
Early Immune Modulation in European Seabass (Dicentrarchus labrax) Juveniles in Response to Betanodavirus Infection

 ,
,  , ,
, ,  and
and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Time-Course Infection with RGNNV
2.2. Hematological Parameters
2.3. Innate Immune Humoral and Antioxidant Parameters
2.4. Viral Quantification
2.5. Immune Gene Expression
2.6. Statistical Analysis
3. Results
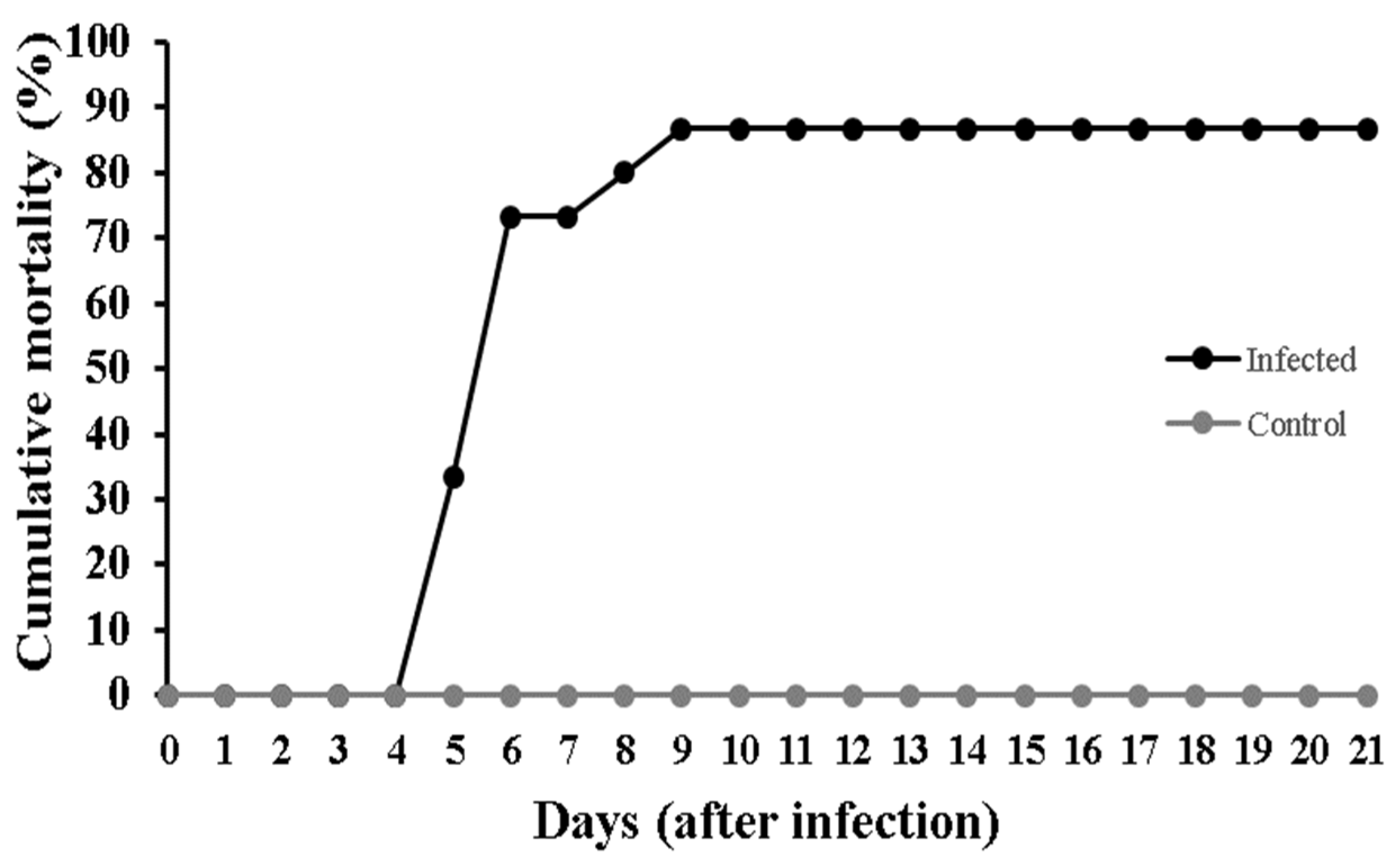
3.1. Mortality
3.2. Hematological Parameters
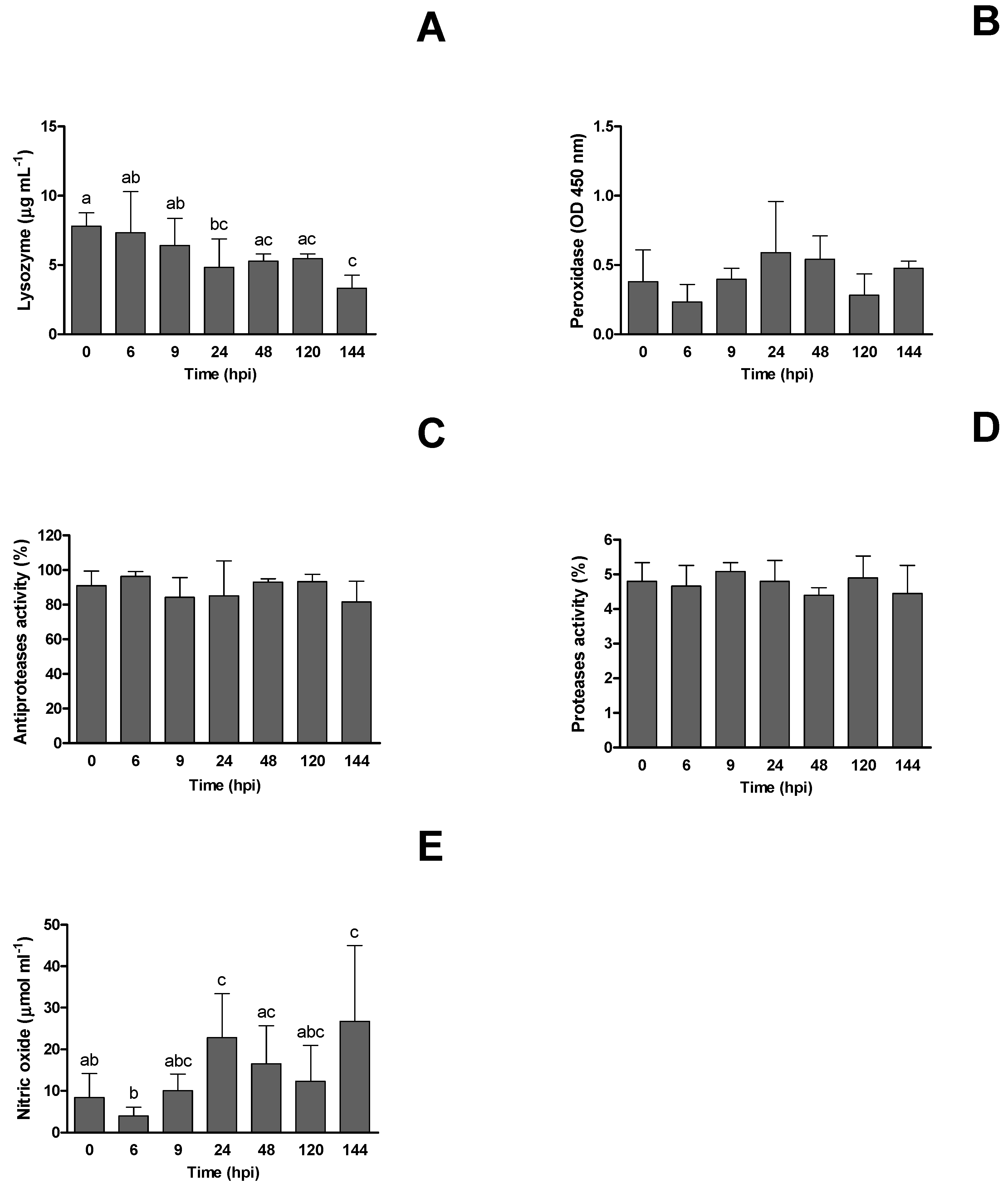
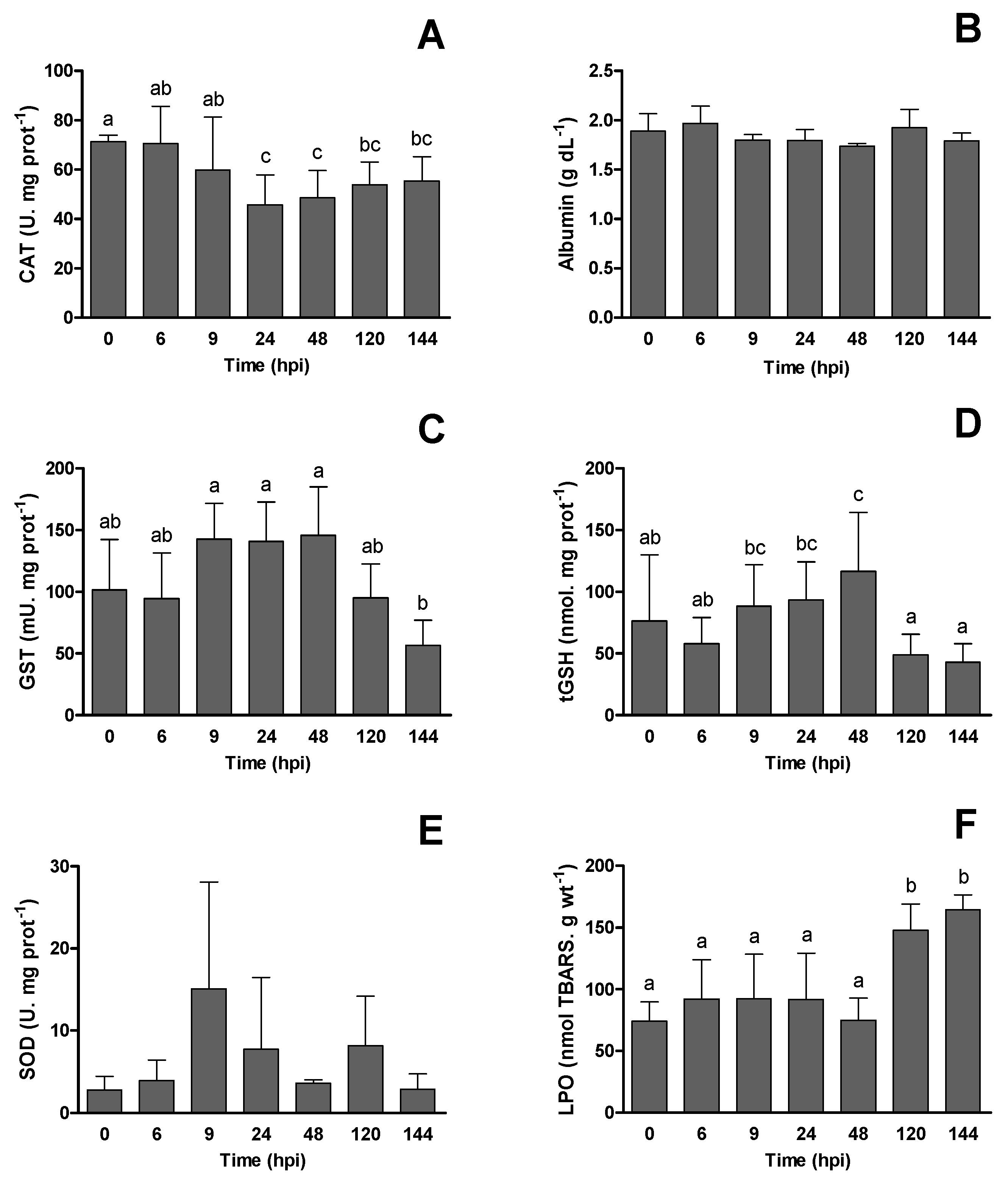
3.3. Innate Humoral Parameters and Biomarkers
3.4. Viral Quantification
3.5. Immune Genes Expression
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yang, Z.; Yue, G.H.; Wong, S.-M. VNN disease and status of breeding for resistance to NNV in aquaculture. Aquac. Fish. 2022, 7, 147–157. [Google Scholar] [CrossRef]
- Bandín, I.; Souto, S. Betanodavirus and VER Disease: A 30-year Research Review. Pathogens 2020, 9, 106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fauquet, C.M.; Mayo, M.A. The 7th ICTV Report. Arch. Virol. 2001, 146, 190–191. [Google Scholar] [CrossRef] [PubMed]
- Mori, K.-I.; Nakai, T.; Muroga, K.; Arimoto, M.; Mushiake, K.; Furusawas, I. Properties of a New Virus Belonging to Nodaviridae Found in Larval Striped Jack (Pseudocaranx dentex) with Nervous Necrosis. Virology 1992, 187, 368–371. [Google Scholar] [CrossRef]
- Delsert, C.; Morin, N.; Comps, M.A. Fish encephalitis virus that differs from other nodaviruses by its capsid protein processing. Arch. Virol. 1997, 142, 2359–2371. [Google Scholar] [CrossRef]
- Nishizawa, T.; Furuhashi, M.; Nagai, T.; Nakai, T.; Muroga, K. Genomic classification of fish nodaviruses by molecular phylogenetic analysis of the coat protein gene. Appl. Environ. Microbiol. 1997, 63, 1633–1636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zorriehzahra, M.J. Chapter 30—Viral Nervous Necrosis Disease. Emerg. Reemerging Viral Pathog. 2020, 1, 673–703. [Google Scholar] [CrossRef]
- Munday, B.L.; Kwang, J.; Moody, N. Betanodavirus infections of teleost fish: A review. J. Fish Dis. 2002, 25, 127–142. [Google Scholar] [CrossRef]
- Johansen, R.; Amundsen, M.; Dannevig, B.H.; Sommer, A.I. Acute and persistent experimental nodaviruses infection in spotted wolfish Anarhichas minor. Dis. Aquat. Org. 2003, 57, 35–41. [Google Scholar] [CrossRef]
- Panzarin, V.; Patarnello, P.; Mori, A.; Rampazzo, E.; Cappellozza, E.; Bovo, G.; Cattoli, G. Development and validation of a real-time TaqMan PCR assay for the detection of betanodavirus in clinical specimens. Arch. Virol. 2010, 155, 1193–1203. [Google Scholar] [CrossRef]
- Bitchava, K.; Chassalevris, T.; Lampou, E.; Athanassopoulou, F.; Economou, V.; Dovas, C.I. Occurrence and molecular characterization of betanodaviruses in fish and invertebrates of the Greek territorial waters. J. Fish Dis. 2019, 42, 1773–1783. [Google Scholar] [CrossRef] [PubMed]
