
Antimicrobial and Toxic Activity of Citronella Essential Oil (Cymbopogon nardus), and Its Effect on the Growth and Metabolism of Gilthead Seabream (Sparus aurata L.)

 , ,
, ,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. In Vitro Antimicrobial Activity
2.2. Assessment of CEO Acute Toxicity
2.3. Experimental Diets and Feeding Trial
2.4. Injection of Poly I:C for Immune-Stimulation after Feeding Trial
2.5. Sampling Procedures
2.6. Growth Performance
2.7. Plasma and Hepatic Parameters
2.8. Haematological Analyses
2.9. Statistical Analyses
3. Results
3.1. Antimicrobial Activity of CEO
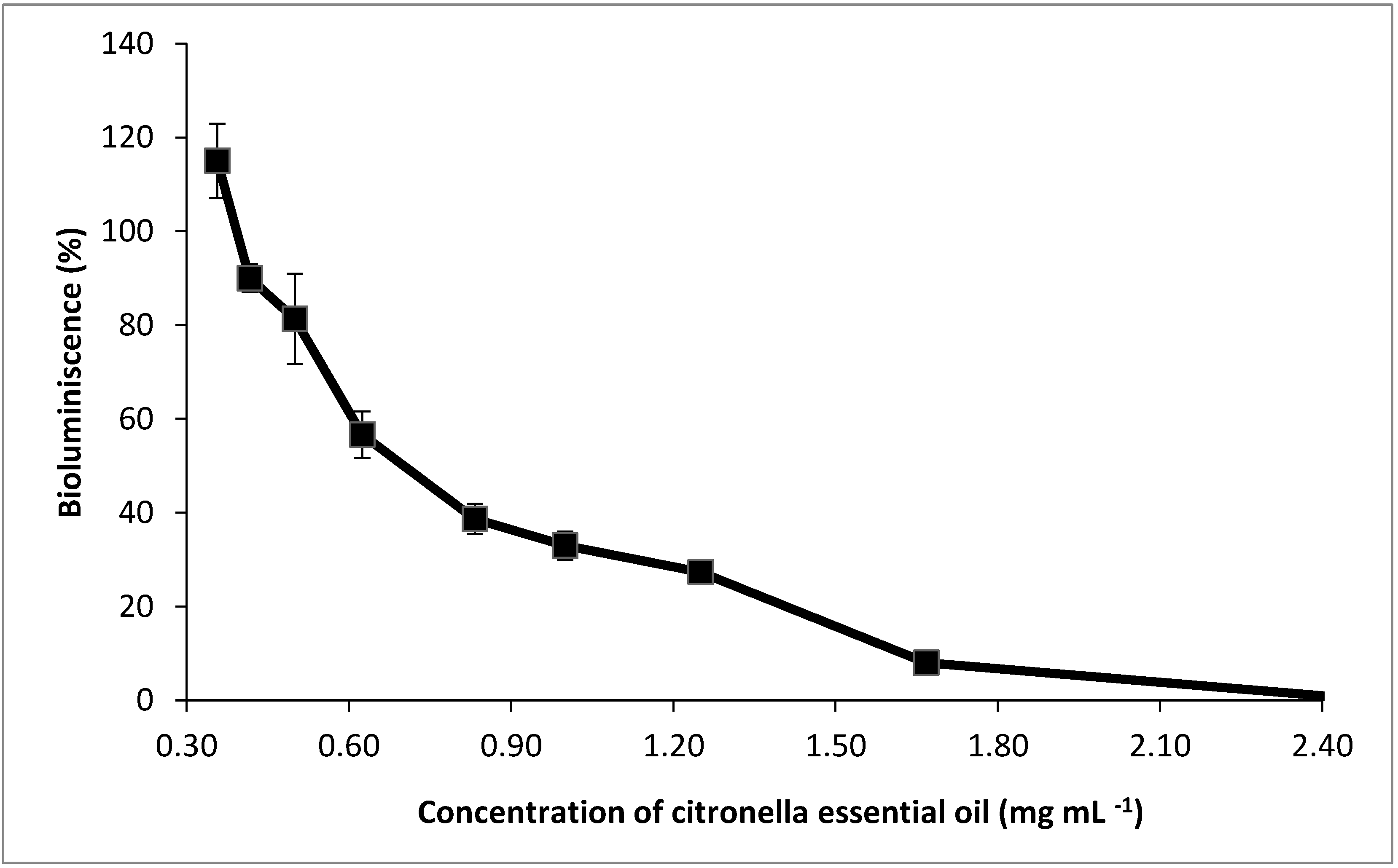
3.2. Acute Toxicity of CEO
3.3. Growth Performance
3.4. Plasma and Liver Metabolites
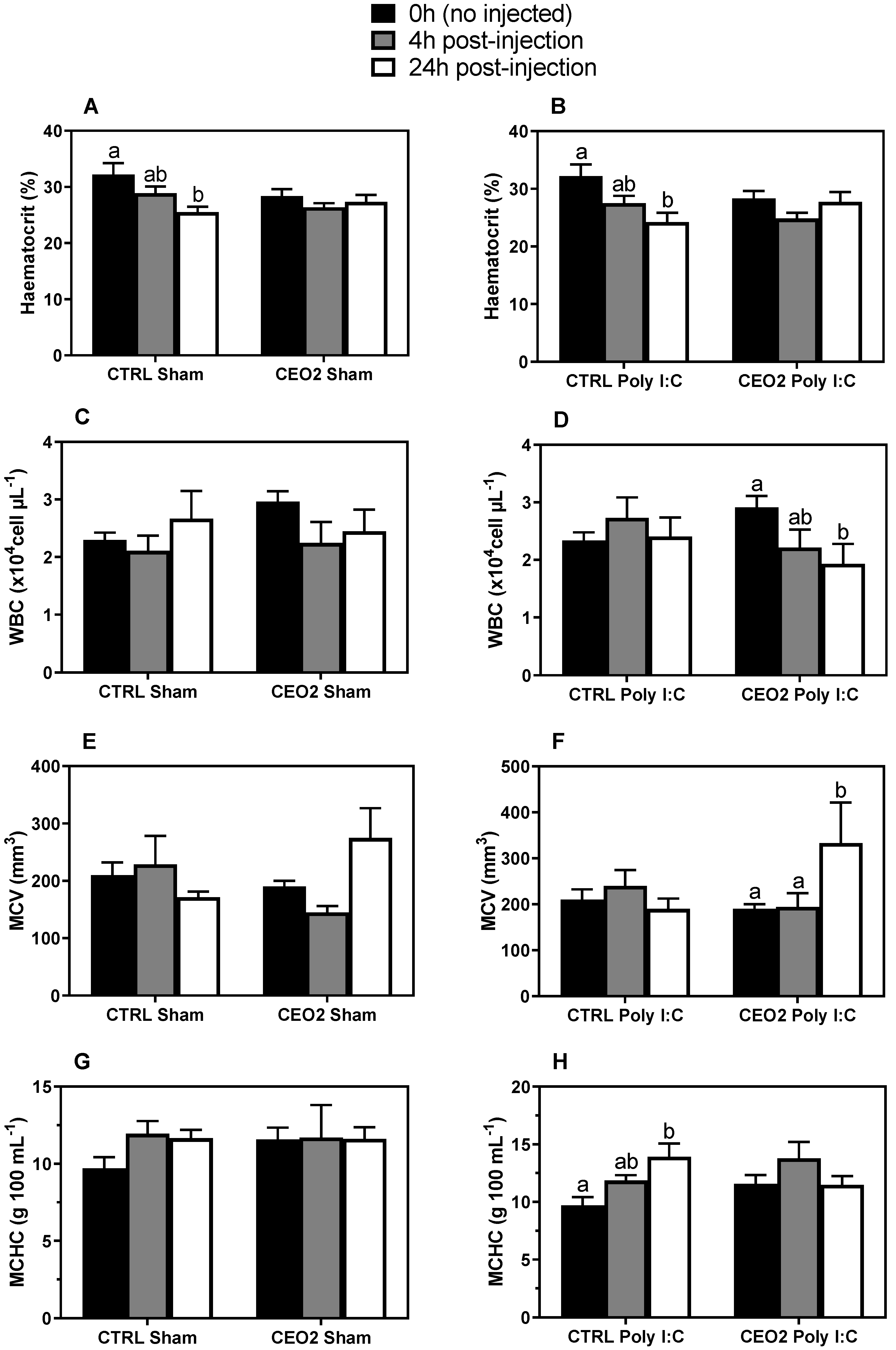
3.5. Hematological Parameters
3.6. Inflammatory Challenge after Feeding Trial
4. Discussion
4.1. Antimicrobial Activity of CEO
4.2. Acute Toxicity of CEO
4.3. In Vivo Effects of CEO
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Buján, N.; Toranzo, A.E.; Magariños, B. Edwardsiella piscicida: A significant bacterial pathogen of cultured fish. Dis. Aquat. Org. 2018, 131, 59–71. [Google Scholar] [CrossRef]
- Abdel-Latif, H.M.R.; Khafaga, A.F. Natural co-infection of cultured Nile tilapia Oreochromis niloticus with Aeromonas hydrophila and Gyrodactylus cichlidarum experiencing high mortality during summer. Aquac. Res. 2020, 51, 1880–1892. [Google Scholar] [CrossRef]
- Schreck, C.B.; Tort, L. The Concept of Stress in Fish. In Biology of Stress in Fish: Fish Physiology; Schreck, C.B., Tort, L., Farrell, A.P., Brauner, C.J., Eds.; Elsevier Inc.: London, UK, 2016; Volume 35, pp. 1–34. [Google Scholar] [CrossRef]
- Bonga, S.E.W. The stress response in fish. Physiol. Rev. 1997, 77, 591–625. [Google Scholar] [CrossRef] [PubMed]
- Andreoni, F.; Magnani, M. Photobacteriosis: Prevention and Diagnosis. J. Immunol. Res. 2014, 2014, 793817. [Google Scholar] [CrossRef]
- Bulfon, C.; Volpatti, D.; Galeotti, M. Current research on the use of plant-derived products in farmed fish. Aquac. Res. 2015, 46, 513–551. [Google Scholar] [CrossRef]
- Rahman, A.N.A.; ElHady, M.; Shalaby, S.I. Efficacy of the dehydrated lemon peels on the immunity, enzymatic antioxidant capacity and growth of Nile tilapia (Oreochromis niloticus) and African catfish (Clarias gariepinus). Aquaculture 2019, 505, 92–97. [Google Scholar] [CrossRef]
