
Morphological and Metabolic Adaptations to Increasing Temperature: Insights from the In Vitro Model of Maraena Whitefish


Abstract
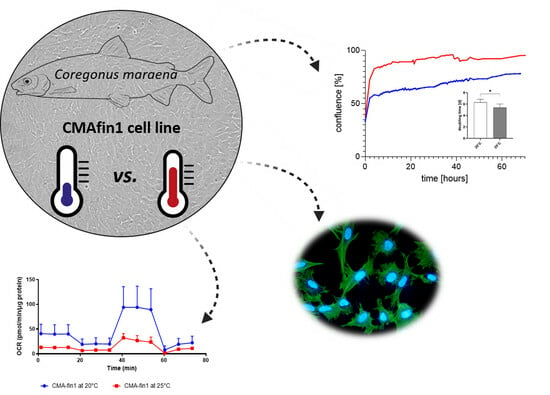
1. Introduction
2. Materials and Methods
2.1. Cell Cultivation
2.2. Temperature Adaptation
2.3. Phase Contrast Pictures
2.4. Immunofluorescent Staining
2.5. Growth Analysis
2.6. Analysis of Metabolic Function
2.7. Gene Expression Analyses
2.8. Statistical Analysis
3. Results
3.1. Morphology Changes and Growth Analysis of Temperature-Adapted CMAfin1 Cells
3.2. Mitochondrial Oxidative Respiration
3.3. Metabolic Phenotype
3.4. Glycolytic Activity
3.5. Gene Expression
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ATP | Adenosine triphosphate |
| CT | Cycle threshold |
| DAPI | 4′,6-diamidino-2-phenylindole |
| ECAR | Extracellular acidification rate |
| FBS | Foetal Bovine Serum |
| FCCP | Carbonyl cyanide-p-trifluoromethoxyphenylhydrazone |
| GOI | Genes of interest |
| L-15 | Leibovitz-15 |
| IUCN | International Union for Conservation of Nature and Natural Resources |
| OCR | Oxygen consumption rate |
| PBS | Dulbecco’s Phosphate-buffered Saline |
| Ro/AA | Rotenone and antimycin A |
References
- IPCC. Climate Change 2023: Synthesis Report; Contribution of Working Groups I, II and III to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; IPCC: Geneva, Switzerland, 2023; 184p. [Google Scholar] [CrossRef]
- Morée, A.L.; Clarke, T.M.; Cheung, W.W.L.; Frölicher, T.L. Impact of deoxygenation and warming on global marine species in the 21st century. Biogeosciences 2023, 20, 2425–2454. [Google Scholar] [CrossRef]
- Nimma, D.; Devi, O.R.; Laishram, B.; Ramesh, J.V.N.; Boddupalli, S.; Ayyasamy, R.; Tirth, V.; Arabil, A. Implications of climate change on freshwater ecosystems and their biodiversity. Desalin. Water Treat. 2025, 321, 100889. [Google Scholar] [CrossRef]
- Meier, H.E.M.; Kniebusch, M.; Dieterich, C.; Gröger, M.; Zorita, E.; Elmgren, R.; Myrberg, K.; Ahola, M.P.; Bartosova, A.; Bonsdorff, E.; et al. Climate change in the Baltic Sea region: A summary. Earth Syst. Dynam. 2022, 13, 457–593. [Google Scholar] [CrossRef]
- Reusch, T.B.H.; Dierking, J.; Andersson, H.C.; Bonsdorff, E.; Carstensen, J.; Casini, M.; Czajkowski, M.; Hasler, B.; Hinsby, K.; Hyytiäinen, K.; et al. The Baltic Sea as a time machine for the future coastal ocean. Sci. Adv. 2018, 4, eaar8195. [Google Scholar] [CrossRef] [PubMed]
- Hellmann, J.J.; Byers, J.E.; Bierwagen, B.G.; Dukes, J.S. Five Potential Consequences of Climate Change for Invasive Species. Conserv. Biol. 2008, 22, 534–543. [Google Scholar] [CrossRef] [PubMed]
- Viitasalo, M.; Bonsdorff, E. Global climate change and the Baltic Sea ecosystem: Direct and indirect effects on species, communities and ecosystem functioning. Earth Syst. Dynam. 2022, 13, 711–747. [Google Scholar] [CrossRef]
- Freyhof, J. Coregonus maraena. In The IUCN Red List of Threatened Species 2013; IUCN: Gland, Switzerland, 2013. [Google Scholar] [CrossRef]
- Ford, M. Coregonus maraena. In The IUCN Red List of Threatened Species 2024; IUCN: Gland, Switzerland, 2024. [Google Scholar] [CrossRef]