- Ødegård, J.; Sommer, A.-I.; Præbel, A.K. Heritability of resistance to viral nervous necrosis in Atlantic cod (Gadus morhua L.). Aquaculture 2010, 300, 59–64. [Google Scholar] [CrossRef]
- Moreno, P.; Souto, S.; Leiva-Rebollo, R.; Borrego, J.J.; Bandín, I.; Alonso, M.C. Capsid amino acids at positions 247 and 270 are involved in the virulence of betanodaviruses to European sea bass. Sci. Rep. 2019, 9, 14068. [Google Scholar] [CrossRef] [PubMed]
- Poisa-Beiro, L.; Dios, S.; Montes, A.; Aranguren, R.; Figueras, A.; Novoa, B. Nodavirus increases the expression of Mx and inflammatory cytokines in fish brain. Mol. Immunol. 2008, 45, 218–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scapigliati, G.; Bounocore, F.; Randelli, E.; Casani, D.; Meloni, S.; Zarletti, G.; Tiberi, M.; Pietretti, D.; Boschi, I.; Manchado, M.; et al. Cellular and molecular immune responses of the sea bass (Dicentrarchus labrax) experimentally infected with betanodavirus. Fish Shellfish Immunol. 2010, 28, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Whyte, S.K. The innate immune response of finfish—A review of current knowledge. Fish Shellfish Immunol. 2007, 23, 1127–1151. [Google Scholar] [CrossRef] [PubMed]
- Bunnoy, A.; Na-Nakorn, U.; Srisapoome, P. Development of a Monoclonal Antibody Specific to the IgM Heavy Chain of Bighead Catfish (Clarias macrocephalus): A Biomolecular Tool for the Detection and Quantification of IgM Molecules and IgM+ Cells in Clarias Catfish. Biomolecules 2020, 10, 567. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, I.A.; Costa, J.Z.; Macchia, V.; Thompson, K.D. Detection of Betanodavirus in experimentally infected European seabass (Dicentrarchus labrax, Linnaeus 1758) using non-lethal sampling methods. J. Fish Dis. 2019, 42, 1097–1105. [Google Scholar] [CrossRef]
- Kaplow, L.S.; Ladd, C. Brief report: Simplified myeloperoxidase stain using benzidine dihychloride. Blood 1965, 26, 215–219. [Google Scholar] [CrossRef] [Green Version]
- Afonso, A.; Lousada, S.; Silva, J.; Ellis, A.E.; Silva, M.T. Neutrophil and macrophage responses to inflammation in the peritoneal cavity of rainbow trout Oncorhynchus mykiss. A light and electron microscopic cytochemical study. Dis. Aquat. Org. 1998, 34, 27–37. [Google Scholar] [CrossRef] [Green Version]
- Machado, M.; Azeredo, R.; Díaz-Rosales, P.; Afonso, A.; Peres, H.; Oliva-Teles, A.; Costas, B. Dietary tryptophan and methionine as modulators of European seabass (Dicentrarchus labrax) immune status and inflammatory response. Fish Shellfish Immunol. 2015, 42, 353–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costas, B.; Conceição, L.E.C.; Dias, J.; Novoa, B.; Figueras, A.; Afonso, A. Dietary arginine and repeated handling increase disease resistance and modulate innate immune mechanisms of Senegalese sole (Solea senegalensis Kaup, 1858). Fish Shellfish Immunol. 2011, 31, 838–847. [Google Scholar] [CrossRef]