- Immanuel, G.; Uma, R.P.; Iyapparaj, P.; Citarasu, T.; Peter, S.M.P.; Babu, M.M.; Palavesam, A. Dietary medicinal plant extracts improve growth, immune activity and survival of tilapia Oreochromis mossambicus. J. Fish Biol. 2009, 74, 1462–1475. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Z.; Zhang, S.; Wang, H.; Piao, X. Essential oil and aromatic plants as feed additives in non-ruminant nutrition: A review. J. Anim. Sci. Biotechnol. 2015, 6, 7. [Google Scholar] [CrossRef]
- Citarasu, T. Herbal biomedicines: A new opportunity for aquaculture industry. Aquac. Int. 2009, 18, 403–414. [Google Scholar] [CrossRef]
- Van Hai, N. The use of medicinal plants as immunostimulants in aquaculture: A review. Aquaculture 2015, 446, 88–96. [Google Scholar] [CrossRef]
- Bandeira, G.; Pês, T.S.; Saccol, E.M.; Sutili, F.J.; Rossi, W.; Murari, A.L.; Heinzmann, B.M.; Pavanato, M.A.; de Vargas, A.C.; Silva, L.D.L.; et al. Potential uses of Ocimum gratissimum and Hesperozygis ringens essential oils in aquaculture. Ind. Crop. Prod. 2017, 97, 484–491. [Google Scholar] [CrossRef]
- Abarike, E.D.; Jian, J.; Tang, J.; Cai, J.; Yu, H.; Lihua, C.; Jun, L. Influence of traditional Chinese medicine and Bacillus species (TCMBS) on growth, immune response and disease resistance in Nile tilapia, Oreochromis niloticus. Aquac. Res. 2018, 49, 2366–2375. [Google Scholar] [CrossRef]
- Timung, R.; Barik, C.R.; Purohit, S.; Goud, V.V. Composition and anti-bacterial activity analysis of citronella oil obtained by hydrodistillation: Process optimization study. Ind. Crop. Prod. 2016, 94, 178–188. [Google Scholar] [CrossRef]
- Nakahara, K.; Alzoreky, N.S.; Yoshihashi, T.; Nguyen, H.T.T.; Trakoontivakorn, G. Chemical composition and antifungal activity of essential oil from Cymbopogon nardus (Citronella grass). Jpn. Agric. Res. Q. 2013, 37, 249–252. [Google Scholar] [CrossRef]
- Wei, L.S.; Wee, W. Chemical composition and antimicrobial activity of Cymbopogon nardus citronella essential oil against systemic bacteria of aquatic animals. Iran. J. Microbiol. 2013, 5, 147–152. [Google Scholar]
- Pereira, F.D.O.; Mendes, J.M.; Lima, I.O.; Mota, K.S.D.L.; De Oliveira, W.A.; Lima, E.D.O. Antifungal activity of geraniol and citronellol, two monoterpenes alcohols, against Trichophyton rubrum involves inhibition of ergosterol biosynthesis. Pharm. Biol. 2014, 53, 228–234. [Google Scholar] [CrossRef]
