
Deep Heat: A Comparison of Water Temperature, Anemone Bleaching, Anemonefish Density and Reproduction between Shallow and Mesophotic Reefs



Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
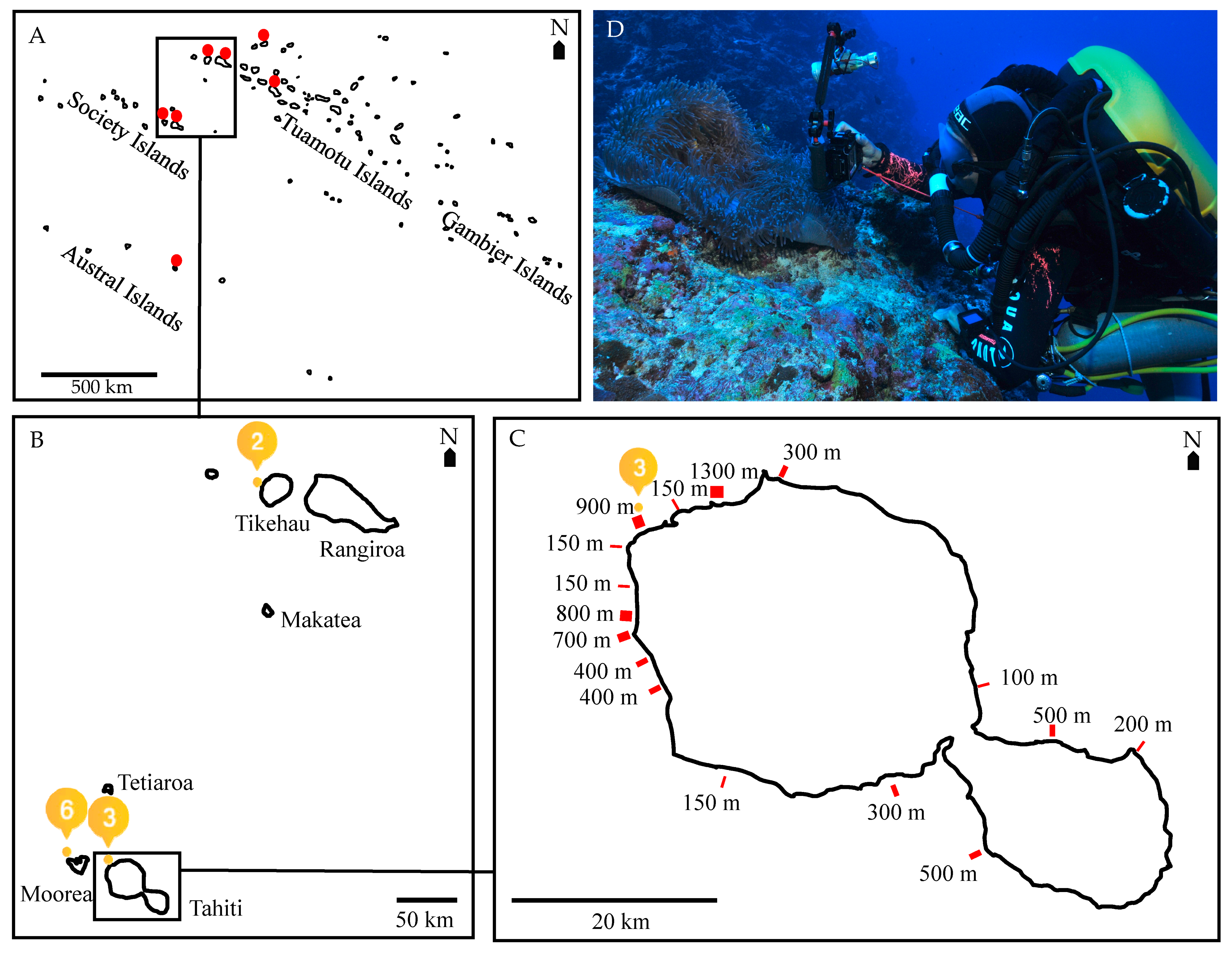
2.1. Temperature Measurements at Shallow and Mesophotic Reefs
2.2. Anemone and Anemonefish Surveys: Anemone Bleaching and Anemonefish Presence, Density Estimates and Reproduction at Shallow and Mesophotic Reefs
3. Results
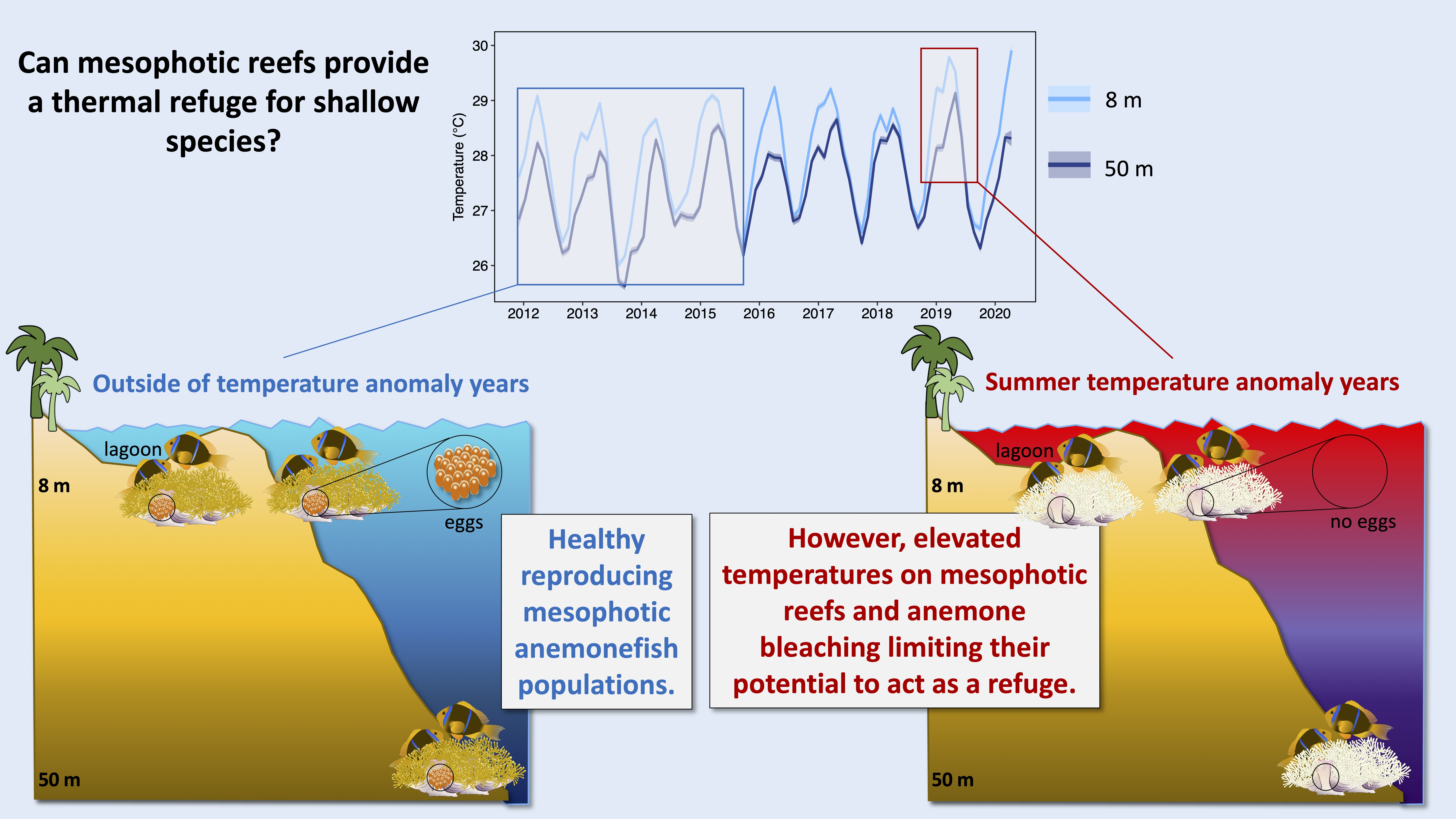
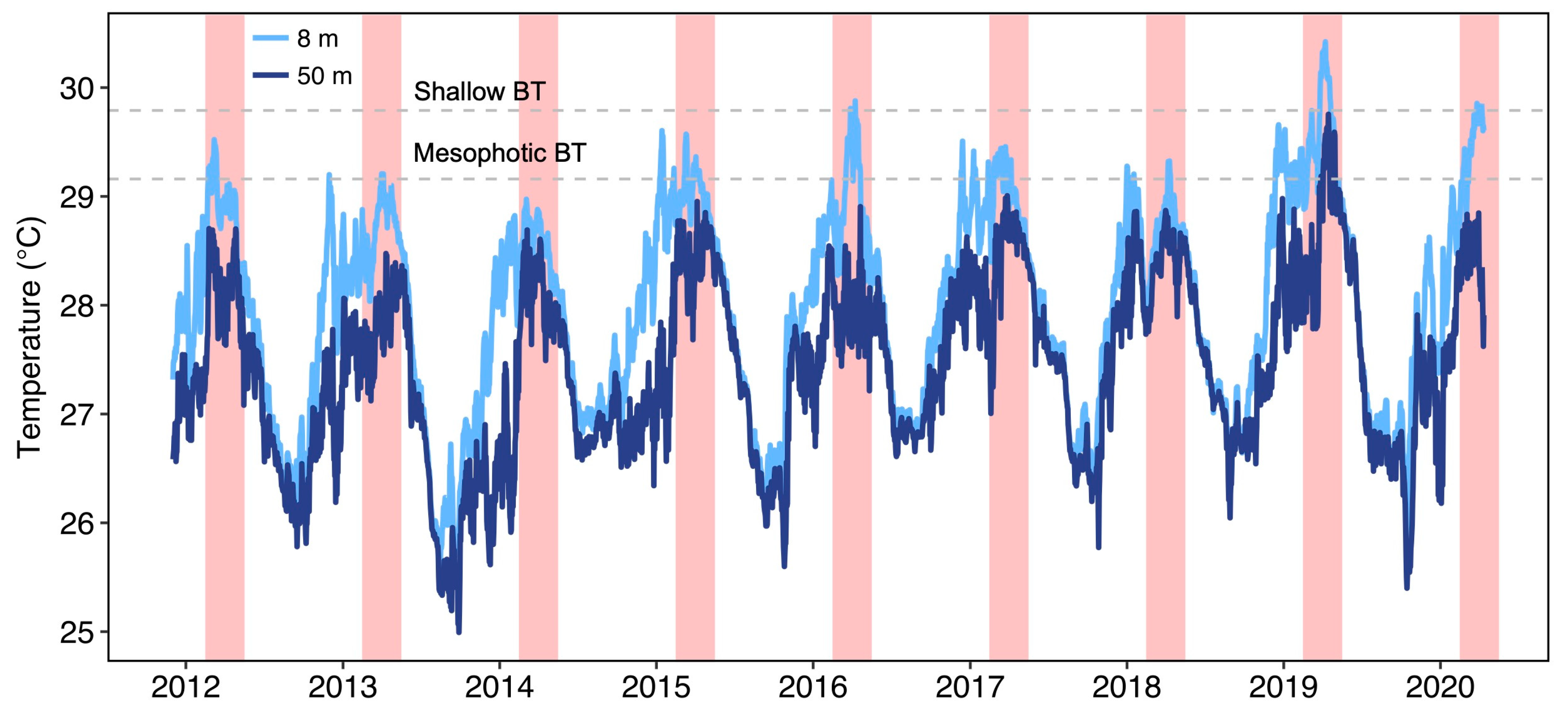
3.1. Comparison of Temperature Measurements between Shallow and Mesophotic Reefs
3.2. Comparison of Anemonefish Presence and Densities between Shallow and Mesophotic Reefs
3.3. Comparison of Host Anemone Bleaching between Shallow and Mesophotic Reefs
3.4. Comparison of Reproductive Events between Shallow and Mesophotic Reefs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pandolfi, J.M.; Bradbury, R.H.; Sala, E.; Hughes, T.P.; Bjorndal, K.A.; Cooke, R.G.; McArdle, D.; McClenachan, L.; Newman, M.J.H.; Paredes, G.; et al. Global trajectories of the long-term decline of coral reef ecosystems. Science 2003, 301, 955–958. [Google Scholar] [CrossRef]
- Hoegh-Guldberg, O.; Mumby, P.J.; Hooten, A.J.; Steneck, R.S.; Greenfield, P.; Gomez, E.; Harvell, C.D.; Sale, P.F.; Edwards, A.J.; Caldeira, K.; et al. Coral reefs under rapid climate change and ocean acidification. Science 2007, 318, 1737–1742. [Google Scholar] [CrossRef]
- Spalding, M.D.; Brown, B.E. Warm-water coral reefs and climate change. Science 2015, 350, 769–771. [Google Scholar] [CrossRef] [PubMed]
- Hughes, T.P.; Kerry, J.T.; Álvarez-Noriega, M.; Álvarez-Romero, J.G.; Anderson, K.D.; Baird, A.H.; Babcock, R.C.; Beger, M.; Bellwood, D.R.; Berkelmans, R.; et al. Global warming and recurrent mass bleaching of corals. Nature 2017, 543, 373–377. [Google Scholar] [CrossRef]
- Hughes, T.P.; Anderson, K.D.; Connolly, S.R.; Heron, S.F.; Kerry, J.T.; Lough, J.M.; Baird, A.H.; Baum, J.K.; Berumen, M.L.; Bridge, T.C.; et al. Spatial and temporal patterns of mass bleaching of corals in the Anthropocene. Science 2018, 359, 80–83. [Google Scholar] [CrossRef] [PubMed]