- Quade, M.J.; Roth, J.A. A rapid, direct assay to measure degranulation of bovine neutrophil primary granules. Vet. Immunol. Immunopathol. 1997, 58, 239–248. [Google Scholar] [CrossRef]
- Taffala, C.; Gómez-León, J.; Novoa, B.; Figueras, A. Nitric oxide production by carpet shell clam (Ruditapes decussatus) hemocytes. Dev. Comp. Immunol. 2003, 27, 197–205. [Google Scholar] [CrossRef]
- Clairborne, A. Catalase activity. In Handbook of Methods of Oxygen Radical Research; Grenwald, R.A., Ed.; CRC Press: Boca Raton, FL, USA, 1985; pp. 283–284. [Google Scholar]
- Rodrigues, A.C.M.; Gravato, C.; Quintaneiro, C.; Bordalo, M.D.; Barata, C.; Soares, A.M.V.M.; Pestana, J.L.T. Energetic costs and biochemical biomarkers associated with esfenvalerate exposure in Sericostoma vittatum. Chemosphere 2017, 189, 445–453. [Google Scholar] [CrossRef] [PubMed]
- Bird, R.P.; Draper, A.H. Comparative studies on different methods of malondihaldehyde determination. Methods Enzymol. 1984, 90, 105–110. [Google Scholar]
- Habig, W.H.; Pabst, M.J.; Jakoby, W.B. Glutathione-S-transferases: The first enzymatic step in mercapturic acid formation. J. Biol. Chem. 1974, 249, 7130–7139. [Google Scholar] [CrossRef]
- Frasco, M.F.; Guilhermino, L. Effects of dimethoate and betaphthoflavone on selected biomarkers of Poecilia reticulate. Fish Physiol. Biochem. 2002, 26, 149–156. [Google Scholar] [CrossRef]
- Almeida, J.R.; Oliveira, C.; Gravato, C.; Guilhermino, L. Linking behavioural alterations with biomarkers responses in the European seabass Dicentrarchus labrax L. exposed to the organophosphate pesticide fenitrothion. Ecotoxicology 2010, 19, 1369–1381. [Google Scholar] [CrossRef]
- Baker, M.A.; Ceniglia, G.J.; Zaman, A. Microtiter plate assay for the measurement of glutathione and glutathione disulfide in large numbers of biological samples. Anal. Biochem. 1990, 190, 360–365. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT–PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Skliris, G.P.; Richards, R.H. Induction of nodavirus disease in seabass, Dicentrarchus labrax, using different infection models. Virus Res. 1999, 63, 85–93. [Google Scholar] [CrossRef]
- Lopez-Jimena, B.; Alonso, M.C.; Thompson, K.D.; Adams, A.; Infante, C.; Castro, D.; Borrego, J.J.; Garcia-Rosado, E. Tissue distribution of Red Spotted Grouper Nervous Necrosis Virus (RGNNV) genome in experimentally infected juvenile European seabass (Dicentrarchus labrax). Vet. Microbiol. 2011, 154, 86–95. [Google Scholar] [CrossRef] [PubMed]