- Kaiser, K.L.E.; Ribo, J.M. Photobacterium phosphoreum toxicity bioassay. II. Toxicity data compilation. Environ. Toxicol. Water Qual. 1988, 3, 195–237. [Google Scholar] [CrossRef]
- Backhaus, T.; Scholze, M.; Grimme, L. The single substance and mixture toxicity of quinolones to the bioluminescent bacterium Vibrio fischeri. Aquat. Toxicol. 2000, 49, 49–61. [Google Scholar] [CrossRef]
- Parvez, S.; Venkataraman, C.; Mukherji, S. A review on advantages of implementing luminescence inhibition test (Vibrio fischeri) for acute toxicity prediction of chemicals. Environ. Int. 2006, 32, 265–268. [Google Scholar] [CrossRef]
- Barbas, L.A.L.; Hamoy, M.; de Mello, V.J.; Barbosa, R.P.M.; de Lima, H.S.T.; Torres, M.F.; Nascimento, L.A.S.D.; da Silva, J.K.D.R.; Andrade, E.H.D.A.; Gomes, M.R.F. Essential oil of citronella modulates electrophysiological responses in tambaqui Colossoma macropomum: A new anaesthetic for use in fish. Aquaculture 2017, 479, 60–68. [Google Scholar] [CrossRef]
- Aydın, B.; Barbas, L.A. Sedative and anesthetic properties of essential oils and their active compounds in fish: A review. Aquaculture 2020, 520, 734999. [Google Scholar] [CrossRef]
- Al-Sagheer, A.A.; Mahmoud, H.K.; Reda, F.M.; Mahgoub, S.A.; Ayyat, M.S. Supplementation of diets for Oreochromis niloticus with essential oil extracts from lemongrass (Cymbopogon citratus) and geranium (Pelargonium graveolens) and effects on growth, intestinal microbiota, antioxidant and immune activities. Aquac. Nutr. 2018, 24, 1006–1014. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations (FAO). Global Aquaculture Production 1950–2016; FAO: Rome, Italy, 2019. [Google Scholar]
- Zorrilla, I.; Chabrillón, M.; Arijo, S.; Díaz-Rosales, P.; Martínez-Manzanares, E.; Balebona, M.; Moriñigo, M. Ángel Bacteria recovered from diseased cultured gilthead sea bream (Sparus aurata L.) in southwestern Spain. Aquaculture 2003, 218, 11–20. [Google Scholar] [CrossRef]
- Arijo, S.; Chabrillón, M.; Díaz-Rosales, P.; Rico, R.; Martínez-Manzanares, E.; Balebona, M.C.; Toranzo, A.E.; Moriñigo, M.A. Bacteria isolated from outbreaks affecting cultured sole, Solea senegalensis (Kaup). Bull. Eur. Assoc. Fish Pathol. 2005, 25, 148–154. [Google Scholar]