- Smale, D.A.; Wernberg, T.; Oliver, E.C.J.; Thomsen, M.; Harvey, B.P.; Straub, S.C.; Burrows, M.T.; Alexander, L.V.; Benthuysen, J.A.; Donat, M.G.; et al. Marine heatwaves threaten global biodiversity and the provision of ecosystem services. Nat. Clim. Chang. 2019, 9, 306–312. [Google Scholar] [CrossRef]
- Hoey, A.S.; Howells, E.; Johansen, J.L.; Hobbs, J.-P.A.; Messmer, V.; McCowan, D.M.; Wilson, S.K.; Pratchett, M.S. Recent advances in understanding the effects of climate change on coral reefs. Diversity 2016, 8, 12. [Google Scholar] [CrossRef]
- Hughes, T.P.; Kerry, J.T.; Baird, A.H.; Connolly, S.R.; Dietzel, A.; Eakin, C.M.; Heron, S.F.; Hoey, A.S.; Hoogenboom, M.O.; Liu, G.; et al. Global warming transforms coral reef assemblages. Nature 2018, 556, 492–496. [Google Scholar] [CrossRef] [PubMed]
- Glynn, P.W. Coral reef bleaching: Facts, hypotheses and implications. Glob. Chang. Biol. 1996, 2, 495–509. [Google Scholar] [CrossRef]
- Bridge, T.; Scott, A.; Steinberg, D. Abundance and diversity of anemonefishes and their host sea anemones at two mesophotic sites on the Great Barrier Reef, Australia. Coral Reefs 2012, 31, 1057–1062. [Google Scholar] [CrossRef]
- Muir, P.R.; Marshall, P.A.; Abdulla, A.; Aguirre, J.D. Species identity and depth predict bleaching severity in reef-building corals: Shall the deep inherit the reef? Proc. R. Soc. B Biol. Sci. 2017, 284, 20171551. [Google Scholar] [CrossRef]
- Slattery, M.; Lesser, M.P.; Brazeau, D.; Stokes, M.D.; Leichter, J.J. Connectivity and stability of mesophotic coral reefs. J. Exp. Mar. Bio. Ecol. 2011, 408, 32–41. [Google Scholar] [CrossRef]
- Kahng, S.E.; Copus, J.M.; Wagner, D. Recent advances in the ecology of mesophotic coral ecosystems (MCEs). Curr. Opin. Environ. Sustain. 2014, 7, 72–81. [Google Scholar] [CrossRef]
- Kahng, S.E.; Garcia-Sais, J.R.; Spalding, H.L.; Brokovich, E.; Wagner, D.; Weil, E.; Hinderstein, L.; Toonen, R.J. Community ecology of mesophotic coral reef ecosystems. Coral Reefs 2010, 29, 225–275. [Google Scholar] [CrossRef]
- Reed, J. Deepest Distribution of Atlantic Hermatypic Corals Discovered in the Bahamas. In Proceedings of the Fifth International Coral Reef Congress, French Polynesia, France, 27 May–1 June 1985; Antenne Museum-EPHE: Moorea, French Polynesia, France, 1985; pp. 249–254. [Google Scholar]