- Guardiola, F.A.; Mabrok, M.; Machado, M.; Azeredo, R.; Afonso, A.; Esteban, M.A.; Costas, B. Mucosal and systemic immune responses in Senegalese sole (Solea senegalensis Kaup) bath challenged with Tenacibaculum maritimum: A time-course study. Fish Shellfish Immunol. 2019, 87, 744–754. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-O.; Kim, S.-J.; Kim, J.-O.; Kim, W.-S.; Oh, M.-J. Distribution of nervous necrosis virus (NNV) in infected sevenband grouper, Hyporthodus septemfasciatus by intramuscular injection or immersion challenge. Aquaculture 2018, 489, 1–8. [Google Scholar] [CrossRef]
- Stosik, M.; Tokarz-Deptula, B.; Deptula, W. Characterisation of thrombocytes in Osteichthyes. J. Vet. Res. 2019, 63, 123–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campos-Sánchez, J.C.; Esteban, M.Á. Review of inflammation in fish and value of the zebrafish model. J. Fish Dis. 2021, 44, 123–139. [Google Scholar] [CrossRef]
- Montoya, L.N.F.; Favero, G.C.; Zanuzzo, F.S.; Urbinati, E.C. Distinct β-glucan molecules modulates differently the circulating cortisol levels and innate immune responses in matrinxã (Brycon amazonicus). Fish Shellfish Immunol. 2018, 83, 314–320. [Google Scholar] [CrossRef] [PubMed]
- Guardiola, F.A.; Cuesta, A.; Abellán, E.; Mesenguer, J.; Esteban, M.A. Comparative analysis of the humoral immunity of skin mucus from several marine teleost fish. Fish Shellfish Immunol. 2014, 40, 24–31. [Google Scholar] [CrossRef] [PubMed]
- Passos, R.; Correia, A.P.; Pires, D.; Pires, P.; Ferreira, I.; Simões, M.; do Carmo, B.; Santos, P.; Pombo, A.; Afonso, C.; et al. Potential use of macroalgae Gracilaria gracilis in diets for European seabass (Dicentrarchus labrax): Health benefits from a sustainable source. Fish Shellfish Immunol. 2021, 119, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Reiss, C.S.; Komatsu, T. Does nitric oxide play a critical role in viral infections? J. Virol. 1998, 72, 4547–4551. Available online: https://journals.asm.org/doi/epdf/10.1128/JVI.72.6.4547-4551.1998 (accessed on 4 January 2022). [CrossRef] [PubMed] [Green Version]
- Eddy, F.B. Role of nitric oxide in larval and juvenile fish. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2005, 142, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Costas, B.; Rêgo, P.C.N.P.; Conceição, L.E.C.; Dias, J.; Afonso, A. Dietary arginine supplementation decreases plasma cortisol levels and modulates immune mechanisms in chronically stressed turbot (Scophthalmus maximus). Aquac. Nutr. 2013, 19, 25–38. [Google Scholar] [CrossRef]
- Gong, Q.; Yang, D.; Jiang, M.; Zheng, J.; Peng, B. l-aspartic acid promotes fish survival against Vibrio alginolyticus infection through nitric oxide-induced phagocytosis. Fish Shellfish Immunol. 2020, 97, 359–366. [Google Scholar] [CrossRef] [PubMed]