- Arijo, S.; Rico, R.; Chabrillón, M.; Díaz-Rosales, P.; Martinez-Manzanares, E.; Balebona, M.C.; Magariños, B.; Toranzo, A.E.; Moriñigo, M.A. Effectiveness of a divalent vaccine for sole, Solea senegalensis (Kaup), against Vibrio harveyi and Photobacterium damselae subsp. piscicida. J. Fish Dis. 2005, 28, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Kantachote, D.; Rattanachuay, P.; Tantirungkij, M.; Nitoda, T.; Kanzaki, H. Inhibition of shrimp pathogenic vibrios by extracellular compounds from a proteolytic bacterium Pseudomonas sp. W3. Electron. J. Biotechnol. 2010, 13, 8–9. [Google Scholar] [CrossRef][Green Version]
- Johnson, B.T. Microtox® Acute Toxicity Test; Springer: Berlin/Heidelberg, Germany, 2005; pp. 69–105. [Google Scholar]
- Ajuzieogu, C.A.; Odokuma, L.O.; Chikere, C.B. Toxicity Assessment of Produced Water Using Microtox Rapid Bioassay. South Asian J. Res. Microbiol. 2018, 1, 1–9. [Google Scholar] [CrossRef]
- Morgado, I.; Campinho, M.A.; Costa, R.; Jacinto, R.; Power, D. Disruption of the thyroid system by diethylstilbestrol and ioxynil in the sea bream (Sparus aurata). Aquat. Toxicol. 2009, 92, 271–280. [Google Scholar] [CrossRef]
- Keppler, D.; Decker, K. Glycogen: Determination with amyloglucosidase. In Methods of Enzymatic Analysis; Bergmeyer, H.U., Ed.; Verlag-Chemie: Deerfield Beach, FL, USA; Academic Press: Cambridge, MA, USA, 1974; pp. 1127–1131. [Google Scholar]
- Stevens, M.L. Fundamentals of Clinical Hematology; W.B. Saunders: Philadelphia, PA, USA, 1997. [Google Scholar]
- Cunha, J.; Heinzmann, B.M.; Baldisserotto, B.; Da Cunha, J.A. The effects of essential oils and their major compounds on fish bacterial pathogens—A review. J. Appl. Microbiol. 2018, 125, 328–344. [Google Scholar] [CrossRef]
- Assane, I.M.; Valladão, G.M.R.; Pilarski, F. Chemical composition, cytotoxicity and antimicrobial activity of selected plant-derived essential oils against fish pathogens. Aquac. Res. 2021, 52, 793–809. [Google Scholar] [CrossRef]
- Hammer, K.A.; Carson, C.F.; Riley, T.V. Antimicrobial activity of essential oils and other plant extracts. J. Appl. Microbiol. 1999, 86, 985–990. [Google Scholar] [CrossRef] [PubMed]
- Abbas, M.; Adil, M.; Ehtisham-Ul-Haque, S.; Munir, B.; Yameen, M.; Ghaffar, A.; Shar, G.A.; Tahir, M.A.; Iqbal, M. Vibrio fischeri bioluminescence inhibition assay for ecotoxicity assessment: A review. Sci. Total Environ. 2018, 626, 1295–1309. [Google Scholar] [CrossRef] [PubMed]