- Liddell, W.D.; Ohlhorst, S.L. Hard substrata community patterns, 1–120 m, North Jamaica. Palaios 1988, 3, 413. [Google Scholar] [CrossRef]
- Weiss, K.R. Can deep reefs rescue shallow ones? Science 2017, 355, 903. [Google Scholar] [CrossRef] [PubMed]
- Rocha, L.A.; Pinheiro, H.T.; Shepherd, B.; Papastamatiou, Y.P.; Luiz, O.J.; Pyle, R.L.; Bongaerts, P. Mesophotic coral ecosystems are threatened and ecologically distinct from shallow water reefs. Science 2018, 361, 281–284. [Google Scholar] [CrossRef]
- Hinderstein, L.M.; Marr, J.C.A.; Martinez, F.A.; Dowgiallo, M.J.; Puglise, K.A.; Pyle, R.L.; Zawada, D.G.; Appeldoorn, R. Theme section on “Mesophotic Coral Ecosystems: Characterization, Ecology, and Management. ” Coral Reefs 2010, 29, 247–251. [Google Scholar] [CrossRef]
- Lindfield, S.J.; Harvey, E.S.; Halford, A.R.; McIlwain, J.L. Mesophotic depths as refuge areas for fishery-targeted species on coral reefs. Coral Reefs 2016, 35, 125–137. [Google Scholar] [CrossRef]
- Andradi-Brown, D.A.; Beer, A.J.E.; Colin, L.; Hastuti; Head, C.E.I.; Hidayat, N.I.; Lindfield, S.J.; Mitchell, C.R.; Pada, D.N.; Piesinger, N.M.; et al. Highly diverse mesophotic reef fish communities in Raja Ampat, West Papua. Coral Reefs 2021, 40, 111–130. [Google Scholar] [CrossRef]
- Asher, J.; Williams, I.D.; Harvey, E.S. An assessment of mobile predator populations along shallow and mesophotic depth gradients in the Hawaiian archipelago. Sci. Rep. 2017, 7, 3905. [Google Scholar] [CrossRef] [PubMed]
- Bongaerts, P.; Ridgway, T.; Sampayo, E.M.; Hoegh-Guldberg, O. Assessing the “deep reef refugia” hypothesis: Focus on Caribbean reefs. Coral Reefs 2010, 29, 309–327. [Google Scholar] [CrossRef]
- Smith, T.B.; Glynn, P.W.; Maté, J.L.; Toth, L.T.; Gyory, J. A depth refugium from catastrophic coral bleaching prevents regional extinction. Ecology 2014, 95, 1663–1673. [Google Scholar] [CrossRef] [PubMed]