- Rebl, A.; Verleih, M.; Nipkow, M.; Altmann, S.; Bochert, R.; Goldammer, T. Gradual and Acute Temperature Rise Induces Crossing Endocrine, Metabolic, and Immunological Pathways in Maraena Whitefish (Coregonus maraena). Front. Genet. 2018, 9, 241. [Google Scholar] [CrossRef] [PubMed]
- Bochert, R.; Luft, P.; Gebhard, R. Temperature preferences of three maraena whitefish (Coregonus maraena) size classes under aquaculture conditions. Aquac. Res. 2020, 51, 2160–2163. [Google Scholar] [CrossRef]
- Edsall, T.A.; Rottiers, D.V.; Brown, E.H. Temperature tolerance of bloater (Coregonus hoyi). J. Fish. Res. Board Can. 1970, 27, 2047–2052. [Google Scholar] [CrossRef]
- Edsall, T.A.; Rottiers, D.V. Temperature tolerance of young-of-the-year lake whitefish, Coregonus clupeaformis. J. Fish. Res. Board Can. 1976, 33, 177–180. [Google Scholar] [CrossRef]
- McCormick, J.H.; Jones, B.R.; Syrett, R.F. Temperature Requirements for Growth and Survival of Larval Ciscos (Coregonus artedii). J. Fish. Board Can. 1971, 28, 924–927. [Google Scholar] [CrossRef]
- Alfonso, S.; Gesto, M.; Sadoul, B. Temperature increase and its effects on fish stress physiology in the context of global warming. J. Fish Biol. 2021, 98, 1496–1508. [Google Scholar] [CrossRef] [PubMed]
- Little, A.G.; Loughland, I.; Seebacher, F. What do warming waters mean for fish physiology and fisheries? J. Fish Biol. 2020, 97, 328–340. [Google Scholar] [CrossRef] [PubMed]
- Cingi, S.; Keinänen, M.; Vuorinen, P.J. Elevated water temperature impairs fertilization and embryonic development of whitefish Coregonus lavaretus. J. Fish Biol. 2010, 76, 502–521. [Google Scholar] [CrossRef] [PubMed]
- Korytar, T.; Nipkow, M.; Altmann, S.; Goldammer, T.; Kollner, B.; Rebl, A. Adverse Husbandry of Maraena Whitefish Directs the Immune System to Increase Mobilization of Myeloid Cells and Proinflammatory Responses. Front. Immunol. 2016, 7, 631. [Google Scholar] [CrossRef] [PubMed]
- Kern, P.; Cramp, R.L.; Franklin, C.E. Physiological responses of ectotherms to daily temperature variation. J. Exp. Biol. 2015, 218, 3068–3076. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.S.; Yoon, S.-J.; Kim, J.M.; Gil, J.W.; Lee, T.W. Effects of temperature changes on the endogenous rhythm of oxygen consumption in the Japanese flounder Paralichthys olivaceus. Fish. Sci. 2005, 71, 471–478. [Google Scholar] [CrossRef]
- Hildebrandt, J.-P.; Bleckmann, H.; Homberg, U. Penzlin—Lehrbuch der Tierphysiologie, 9th ed.; Springer Spektrum: Berlin/Heidelberg, Germany, 2021. [Google Scholar]
- Brett, J.R. Energetic Responses of Salmon to Temperature. A Study of Some Thermal Relations in the Physiology and Freshwater Ecology of Sockeye Salmon (Oncorhynchus nerkd). Am. Zool. 1971, 11, 99–113. [Google Scholar] [CrossRef]
- Reynolds, W.W.; Casterlin, M.E. The Role of Temperature in the Environmental Physiology of Fishes; Springer: Boston, MA, USA, 1980. [Google Scholar]
- Morgan, M.J. Low-Temperature Tolerance of American Plaice in Relation to Declines in Abundance. Trans. Am. Fish. Soc. 1992, 121, 399–402. [Google Scholar] [CrossRef]