- Conforti, F.; Ioele, G.; Statti, G.A.; Marrelli, M.; Ragno, G.; Menichini, F. Antiproliferative activity against human tumor cell lines and toxicity test on Mediterranean dietary plants. Food Chem. Toxicol. 2008, 46, 3325–3332. [Google Scholar] [CrossRef]
- Skotti, E.; Anastasaki, E.; Kanellou, G.; Polissiou, M.; Tarantilis, P.A. Total phenolic content, antioxidant activity and toxicity of aqueous extracts from selected Greek medicinal and aromatic plants. Ind. Crop. Prod. 2014, 53, 46–54. [Google Scholar] [CrossRef]
- Sotiropoulou, Ν.-S.D.; Flampouri, E.; Skotti, E.; Pappas, C.; Kintzios, S.; Tarantilis, P.A. Bioactivity and toxicity evaluation of infusions from selected Greek herbs. Food Biosci. 2020, 35, 100598. [Google Scholar] [CrossRef]
- Dinardo, F.R.; Deflorio, M.; Casalino, E.; Crescenzo, G.; Centoducati, G. Effect of feed supplementation with Origanum vulgare L. essential oil on sea bass (Dicentrarchus labrax): A preliminary framework on metabolic status and growth performances. Aquac. Rep. 2020, 18, 100511. [Google Scholar] [CrossRef]
- Ghafarifarsani, H.; Hoseinifar, S.H.; Adorian, T.J.; Ferrigolo, F.R.G.; Raissy, M.; Van Doan, H. The effects of combined inclusion of Malvae sylvestris, Origanum vulgare, and Allium hirtifolium boiss for common carp (Cyprinus carpio) diet: Growth performance, antioxidant defense, and immunological parameters. Fish Shellfish Immunol. 2021, 119, 670–677. [Google Scholar] [CrossRef]
- Tavares-Dias, M. Current knowledge on use of essential oils as alternative treatment against fish parasites. Aquat. Living Resour. 2018, 31, 13. [Google Scholar] [CrossRef]
- Talpur, A.D.; Ikhwanuddin, M. Azadirachta indica (neem) leaf dietary effects on the immunity response and disease resistance of Asian seabass, Lates calcarifer challenged with Vibrio harveyi. Fish Shellfish Immunol. 2013, 34, 254–264. [Google Scholar] [CrossRef]
- Awad, E.; Cerezuela, R.; Esteban, M. Ángeles Effects of fenugreek (Trigonella foenum graecum) on gilthead seabream (Sparus aurata L.) immune status and growth performance. Fish Shellfish Immunol. 2015, 45, 454–464. [Google Scholar] [CrossRef]
- Gabriel, N.N.; Qiang, J.; He, J.; Ma, X.Y.; Kpundeh, M.D.; Xu, P. Dietary Aloe vera supplementation on growth performance, some haemato-biochemical parameters and disease resistance against Streptococcus iniae in tilapia (GIFT). Fish Shellfish Immunol. 2015, 44, 504–514. [Google Scholar] [CrossRef] [PubMed]