- Turner, J.A.; Andradi-Brown, D.A.; Gori, A.; Bongaerts, P.; Burdett, H.L.; Ferrier-Pagès, C.; Voolstra, C.R.; Weinstein, D.K.; Bridge, T.C.L.; Costantini, F.; et al. Key Questions for Research and Conservation of Mesophotic Coral Ecosystems and Temperate Mesophotic Ecosystems. In Mesophotic Coral Ecosystems; Springer: Cham, Switzerland, 2019. [Google Scholar]
- Frade, P.R.; Bongaerts, P.; Englebert, N.; Rogers, A.; Gonzalez-Rivero, M.; Hoegh-Guldberg, O. Deep reefs of the Great Barrier Reef offer limited thermal refuge during mass coral bleaching. Nat. Commun. 2018, 9, 1–8. [Google Scholar] [CrossRef]
- Welch, D. Book Review: Climate Change as Social Drama: Global Warming in the Public Sphere. Sociol. Rev. 2016, 64, 387–390. [Google Scholar] [CrossRef]
- Laverick, J.H.; Rogers, A.D. Experimental evidence for reduced mortality of Agaricia lamarcki on a mesophotic reef. Mar. Environ. Res. 2018, 134, 37–43. [Google Scholar] [CrossRef]
- Nir, O.; Gruber, D.F.; Shemesh, E.; Glasser, E.; Tchernov, D. Seasonal mesophotic coral bleaching of Stylophora pistillata in the northern Red Sea. PLoS ONE 2014, 9, e84968. [Google Scholar] [CrossRef]
- Eyal, G.; Tamir, R.; Kramer, N.; Eyal-Shaham, L.; Loya, Y. The Red Sea: Israel. In Mesophotic Coral Ecosystems; Springer: Cham, Switzerland, 2019. [Google Scholar]
- Smith, T.B.; Gyory, J.; Brandt, M.E.; Miller, W.J.; Jossart, J.; Nemeth, R.S. Caribbean mesophotic coral ecosystems are unlikely climate change refugia. Glob. Chang. Biol. 2016, 22, 2759–2765. [Google Scholar] [CrossRef]
- Brokovich, E.; Ayalon, I.; Einbinder, S.; Segev, N.; Shaked, Y.; Genin, A.; Kark, S.; Kiflawi, M. Grazing pressure on coral reefs decreases across a wide depth gradient in the Gulf of Aqaba, Red Sea. Mar. Ecol. Prog. Ser. 2010, 399, 69–80. [Google Scholar] [CrossRef]
- Malcolm, H.A.; Jordan, A.; Smith, S.D.A. Testing a depth-based habitat classification system against reef fish assemblage patterns in a subtropical marine park. Aquat. Conserv. Mar. Freshw. Ecosyst. 2011, 21, 173–185. [Google Scholar] [CrossRef]
- Feitoza, B.M.; Rosa, R.S.; Rocha, L.A. Ecology and zoogeography of deep-reef fishes in northeastern Brazil. Bull. Mar. Sci. 2005, 76, 725–742. [Google Scholar]
- Pearson, R.; Stevens, T. Distinct cross-shelf gradient in mesophotic reef fish assemblages in subtropical eastern Australia. Mar. Ecol. Prog. Ser. 2015, 532, 185–196. [Google Scholar] [CrossRef]