- Morgan, M.J.; Rideout, R.M.; Colbourne, E.B. Impact of environmental temperature on Atlantic cod Gadus morhua energy allocation to growth, condition and reproduction. Mar. Ecol. Progress. Ser. 2010, 404, 185–195. [Google Scholar] [CrossRef]
- Van Dijk, P.L.M.; Tesch, C.; Hardewig, I.; Pörtner, H.O. Physiological disturbances at critically high temperatures: A comparison between stenothermal antarctic and eurythermal temperate eelpouts (Zoarcidae). J. Exp. Biol. 1999, 202, 3611–3621. [Google Scholar] [CrossRef] [PubMed]
- Evans, D.O. Metabolic Thermal Compensation by Rainbow Trout: Effects on Standard Metabolic Rate and Potential Usable Power. Trans. Am. Fish. Soc. 1990, 119, 585–600. [Google Scholar] [CrossRef]
- Gerber, L.; Clow, K.A.; Gamperl, A.K. Acclimation to warm temperatures has important implications for mitochondrial function in Atlantic salmon (Salmo salar). J. Exp. Biol. 2021, 224, jeb236257. [Google Scholar] [CrossRef] [PubMed]
- Bols, N.C.; Mosser, D.D.; Steels, G.B. Temperature studies and recent advances with fish cells in vitro. Comp. Biochem. Physiol. Part A Physiol. 1992, 103, 1–14. [Google Scholar] [CrossRef]
- Cui, Y.; Liu, B.; Xie, J.; Xu, P.; Tsion, H.M.H.; Zhang, Y. The effect of hyperthermia on cell viability, oxidative damage, and heat shock protein expression in hepatic cells of grass carp (Ctenopharyngodon idellus). J. Therm. Biol. 2013, 38, 355–361. [Google Scholar] [CrossRef]
- Bloch, S.R.; Vo, N.T.K.; Walsh, S.K.; Chen, C.; Lee, L.E.J.; Hodson, P.V.; Bols, N.C. Development of a cell line from the American eel brain expressing endothelial cell properties. In Vitr. Cell. Dev. Biol.-Anim. 2016, 52, 395–409. [Google Scholar] [CrossRef] [PubMed]
- Lutze, P.; Brenmoehl, J.; Tesenvitz, S.; Ohde, D.; Wanka, H.; Meyer, Z.; Grunow, B. Effects of Temperature Adaptation on the Metabolism and Physiological Properties of Sturgeon Fish Larvae Cell Line. Cells 2024, 13, 269. [Google Scholar] [CrossRef] [PubMed]
- Grunow, B.; Franz, G.P.; Tönißen, K. In Vitro Fish Models for the Analysis of Ecotoxins and Temperature Increase in the Context of Global Warming. Toxics 2021, 9, 286. [Google Scholar] [CrossRef] [PubMed]
- Kaya, Y.; Tönißen, K.; Verleih, M.; Rebl, H.; Grunow, B. Establishment of an in vitro model from the vulnerable fish species Coregonus maraena (maraena whitefish): Optimization of growth conditions and characterization of the cell line. Cell Biol. Int. 2022, 47, 548–559. [Google Scholar] [CrossRef] [PubMed]
- Goswami, M.; Yashwanth, B.S.; Trudeau, V.; Lakra, W.S. Role and relevance of fish cell lines in advanced in vitro research. Mol. Biol. Rep. 2022, 49, 2393–2411. [Google Scholar] [CrossRef] [PubMed]
- Grunow, B.; Strauch, S.M. Status assessment and opportunities for improving fish welfare in animal experimental research according to the 3R-Guidelines. Rev. Fish Biol. Fish. 2023, 33, 1075–1093. [Google Scholar] [CrossRef]
- Rehberger, K.; Kropf, C.; Segner, H. In vitro or not in vitro: A short journey through a long history. Environ. Sci. Eur. 2018, 30, 23. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.V.; Kumar, A.; Trang, P.N. The use of fish cell lines as in-vitro ecotoxicological tools: A cellular solution to aquaculture sustainability. Aquaculture 2024, 593, 741302. [Google Scholar] [CrossRef]