- Saccol, E.; Parrado-Sanabria, Y.; Gagliardi, L.; Jerez-Cepa, I.; Mourão, R.; Heinzmann, B.; Baldisserotto, B.; Pavanato, M.; Mancera, J.; Martos-Sitcha, J. Myrcia sylvatica essential oil in the diet of gilthead sea bream (Sparus aurata L.) attenuates the stress response induced by high stocking density. Aquac. Nutr. 2018, 24, 1381–1392. [Google Scholar] [CrossRef]
- Beltrán, J.M.G.; Silvera, D.G.; Ruiz, C.E.; Campo, V.; Chupani, L.; Faggio, C.; Esteban, M. Ángeles Effects of dietary Origanum vulgare on gilthead seabream (Sparus aurata L.) immune and antioxidant status. Fish Shellfish Immunol. 2020, 99, 452–461. [Google Scholar] [CrossRef] [PubMed]
- Firmino, J.P.; Vallejos-Vidal, E.; Sarasquete, C.; Ortiz-Delgado, J.B.; Balasch, J.C.; Tort, L.; Estevez, A.; Reyes-López, F.E.; Gisbert, E. Unveiling the effect of dietary essential oils supplementation in Sparus aurata gills and its efficiency against the infestation by Sparicotyle chrysophrii. Sci. Rep. 2020, 10, 17764. [Google Scholar] [CrossRef]
- Barany, A.; Guilloto, M.; Cosano, J.; de Boevre, M.; Oliva, M.; de Saeger, S.; Fuentes, J.; Martínez-Rodriguez, G.; Mancera, J. Dietary aflatoxin B1 (AFB1) reduces growth performance, impacting growth axis, metabolism, and tissue integrity in juvenile gilthead sea bream (Sparus aurata). Aquaculture 2021, 533, 736189. [Google Scholar] [CrossRef]
- Glover, E.C.; Daland, G.A.; Schmitz, H.L. The metabolism of normal and leukemic leukocytes. Arch. Intern. Med. 1930, 46, 46–66. [Google Scholar] [CrossRef]
- Harikrishnan, R.; Kim, M.-C.; Kim, J.-S.; Balasundaram, C.; Heo, M.-S. Protective effect of herbal and probiotics enriched diet on haematological and immunity status of Oplegnathus fasciatus (Temminck & Schlegel) against Edwardsiella tarda. Fish Shellfish Immunol. 2011, 30, 886–893. [Google Scholar] [CrossRef]
- Kuang, S.-Y.; Xiao, W.-W.; Feng, L.; Liu, Y.; Jiang, J.; Jiang, W.-D.; Hu, K.; Li, S.-H.; Tang, L.; Zhou, X.-Q. Effects of graded levels of dietary methionine hydroxy analogue on immune response and antioxidant status of immune organs in juvenile Jian carp (Cyprinus carpio var. Jian). Fish Shellfish Immunol. 2012, 32, 629–636. [Google Scholar] [CrossRef]