- Coleman, R.R.; Copus, J.M.; Coffey, D.M.; Whitton, R.K.; Bowen, B.W. Shifting reef fish assemblages along a depth gradient in Pohnpei, Micronesia. PeerJ 2018, 6, e4650. [Google Scholar] [CrossRef]
- Tenggardjaja, K.A.; Bowen, B.W.; Bernardi, G. Vertical and horizontal genetic connectivity in Chromis verater, an endemic damselfish found on shallow and mesophotic reefs in the Hawaiian archipelago and adjacent Johnston atoll. PLoS ONE 2014, 9, e115493. [Google Scholar] [CrossRef]
- Van Oppen, M.J.H.; Bongaerts, P.; Underwood, J.N.; Peplow, L.M.; Cooper, T.F. The role of deep reefs in shallow reef recovery: An assessment of vertical connectivity in a brooding coral from west and east Australia. Mol. Ecol. 2011, 20, 1647–1660. [Google Scholar] [CrossRef] [PubMed]
- Serrano, X.M.; Baums, I.B.; O’Reilly, K.; Smith, T.B.; Jones, R.J.; Shearer, T.L.; Nunes, F.L.D.; Baker, A.C. Geographic differences in vertical connectivity in the Caribbean coral Montastraea cavernosa despite high levels of horizontal connectivity at shallow depths. Mol. Ecol. 2014, 23, 4226–4240. [Google Scholar] [CrossRef] [PubMed]
- Shlesinger, T.; Grinblat, M.; Rapuano, H.; Amit, T.; Loya, Y. Can mesophotic reefs replenish shallow reefs? Reduced coral reproductive performance casts a doubt. Ecology 2018, 99, 421–437. [Google Scholar] [CrossRef]
- Holstein, D.M.; Paris, C.B.; Vaz, A.C.; Smith, T.B. Modeling vertical coral connectivity and mesophotic refugia. Coral Reefs 2016, 35, 23–37. [Google Scholar] [CrossRef]
- Mariscal, R.N.; Fautin, D.G.; Allen, G.R. Field Guide to Anemonefishes and Their Host Sea Anemones. Copeia 1993, 1993. [Google Scholar] [CrossRef]
- Norin, T.; Mills, S.C.; Crespel, A.; Cortese, D.; Killen, S.S.; Beldade, R. Anemone bleaching increases the metabolic demands of symbiont anemonefish. Proc. R. Soc. B Biol. Sci. 2018, 285, 20180282. [Google Scholar] [CrossRef]