- Espinosa-Ruiz, C.; Mayor-Lafuente, J.; Esteban, M.Á. Mitochondrial Metabolism Characterization of Four Different Fish Cell Lines. Fishes 2022, 7, 354. [Google Scholar] [CrossRef]
- Robins, H.I.; Steeves, R.A.; Clark, A.W.; Martin, P.A.; Miller, K.; Dennis, W.H. Differential sensitivity of AKR murine leukemia and normal bone marrow cells to hyperthermia. Cancer Res. 1983, 43, 4951–4955. [Google Scholar] [CrossRef] [PubMed]
- Lian, P.; Braber, S.; Varasteh, S.; Wichers, H.J.; Folkerts, G. Hypoxia and heat stress affect epithelial integrity in a Caco-2/HT-29 co-culture. Sci. Rep. 2021, 11, 13186. [Google Scholar] [CrossRef] [PubMed]
- Serano, M.; Pietrangelo, L.; Paolini, C.; Guarnier, F.A.; Protasi, F. Oxygen Consumption and Basal Metabolic Rate as Markers of Susceptibility to Malignant Hyperthermia and Heat Stroke. Cells 2022, 11, 2468. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, M.; Liu, C.; Oyama, K.; Yamazawa, T. Trans-scale thermal signaling in biological systems. J. Biochem. 2023, 174, 217–225. [Google Scholar] [CrossRef] [PubMed]
- Gstraunthaler, G.; Lindl, T. Zell- und Gewebekultur, Allgemeine Grundlagen und spezielle Anwendungen; Springer Spektrum: Berlin/Heidelberg, Germany, 2021. [Google Scholar] [CrossRef]
- Grunow, B.; Noglick, S.; Kruse, C.; Gebert, M. Isolation of cells from Atlantic sturgeon Acipenser oxyrinchus oxyrinchus and optimization of culture conditions. Aquat. Biol. 2011, 14, 67–75. [Google Scholar] [CrossRef]
- Ferrick, D.A.; Neilson, A.; Beeson, C. Advances in measuring cellular bioenergetics using extracellular flux. Drug Discov. Today 2008, 13, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Gu, X.; Ma, Y.; Liu, Y.; Wan, Q. Measurement of mitochondrial respiration in adherent cells by Seahorse XF96 Cell Mito Stress Test. STAR Protoc. 2021, 2, 100245. [Google Scholar] [CrossRef] [PubMed]
- Leung, D.T.H.; Chu, S. Measurement of Oxidative Stress: Mitochondrial Function Using the Seahorse System. In Preeclampsia: Methods and Protocols; Murthi, P., Vaillancourt, C., Eds.; Springer: New York, NY, USA, 2018; pp. 285–293. [Google Scholar]
- Divakaruni, A.S.; Paradyse, A.; Ferrick, D.A.; Murphy, A.N.; Jastroch, M. Analysis and interpretation of microplate-based oxygen consumption and pH data. Methods Enzymol. 2014, 547, 309–354. [Google Scholar] [CrossRef] [PubMed]
- Wanka, H.; Lutze, P.; Staar, D.; Grunow, B.; Peters, B.S.; Peters, J. An alternative renin isoform is cardioprotective by modulating mitochondrial metabolism. J. Cell Mol. Med. 2018, 22, 5991–6001. [Google Scholar] [CrossRef] [PubMed]
- Vangapandu, H.V.; Gandhi, V. Extracellular Flux Assays to Determine Oxidative Phosphorylation and Glycolysis in Chronic Lymphocytic Leukemia Cells. In Chronic Lymphocytic Leukemia: Methods and Protocols; Malek, S.N., Ed.; Springer: New York, NY, USA, 2019; pp. 121–128. [Google Scholar]