- Machado, M.; Azeredo, R.; Diaz-Rosales, P.; Afonso, A.; Peres, H.; Oliva-Teles, A.; Costas, B. Dietary tryptophan and methionine as modulators of European seabass (Dicentrarchus labrax) immune status and inflammatory response. Fish Shellfish Immunol. 2015, 42, 353–362. [Google Scholar] [CrossRef]
- Machado, M.; Azeredo, R.; Fontinha, F.; Fernández-Boo, S.; Conceicao, L.; Dias, J.; Costas, B. Dietary Methionine Improves the European Seabass (Dicentrarchus labrax) Immune Status, Inflammatory Response, and Disease Resistance. Front. Immunol. 2018, 9, 2672. [Google Scholar] [CrossRef]
- Vallejos-Vidal, E.; Reyes-López, F.; Teles, M.; MacKenzie, S. The response of fish to immunostimulant diets. Fish Shellfish Immunol. 2016, 56, 34–69. [Google Scholar] [CrossRef] [PubMed]
- Malka, R.; Delgado, F.F.; Manalis, S.R.; Higgins, J.M. In Vivo Volume and Hemoglobin Dynamics of Human Red Blood Cells. PLoS Comput. Biol. 2014, 10, e1003839. [Google Scholar] [CrossRef] [PubMed]
- De Andrade, T.U.; Brasil, G.A.; Endringer, D.C.; Da Nóbrega, F.R.; de Sousa, D. Cardiovascular Activity of the Chemical Constituents of Essential Oils. Molecules 2017, 22, 1539. [Google Scholar] [CrossRef] [PubMed]
- Machado, M.; Azeredo, R.; Domingues, A.; Boo, S.F.; Dias, J.; Conceicao, L.; Costas, B. Dietary tryptophan deficiency and its supplementation compromises inflammatory mechanisms and disease resistance in a teleost fish. Sci. Rep. 2019, 9, 7689. [Google Scholar] [CrossRef]
- Engelsma, M. Multiple acute temperature stress affects leucocyte populations and antibody responses in common carp, Cyprinus carpio L. Fish Shellfish Immunol. 2003, 15, 397–410. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Extract | V. harveyi | A. hydrophila | P. damselae subsp. piscicida | V. fischeri |
|---|---|---|---|---|
| Saline solution | 27.8 | 55.6 | 27.8 | 27.8 |
| Ethanol | 111.3 | 27.8 | 27.8 | 27.8 |
| CTRL | CEO1 | CEO2 | |
|---|---|---|---|
| Initial Weight (g) | 17.9 ± 0.3 | 18.0 ± 0.3 | 17.9 ± 0.3 |
| Final Weight (g) | 44.8 ± 0.9 | 44.5 ± 1.0 | 44.6 ± 0.8 |
| Initial Length (cm) | 10.4 ± 0.1 | 10.4 ± 0.1 | 10.3 ± 0.1 |
| Final Length (cm) | 13.3 ± 0.1 | 13.2 ± 0.1 | 13.2 ± 0.1 |
| WG (g) | 27.6 ± 6.8 | 25.9 ± 7.4 | 27.3 ± 6.2 |
| WG (%) | 163.3 ± 6.7 | 142.7 ± 7.1 | 165.5 ± 6.5 |
| SGR (% day−1) | 1.6 ± 0.0 ab | 1.4 ± 0.0 a | 1.6 ± 0.0 b |