- Beldade, R.; Blandin, A.; O’Donnell, R.; Mills, S.C. Cascading effects of thermally-induced anemone bleaching on associated anemonefish hormonal stress response and reproduction. Nat. Commun. 2017, 8, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Cortese, D.; Norin, T.; Beldade, R.; Crespel, A.; Killen, S.S.; Mills, S.C. Physiological and behavioural effects of anemone bleaching on symbiont anemonefish in the wild. Funct. Ecol. 2021, 35, 663–674. [Google Scholar] [CrossRef]
- Leray, M.; Beldade, R.; Holbrook, S.J.; Schmitt, R.J.; Planes, S.; Bernardi, G. Allopatric divergence and speciation in coral reef fish: The three-spot dascyllus, Dascyllus trimaculatus, species complex. Evolution 2010, 64, 1218–1230. [Google Scholar] [CrossRef]
- Brokovich, E.; Einbinder, S.; Shashar, N.; Kiflawi, M.; Kark, S. Descending to the twilight-zone: Changes in coral reef fish assemblages along a depth gradient down to 65 m. Mar. Ecol. Prog. Ser. 2008, 371, 253–262. [Google Scholar] [CrossRef]
- Steinberg, R.K.; van der Meer, M.H.; Pratchett, M.S.; van Herwerden, L.; Hobbs, J.-P.A. Keep your friends close and your anemones closer—Ecology of the endemic wideband anemonefish, Amphiprion latezonatus. Environ. Biol. Fishes 2020, 103, 1513–1526. [Google Scholar] [CrossRef]
- Kühlmann, D.H.H. Composition and ecology of deep-water coral associations. Helgoländer Meeresunters. 1983, 36, 183–204. [Google Scholar] [CrossRef]
- Pyle, R.L. Assessing undiscovered fish biodiversity on deep coral reefs using advanced self-contained diving technology. Mar. Technol. Soc. J. 2000, 34, 82–91. [Google Scholar] [CrossRef]
- Pyle, R.L. Exploring deep coral reefs: How much biodiversity are we missing? Glob. Biodivers. 1996, 6, 3–7. [Google Scholar]
- Sieber, A.; Pyle, R. A review of the use of closed-circuit rebreathers for scientific diving. Underw. Technol. 2010, 29, 73–78. [Google Scholar] [CrossRef]
- Pinheiro, H.T.; Mazzei, E.; Moura, R.L.; Amado-Filho, G.M.; Carvalho-Filho, A.; Braga, A.C.; Costa, P.A.S.; Ferreira, B.P.; Ferreira, C.E.L.; Floeter, S.R.; et al. Fish biodiversity of the Vitória-Trindade seamount chain, southwestern Atlantic: An updated database. PLoS ONE 2015, 10, e0118180. [Google Scholar] [CrossRef]