- Wu, M.; Neilson, A.; Swift, A.L.; Moran, R.; Tamagnine, J.; Parslow, D.; Armistead, S.; Lemire, K.; Orrell, J.; Teich, J.; et al. Multiparameter metabolic analysis reveals a close link between attenuated mitochondrial bioenergetic function and enhanced glycolysis dependency in human tumor cells. Am. J. Physiol. Cell Physiol. 2007, 292, C125–C136. [Google Scholar] [CrossRef] [PubMed]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE guidelines: Minimum information for publication of quantitative real-time PCR experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [PubMed]
- Altmann, S.; Rebl, A.; Kuhn, C.; Goldammer, T. Identification and de novo sequencing of housekeeping genes appropriate for gene expression analyses in farmed maraena whitefish (Coregonus maraena) during crowding stress. Fish Physiol. Biochem. 2015, 41, 397–412. [Google Scholar] [CrossRef] [PubMed]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, research0034.1. [Google Scholar] [CrossRef] [PubMed]
- Morrison, T.B.; Weis, J.J.; Wittwer, C.T. Quantification of low-copy transcripts by continuous SYBR Green I monitoring during amplification. Biotechniques 1998, 24, 954–958, 960, 962. [Google Scholar] [PubMed]
- Freeman, W.M.; Walker, S.J.; Vrana, K.E. Quantitative RT-PCR: Pitfalls and potential. BioTechniques 1999, 26, 112–125. [Google Scholar] [CrossRef] [PubMed]
- Martorell-Ribera, J.; Nipkow, M.; Viergutz, T.; Brunner, R.M.; Bochert, R.; Koll, R.; Goldammer, T.; Gimsa, U.; Rebl, A. Early response of salmonid head-kidney cells to stress hormones and toll-like receptor ligands. Fish Shellfish Immunol. 2020, 98, 950–961. [Google Scholar] [CrossRef] [PubMed]
- Bustin, S.A.; Nolan, T. Pitfalls of quantitative real-time reverse-transcription polymerase chain reaction. J. Biomol. Tech. 2004, 15, 155–166. [Google Scholar] [PubMed]
- Hendriks-Balk, M.C.; Michel, M.C.; Alewijnse, A.E. Pitfalls in the normalization of real-time polymerase chain reaction data. Basic. Res. Cardiol. 2007, 102, 195–197. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Gerber, L.; Clow, K.A.; Mark, F.C.; Gamperl, A.K. Improved mitochondrial function in salmon (Salmo salar) following high temperature acclimation suggests that there are cracks in the proverbial ‘ceiling’. Sci. Rep. 2020, 10, 21636. [Google Scholar] [CrossRef] [PubMed]
- Hickey, A.J.R.; Harford, A.R.; Blier, P.U.; Devaux, J.B. What causes cardiac mitochondrial failure at high environmental temperatures? J. Exp. Biol. 2024, 227, jeb247432. [Google Scholar] [CrossRef] [PubMed]
- Grunow, B.; Stange, K.; Bochert, R.; Tönißen, K. Histological and biochemical evaluation of skeletal muscle in the two salmonid species Coregonus maraena and Oncorhynchus mykiss. PLoS ONE 2021, 16, e0255062. [Google Scholar] [CrossRef] [PubMed]
- Willmer, P.; Stone, G.; Johnston, I. Environmental Physiology of Animals; John Wiley & Sons: Hoboken, NJ, USA, 2009. [Google Scholar]
- Regier, H.A.; Holmes, J.A.; Pauly, D. Influence of Temperature Changes on Aquatic Ecosystems: An Interpretation of Empirical Data. Trans. Am. Fish. Soc. 1990, 119, 374–389. [Google Scholar] [CrossRef]
- Pörtner, H.O.; Peck, M.A. Climate change effects on fishes and fisheries: Towards a cause-and-effect understanding. J. Fish Biol. 2010, 77, 1745–1779. [Google Scholar] [CrossRef] [PubMed]
