381 ± 43.220 B |
| Group | Amylase Activity/(U·g−1) | Lipase Activity/(U·g−1) | Trypsin Activity/(U·g−1) |
|---|---|---|---|
| Slow-growing group (SG) | 0.289 ± 0.005 | 9.756 ± 0.122 A | 2487.939 ± 325.567 |
| Fast-growing (FG) | 0.287 ± 0.011 | 13.369 ± 0.224 B | 2849.089 ± 137.591 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Su, X.; Ji, D.; Yao, J.; Zou, Y.; Yan, M. Comparative Analysis of Intestinal Characteristics of Largemouth Bass (Micropterus salmoides) and Intestinal Flora with Different Growth Rates. Fishes 2022, 7, 65. https://doi.org/10.3390/fishes7020065
Su X, Ji D, Yao J, Zou Y, Yan M. Comparative Analysis of Intestinal Characteristics of Largemouth Bass (Micropterus salmoides) and Intestinal Flora with Different Growth Rates. Fishes. 2022; 7(2):65. https://doi.org/10.3390/fishes7020065
Chicago/Turabian StyleSu, Xin, Da Ji, Junjie Yao, Yuanlong Zou, and Mengzhi Yan. 2022. "Comparative Analysis of Intestinal Characteristics of Largemouth Bass (Micropterus salmoides) and Intestinal Flora with Different Growth Rates" Fishes 7, no. 2: 65. https://doi.org/10.3390/fishes7020065
APA StyleSu, X., Ji, D., Yao, J., Zou, Y., & Yan, M. (2022). Comparative Analysis of Intestinal Characteristics of Largemouth Bass (Micropterus salmoides) and Intestinal Flora with Different Growth Rates. Fishes, 7(2), 65. https://doi.org/10.3390/fishes7020065