- Taffala, C.; Figueras, A.; Novoa, B. Role of nitric oxide on the replication of viral haemorrhagic septicemia virus (VHSV), a fish rhabdovirus. Vet. Immunol. Immunopathol. 1999, 72, 249–256. [Google Scholar] [CrossRef]
- López-Guerrero, J.A.; Carrasco, L. Effect of nitric oxide on poliovirus infection of two human cell lines. J. Virol. 1998, 72, 2538–2540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lisi, F.; Zelikin, A.N.; Chandrawati, R. Nitric oxide to fight viral infections. Adv. Sci. 2021, 8, 2003895. [Google Scholar] [CrossRef]
- Devi, G.; Harikrishnan, R.; Paray, B.A.; Al-Sadoon, M.K.; Hoseinifar, S.H.; Balasundaram, C. Effect of symbiotic supplemented diet on innate-adaptive immune response, cytokine gene regulation and antioxidant property in Labeo rohita against Aeromonas hydrophila. Fish Shellfish Immunol. 2019, 89, 687–700. [Google Scholar] [CrossRef]
- Hoseinifar, S.H.; Yousefi, S.; Doan, H.V.; Ashouri, G.; Gioacchini, G.; Maradonna, F.; Carnevali, O. Oxidative Stress and Antioxidant Defense in Fish: The Implications of Probiotic, Prebiotic, and Synbiotics. Rev. Fish. Sci. Aquac. 2020, 29, 198–217. [Google Scholar] [CrossRef]
- Wu, Y.-C.; Lu, Y.-F.; Chi, S.-C. Anti-viral mechanism of barramundi Mx against betanodavirus involves the inhibition of viral RNA synthesis through the interference of RdRp. Fish Shellfish Immunol. 2010, 28, 467–475. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.-C.; Tsai, P.-Y.; Chan, J.-C.; Chi, S.-C. Endogenous grouper and barramundi Mx proteins facilitated the clearance of betanodavirus RNA-dependent RNA polymerase. Dev. Comp. Immunol. 2016, 59, 110–120. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-M.; Su, Y.-L.; Shie, P.-S.; Huang, S.-L.; Yang, H.-L.; Chen, T.-Y. Grouper Mx confers resistance to nodavirus and interacts with coat protein. Dev. Comp. Immunol. 2008, 32, 825–836. [Google Scholar] [CrossRef] [PubMed]
- Chaves-Pozo, E.; Guardiola, F.A.; Meseguer, J.; Esteban, M.A.; Cuesta, A. Nodavirus infection induces a great innate cell-mediated cytotoxic activity in resistant, gilthead seabream, and susceptible, European sea bass, teleost fish. Fish Shellfish Immunol. 2012, 33, 1159–1166. [Google Scholar] [CrossRef]
- Carballo, C.; Garcia-Rosado, E.; Borrego, J.J.; Alonso, M.C. SJNNV down-regulates RGNNV replication in European sea bass by the induction of the type I interferon system. Vet. Res. 2016, 47, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreno, P.; Lopez-Jimena, B.; Randelli, E.; Scapigliati, G.; Buonocore, F.; Garcia-Rosado, E.; Borrego, J.J.; Alonso, M.C. Immuno-related gene transcription and antibody response in nodavirus (RGNNV and SJNNV)-infected European sea bass (Dicentrarchus labrax L.). Fish Shellfish Immunol. 2018, 78, 270–278. [Google Scholar] [CrossRef] [PubMed]