- Dolfi, S.C.; Chan, L.L.-Y.; Qiu, J.; Tedeschi, P.M.; Bertino, J.R.; Hirshfield, K.M.; Oltvai, Z.N.; Vazquez, A. The metabolic demands of cancer cells are coupled to their size and protein synthesis rates. Cancer Metab. 2013, 1, 20. [Google Scholar] [CrossRef] [PubMed]
- Muranova, L.K.; Shatov, V.M.; Gusev, N.B. Role of Small Heat Shock Proteins in the Remodeling of Actin Microfilaments. Biochemistry 2022, 87, 800–811. [Google Scholar] [CrossRef] [PubMed]
- Mitchison, T.J.; Cramer, L.P. Actin-Based Cell Motility and Cell Locomotion. Cell 1996, 84, 371–379. [Google Scholar] [CrossRef] [PubMed]
- Pollard, T.D.; Borisy, G.G. Cellular Motility Driven by Assembly and Disassembly of Actin Filaments. Cell 2003, 112, 453–465. [Google Scholar] [CrossRef] [PubMed]
- Svitkina, T. The Actin Cytoskeleton and Actin-Based Motility. Cold Spring Harb. Perspect. Biol. 2018, 10, a018267. [Google Scholar] [CrossRef] [PubMed]
- Ng, I.C.; Pawijit, P.; Tan, J.; Yu, H. Anatomy and Physiology for Biomaterials Research and Development. In Encyclopedia of Biomedical Engineering; Narayan, R., Ed.; Elsevier: Oxford, UK, 2019; pp. 225–236. [Google Scholar]
- Buracco, S.; Claydon, S.; Insall, R. Control of actin dynamics during cell motility. F1000Research 2019, 8, 1977. [Google Scholar] [CrossRef] [PubMed]
- Tojkander, S.; Gateva, G.; Lappalainen, P. Actin stress fibers—Assembly, dynamics and biological roles. J. Cell Sci. 2012, 125, 1855–1864. [Google Scholar] [CrossRef] [PubMed]
- Rottner, K.; Faix, J.; Bogdan, S.; Linder, S.; Kerkhoff, E. Actin assembly mechanisms at a glance. J. Cell Sci. 2017, 130, 3427–3435. [Google Scholar] [CrossRef] [PubMed]
- Rupik, W.; Jasik, K.; Bembenek, J.; Widłak, W. The expression patterns of heat shock genes and proteins and their role during vertebrate’s development. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2011, 159, 349–366. [Google Scholar] [CrossRef] [PubMed]
- Stefanovic, D.I.; Manzon, L.A.; McDougall, C.S.; Boreham, D.R.; Somers, C.M.; Wilson, J.Y.; Manzon, R.G. Thermal stress and the heat shock response in embryonic and young of the year juvenile lake whitefish. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2016, 193, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Nipkow, M.; Wirthgen, E.; Luft, P.; Rebl, A.; Hoeflich, A.; Goldammer, T. Characterization of igf1 and igf2 genes during maraena whitefish (Coregonus maraena) ontogeny and the effect of temperature on embryogenesis and igf expression. Growth Horm. IGF Res. 2018, 40, 32–43. [Google Scholar] [CrossRef] [PubMed]
- Manzon, L.A.; Zak, M.A.; Agee, M.; Boreham, D.R.; Wilson, J.Y.; Somers, C.M.; Manzon, R.G. Thermal acclimation alters both basal heat shock protein gene expression and the heat shock response in juvenile lake whitefish (Coregonus clupeaformis). J. Therm. Biol. 2022, 104, 103185. [Google Scholar] [CrossRef] [PubMed]