- Liu, G.; Strong, A.; Skirving, W.; Arzayus, F. Overview of NOAA Coral Reef Watch Program’s Near-Real Time Satellite Global Coral Bleaching Monitoring Activities. In Proceedings of the 10th International Coral Reef Symposium, Okinawa, Japan, 28 June–2 July 2003; Gurugram: Okinawa, Japan, 2006; Volume 1793, pp. 1783–1793. [Google Scholar]
- Burgess, S.C.; Johnston, E.C.; Wyatt, A.S.J.; Leichter, J.J.; Edmunds, P.J. Response diversity in corals: Hidden differences in bleaching mortality among cryptic Pocillopora species. Ecology 2021, 102, e03324. [Google Scholar] [CrossRef]
- Seymour, J.R.; Barbasch, T.A.; Buston, P.M. Lunar cycles of reproduction in the clown anemonefish Amphiprion percula: Individual-level strategies and population-level patterns. Mar. Ecol. Prog. Ser. 2018, 594, 193–201. [Google Scholar] [CrossRef]
- Pryor, S.H.; Andrews, L.; Kelaher, B.P.; Tagliafico, A.; Scott, A. Ocean temperature, but not acidification, causes sea anemone bleaching under a near-future climate scenario. Coral Reefs 2021, 40, 355–364. [Google Scholar] [CrossRef]
- Tada, K.; Sakai, K.; Nakano, Y.; Takemura, A.; Montani, S. Size-fractionated phytoplankton biomass in coral reef waters off Sesoko Island, Okinawa, Japan. J. Plankton Res. 2003, 25, 991–997. [Google Scholar] [CrossRef]
- Beldade, R.; Holbrook, S.J.; Schmitt, R.J.; Planes, S.; Malone, D.; Bernardi, G. Larger female fish contribute disproportionately more to self-replenishment. Proc. R. Soc. B Biol. Sci. 2012, 279, 2116–2121. [Google Scholar] [CrossRef] [PubMed]
- Beldade, R.; Holbrook, S.J.; Schmitt, R.J.; Planes, S.; Bernardi, G. Spatial patterns of self-recruitment of a coral reef fish in relation to island-scale retention mechanisms. Mol. Ecol. 2016, 25, 5203–5211. [Google Scholar] [CrossRef]
- Emms, M.A.; Saenz-Agudelo, P.; Giles, E.C.; Gatins, R.; Nanninga, G.B.; Scott, A.; Hobbs, J.P.A.; Frisch, A.J.; Mills, S.C.; Beldade, R.; et al. Comparative phylogeography of three host sea anemones in the Indo-Pacific. J. Biogeogr. 2020, 47, 487–500. [Google Scholar] [CrossRef]
- Dixon, A.K.; Needham, D.; Al-Horani, F.A.; Chadwick, N.E. Microhabitat use and photoacclimation in the clownfish sea anemone Entacmaea quadricolor. J. Mar. Biol. Assoc. UK 2014, 94, 473–480. [Google Scholar] [CrossRef]