- Costa, J.Z.; Thompson, K.D. Understanding the interaction between Betanodavirus and its host for the development of prophylactic measures for viral encephalopathy and retinopathy. Fish Shellfish Immunol. 2016, 53, 35–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Muñoz, A.; Sepulcre, M.P.; García-Moreno, D.; Fuentes, I.; Béjar, J.; Manchado, M.; Álvarez, M.C.; Mesenguer, J.; Mulero, V. Viral nervous necrosis virus persistently replicates in the central nervous system of asymptomatic gilthead seabream and promotes a transient inflammatory response followed by the infiltration of IgM+ B lymphocytes. Dev. Comp. Immunol. 2012, 37, 429–437. [Google Scholar] [CrossRef] [PubMed]
- Valero, Y.; García-Alcázar, A.; Esteban, M.Á.; Cuesta, A.; Chaves-Pozo, E. Antimicrobial response is increased in the testis of European sea bass, but not in gilthead seabream, upon nodavirus infection. Fish Shellfish Immunol. 2015, 44, 203–213. [Google Scholar] [CrossRef]
- Valero, Y.; Arizcun, M.; Cortés, J.; Ramírez-Cepeda, F.; Guzmán, F.; Mercado, L.; Esteban, M.Á.; Chaves-Pozo, E.; Cuesta, A. NK-lysin, dicentracin and hepcidin antimicrobial peptides in European sea bass. Ontogenetic development and modulation in juveniles by nodavirus. Dev. Comp. Immunol. 2020, 103, 103516. [Google Scholar] [CrossRef]
- Falco, A.; Ortega-Villaizan, M.; Chico, V.; Brocal, I.; Perez, L.; Coll, J.M.; Estepa, A. Antimicrobial Peptides as Model Molecules for the Development of Novel Antiviral Agents in Aquaculture. Mini Rev. Med. Chem. 2009, 9, 1159–1164. [Google Scholar] [CrossRef]
- Boshra, H.; Li, J.; Sunyer, J.O. Recent advances on the complement system of teleost fish. Fish Shellfish Immunol. 2006, 20, 239–262. [Google Scholar] [CrossRef] [PubMed]
- Dezfuli, B.S.; Lui, A.; Giari, L.; Castaldelli, G.; Mulero, V.; Noga, E.J. Infiltration and activation of acidophilic granulocytes in skin lesions of gilthead seabream, Sparus aurata, naturally infected with lymphocystis disease virus. Dev. Comp. Immunol. 2012, 36, 174–182. [Google Scholar] [CrossRef] [PubMed]
- González-Fernández, C.; Esteban, M.A.; Cuesta, A. Molecular characterization of the T cell costimulatory receptors CD28 and CTLA4 in the European sea bass. Fish Shellfish Immunol. 2021, 109, 106–115. [Google Scholar] [CrossRef] [PubMed]
- Jeswin, J.; Jeong, S.-M.; Jeong, J.-M.; Bae, J.-S.; Kim, M.-C.; Kim, D.-H.; Park, C.-I. Molecular characterization of a T cell co-stimulatory receptor, CD28 of rock bream (Oplegnathus fasciatus): Transcriptional expression during bacterial and viral stimulation. Fish Shellfish Immunol. 2017, 66, 354–359. [Google Scholar] [CrossRef] [PubMed]