| K (g cm−3) | 19.1 ± 0.2 | 19.2 ± 0.1 | 19.4 ± 0.2 |
| HSI (%) | 1.3 ± 0.1 | 1.3 ± 0.1 | 1.2 ± 0.1 |
| CTRL | CEO1 | CEO2 | |
|---|---|---|---|
| Plasma | |||
| Glucose (mM) | 3.1 ± 0.2 | 3.6 ± 0.2 | 3.3 ± 0.2 |
| Triglycerides (mM) | 2.4 ± 0.1 | 2.1 ± 0.1 | 2.0 ± 0.2 |
| Lactate (mM) | 2.2 ± 0.3 | 1.4 ± 0.1 | 1.1 ± 0.1 |
| Liver | |||
| Glucose (mmol g−1 w.w.) | 36.3 ± 3.8 | 37.9 ± 4.5 | 41.2 ± 1.9 |
| Glycogen (mmol g−1 w.w.) | 285.4 ± 29.5 a | 267.6 ± 20.5 ab | 248.9 ± 20.6 b |
| Triglycerides (mmol g−1 w.w.) | 7.2 ± 0.5 | 4.9 ± 1.0 | 5.1 ± 0.6 |
| CTRL | CEO1 | CEO2 | |
|---|---|---|---|
| WBC (×104 cell µL−1) | 2.3 ± 0.1 a | 2.5 ± 0.1 ab | 3.0 ± 0.2 b |
| RBC (×106 cell µL−1) | 1.6 ± 0.2 a | 1.1 ± 0.1 b | 1.5 ± 0.1 ab |
| Hematocrit (%) | 32.2 ± 2.0 | 29.1 ± 1.3 | 28.3 ± 1.3 |
| Hemoglobin (g dL−1) | 3.1 ± 0.3 | 3.1 ± 0.2 | 3.2 ± 0.1 |
| MCV (mm3) | 209.9 ± 22.3 ab | 256.6 ± 15.2 a | 190.3 ± 10.0 b |
| MCH (pg cell−1) | 21.0 ± 2.8 | 27.2 ± 4.4 | 22.1 ± 1.8 |
| MCHC (g 100 mL−1) | 9.7 ± 0.7 | 9.8 ± 1.5 | 11.5 ± 0.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Márquez, J.; Barany, A.; Ruiz, Á.B.; Costas, B.; Arijo, S.; Mancera, J.M. Antimicrobial and Toxic Activity of Citronella Essential Oil (Cymbopogon nardus), and Its Effect on the Growth and Metabolism of Gilthead Seabream (Sparus aurata L.). Fishes 2021, 6, 61. https://doi.org/10.3390/fishes6040061
García-Márquez J, Barany A, Ruiz ÁB, Costas B, Arijo S, Mancera JM. Antimicrobial and Toxic Activity of Citronella Essential Oil (Cymbopogon nardus), and Its Effect on the Growth and Metabolism of Gilthead Seabream (Sparus aurata L.). Fishes. 2021; 6(4):61. https://doi.org/10.3390/fishes6040061
Chicago/Turabian StyleGarcía-Márquez, Jorge, Andre Barany, Álvaro Broz Ruiz, Benjamín Costas, Salvador Arijo, and Juan Miguel Mancera. 2021. "Antimicrobial and Toxic Activity of Citronella Essential Oil (Cymbopogon nardus), and Its Effect on the Growth and Metabolism of Gilthead Seabream (Sparus aurata L.)" Fishes 6, no. 4: 61. https://doi.org/10.3390/fishes6040061
APA StyleGarcía-Márquez, J., Barany, A., Ruiz, Á. B., Costas, B., Arijo, S., & Mancera, J. M. (2021). Antimicrobial and Toxic Activity of Citronella Essential Oil (Cymbopogon nardus), and Its Effect on the Growth and Metabolism of Gilthead Seabream (Sparus aurata L.). Fishes, 6(4), 61. https://doi.org/10.3390/fishes6040061