- MacDonald, C.; Bridge, T.C.L.; Jones, G.P. Depth, bay position and habitat structure as determinants of coral reef fish distributions: Are deep reefs a potential refuge? Mar. Ecol. Prog. Ser. 2016, 561, 217–231. [Google Scholar] [CrossRef]
- Rouzé, H.; Galand, P.E.; Medina, M.; Bongaerts, P.; Pichon, M.; Pérez-Rosales, G.; Torda, G.; Moya, A.; Bardout, G.; Périé-Bardout, E.; et al. Symbiotic associations of the deepest recorded photosynthetic scleractinian coral (172 m depth). ISME J. 2021, 15, 1564–1568. [Google Scholar] [CrossRef]
- Goldstein, E.D.; D’Alessandro, E.K.; Sponaugle, S. Demographic and reproductive plasticity across the depth distribution of a coral reef fish. Sci. Rep. 2016, 6, 34077. [Google Scholar] [CrossRef] [PubMed]
- Favorite, F.; Laevastu, T.; Straty, R. Oceanography of the Northestern Pacific Ocean and Eastern Bering Sea and Relations to Various Living Marine Resources; National Marine Fisheries Service, Northwest and Alaska Fisheries Center: Washington, WA, USA, 1977. [Google Scholar]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Haguenauer, A.; Zuberer, F.; Siu, G.; Cortese, D.; Beldade, R.; Mills, S.C. Deep Heat: A Comparison of Water Temperature, Anemone Bleaching, Anemonefish Density and Reproduction between Shallow and Mesophotic Reefs. Fishes 2021, 6, 37. https://doi.org/10.3390/fishes6030037
Haguenauer A, Zuberer F, Siu G, Cortese D, Beldade R, Mills SC. Deep Heat: A Comparison of Water Temperature, Anemone Bleaching, Anemonefish Density and Reproduction between Shallow and Mesophotic Reefs. Fishes. 2021; 6(3):37. https://doi.org/10.3390/fishes6030037
Chicago/Turabian StyleHaguenauer, Anne, Frédéric Zuberer, Gilles Siu, Daphne Cortese, Ricardo Beldade, and Suzanne C. Mills. 2021. "Deep Heat: A Comparison of Water Temperature, Anemone Bleaching, Anemonefish Density and Reproduction between Shallow and Mesophotic Reefs" Fishes 6, no. 3: 37. https://doi.org/10.3390/fishes6030037
APA StyleHaguenauer, A., Zuberer, F., Siu, G., Cortese, D., Beldade, R., & Mills, S. C. (2021). Deep Heat: A Comparison of Water Temperature, Anemone Bleaching, Anemonefish Density and Reproduction between Shallow and Mesophotic Reefs. Fishes, 6(3), 37. https://doi.org/10.3390/fishes6030037