- Carreno, B.M.; Collins, M. The B7 family of ligands and its receptors: New Pathways for Costimulation and Inhibition of Immune Responses. Annu. Rev. Immunol. 2002, 20, 29–53. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, T.; Takizawa, F.; Furihata, M.; Soto-Lampe, V.; Dijkstra, J.M.; Fischer, U. Teleost cytotoxic T cells. Fish Shellfish Immunol. 2019, 95, 422–439. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.-h.; Sun, B.-g.; Deng, T.; Sun, L. Molecular characterization of Cynoglossus semilaevis CD28. Fish Shellfish Immunol. 2012, 32, 934–938. [Google Scholar] [CrossRef]
- Chen, Y.-M.; Tan, C.S.; Wang, T.-Y.; Hwong, C.-L.; Chen, T.-Y. Characterization of betanodavirus quasispecies influences on the subcellular localization and expression of tumor necrosis factor (TNF). Fish Shellfish Immunol. 2020, 103, 332–341. [Google Scholar] [CrossRef]
- Zou, J.; Secombes, C.J. The Function of Fish Cytokines. Biology 2016, 5, 23. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Chen, B.; Mao, M.; Chen, M.; Liu, X.; Cui, Q.; Liu, Y.; Jiang, C. Molecular characterization and expression analysis of the interleukin 1b gene in Pacific cod (Gadus macrocephalus). Dev. Comp. Immunol. 2018, 88, 213–218. [Google Scholar] [CrossRef]
- Chiang, Y.-H.; Wu, Y.-C.; Chi, S.-C. Interleukin-1β secreted from betanodavirus-infected microglia caused the death of neurons in giant grouper brains. Dev. Comp. Immunol. 2017, 70, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Park, H.; Li, Z.; Yang, X.O.; Chang, S.H.; Nurieva, R.; Wang, Y.-H.; Wang, Y.; Hood, L.; Zhu, Z.; Tian, Q.; et al. A distinct lineage of CD4 T cells regulates tissue inflammation by producing interleukin 17. Nat. Immunol. 2015, 6, 1133–1141. [Google Scholar] [CrossRef] [PubMed]
- Byadgi, O.; Beraldo, P.; Volpatti, D.; Massimo, M.; Bulfon, C.; Galeotti, M. Expression of infection-related immune response in European sea bass (Dicentrarchus labrax) during a natural outbreak from a unique dinoflagellate Amyloodinium ocellatum. Fish Shellfish Immunol. 2019, 84, 62–72. [Google Scholar] [CrossRef] [PubMed]
- Moreno, P.; Gemez-Mata, J.; Garcia-Rosado, E.; Bejar, J.; Labella, A.M.; Souto, S.; Alonso, M.C. Differential immunogene expression profile of European sea bass (Dicentrarchus labrax, L.) in response to highly and low virulent NNV. Fish Shellfish Immunol. 2020, 106, 56–70. [Google Scholar] [CrossRef] [PubMed]
- Valero, Y.; Morcillo, P.; Meseguer, J.; Bounocore, F.; Esteban, M.Á.; Chaves-Pozo, E.; Cuesta, A. Characterization of the IFN pathway in the teleost fish gonad against vertically transmitted viral nervous necrosis virus. J. Med. Virol. 2015, 96, 2176–2187. [Google Scholar] [CrossRef] [PubMed]
- Machado, M.; Azeredo, R.; Fontinha, F.; Fernández-Boo, S.; Conceição, L.E.C.; Dias, J.; Costas, B. Dietary Methionine Improves the European Seabass (Dicentrarchus labrax) Immune Status, Inflammatory Response, and Disease Resistance. Front. Immunol. 2018, 9, 2–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time (hpi) | |||||||
|---|---|---|---|---|---|---|---|
| 0 | 6 | 9 | 24 | 48 | 120 | 144 | |
| RBC (106 μL−1) | 2.49 ± 0.53 | 2.70 ± 0.48 | 3.20 ± 0.12 | 2.53 ± 0.56 | 2.96 ± 1.02 | 2.63 ± 0.49 | 2.27 ± 0.31 |
| WBC (104 μL−1) | 6.17 ± 1.21 a | 5.02 ± 1.19 ab | 3.92 ± 1.99 b | 4.98 ± 0.35 ab | 4.57 ± 1.40 ab | 5.37 ± 0.94 ab | 6.93 ± 2.00 a |
| Neutrophils (104 µL−1) | 0.09 ± 0.08 a | 1.04 ± 0.43 bc | 0.35 ± 0.19 b | 0.35 ± 0.05 b | 0.18 ± 0.12 a | 0.58 ± 0.22 bc | 1.40 ± 0.44 c |
| Monocytes (104 µL−1) | 0.07 ± 0.06 a | 0.28 ± 0.13 b | 0.09 ± 0.09 ac | 0.19 ± 0.07 bc | 0.07 ± 0.05 ac | 0.21 ± 0.11 bc | 0.94 ± 0.57 b |
| Lymphocytes (104 µL−1) | 2.29 ± 0.47 a | 1.11 ± 0.26 b | 1.22 ± 0.58 b | 1.34 ± 0.37 b | 1.76 ± 0.36 ab c | 2.25 ± 0.24 ac | 1.48 ± 0.83 bc |
| Thrombocytes (104 µL−1) | 3.81 ± 0.85 a | 2.60 ± 0.79 ab | 2.05 ± 1.01 b | 3.29 ± 0.58 ab | 2.55 ± 1.06 ab | 2.56 ± 0.42 ab | 3.12 ± 0.99 ab |
| Hb (g dL−1) | 1.85 ± 0.70 a | 2.57 ± 0.66 ab | 2.37 ± 0.50 ab | 3.36 ± 1.21 b | 2.79 ± 0.93 ab | 2.66 ± 0.34 ab | 2.92 ± 0.50 ab |
| Ht (%) | 25.45 ± 3.59 | 25.80 ± 1.48 | 22.80 ± 2.39 | 25.20 ± 3.70 | 25.50 ± 3.11 | 26.33 ± 3.39 | 22.50 ± 3.67 |
| MCV (μm3) | 89.81 ± 9.60 | 92.52 ± 2.93 | 75.19 ± 1.60 | 103.68 ± 21.3 | 114.10 ± 83.5 | 95.06 ± 10.56 | 91.97 ± 17.79 |
| MCH (pg cell−1) | 7.27 ± 2.31 a | 9.60 ± 2.25 ab c | 7.33 ± 1.70 ac | 13.70 ± 4.88 b | 11.55 ± 9.3 ab c | 10.30 ± 1.58 b | 12.01 ± 2.72 b |
| MCHC (g 100 mL−1) | 8.04 ± 0.53 a | 10.22 ± 1.67 ab | 10.25 ± 1.13 ab | 13.52 ± 4.47 b | 12.57 ± 0.74 b | 10.14 ± 1.16 ab | 13.04 ± 1.61 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vaz, M.; Pires, D.; Pires, P.; Simões, M.; Pombo, A.; Santos, P.; do Carmo, B.; Passos, R.; Costa, J.Z.; Thompson, K.D.; et al. Early Immune Modulation in European Seabass (Dicentrarchus labrax) Juveniles in Response to Betanodavirus Infection. Fishes 2022, 7, 63. https://doi.org/10.3390/fishes7020063
Vaz M, Pires D, Pires P, Simões M, Pombo A, Santos P, do Carmo B, Passos R, Costa JZ, Thompson KD, et al. Early Immune Modulation in European Seabass (Dicentrarchus labrax) Juveniles in Response to Betanodavirus Infection. Fishes. 2022; 7(2):63. https://doi.org/10.3390/fishes7020063
Chicago/Turabian StyleVaz, Mariana, Damiana Pires, Pedro Pires, Marco Simões, Ana Pombo, Paulo Santos, Beatriz do Carmo, Ricardo Passos, Janina Z. Costa, Kim D. Thompson, and et al. 2022. "Early Immune Modulation in European Seabass (Dicentrarchus labrax) Juveniles in Response to Betanodavirus Infection" Fishes 7, no. 2: 63. https://doi.org/10.3390/fishes7020063
APA StyleVaz, M., Pires, D., Pires, P., Simões, M., Pombo, A., Santos, P., do Carmo, B., Passos, R., Costa, J. Z., Thompson, K. D., & Baptista, T. (2022). Early Immune Modulation in European Seabass (Dicentrarchus labrax) Juveniles in Response to Betanodavirus Infection. Fishes, 7(2), 63. https://doi.org/10.3390/fishes7020063