- Kern, K.; Mertineit, C.L.; Brinkmann, R.; Miura, Y. Expression of heat shock protein 70 and cell death kinetics after different thermal impacts on cultured retinal pigment epithelial cells. Exp. Eye Res. 2018, 170, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Heydari, A.R.; Takahashi, R.; Gutsmann, A.; You, S.; Richardson, A. Hsp70 and aging. Experientia 1994, 50, 1092–1098. [Google Scholar] [CrossRef] [PubMed]
- Gabriel, J.E.; Ferro, J.A.; Stefani, R.M.; Ferro, M.I.; Gomes, S.L.; Macari, M. Effect of acute heat stress on heat shock protein 70 messenger RNA and on heat shock protein expression in the liver of broilers. Br. Poult. Sci. 1996, 37, 443–449. [Google Scholar] [CrossRef] [PubMed]
- Pike Winer, L.S.; Wu, M. Rapid Analysis of Glycolytic and Oxidative Substrate Flux of Cancer Cells in a Microplate. PLoS ONE 2014, 9, e109916. [Google Scholar] [CrossRef] [PubMed]
- Dohm, G.L.; Huston, R.L.; Askew, E.W.; Fleshood, H.L. Effects of exercise, training, and diet on muscle citric acid cycle enzyme activity. Can. J. Biochem. 1973, 51, 849–854. [Google Scholar] [CrossRef] [PubMed]
- McClelland, G.B.; Craig, P.M.; Dhekney, K.; Dipardo, S. Temperature- and exercise-induced gene expression and metabolic enzyme changes in skeletal muscle of adult zebrafish (Danio rerio). J. Physiol. 2006, 577, 739–751. [Google Scholar] [CrossRef] [PubMed]
- Peterson, R.H.; Anderson, J.M. Influence of Temperature Change on Spontaneous Locomotor Activity and Oxygen Consumption of Atlantic Salmon, Salmo salar, Acclimated to Two Temperatures. J. Fish. Res. Board Can. 1969, 26, 93–109. [Google Scholar] [CrossRef]
- Volkoff, H.; Rønnestad, I. Effects of temperature on feeding and digestive processes in fish. Temperature 2020, 7, 307–320. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Amplicon Size [bp] | Efficiency [E] | Sense (5′-3′) Antisense (5′-3′) | Source Acc. No. | Species |
|---|---|---|---|---|---|
| Hsf1 | 165 | 1.82 | GACTCTACACCTCAGACCTCACC | XM_014142743.1 | Salmo salar |
| AATTCAGAACCAGGCAGGGA | |||||
| Hsf2 | 331 | 1.85 | AGTTGATAGGCCACAGAGGTT | NM_001173904.1 | Salmo salar |
| CCAGACTTGCCTCCTTTGGA | |||||
| Hsp70 | 177 | 1.99 | CTCCTCTGGGTTGAAGGATTTG | Martorell-Ribera et al., 2020 [58] | Coregonus clupeaformis |
| GGCCATGAACCCCAACAACAC | |||||
| Hsp90 | 239 | 2.15 | AGATACGAGAGCTTGACAGACC | Martorell-Ribera et al., 2020 [58] | Coregonus clupeaformis |
| TGCCAGACTTGGCGATGGTTC | |||||
| Cs | 190 | 1.84 | CACCAACCTCCACCCTATGT | MH744411.1 | Coregonus clupeaformis |
| GCGGTAAAGGTTGCGGTAAA | |||||
| Eef1a1b | 189 | 2.09 | CCTCCACTTGGTCGTTTCG | Altmann et al., 2015 [54] | Coregonus clupeaformis |
| CGCAGGATGTAGGGCAGCAGA | |||||
| Rpl9 | 163 | 1.99 | ACCACATCAACCTGGAACTCA | Altmann et al., 2015 [54] | Coregonus clupeaformis |
| CGCATCTTGTAACGGAAACC | |||||
| Gapdh | 266 | 2.03 | CAAGGCTGTGGGCAAGGT | Altmann et al., 2015 [54] | Coregonus clupeaformis |
| GATGGAGGGAGTGGGTGTCT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tönißen, K.; Brenmoehl, J.; Wanka, H.; Grunow, B. Morphological and Metabolic Adaptations to Increasing Temperature: Insights from the In Vitro Model of Maraena Whitefish. Fishes 2025, 10, 352. https://doi.org/10.3390/fishes10070352
Tönißen K, Brenmoehl J, Wanka H, Grunow B. Morphological and Metabolic Adaptations to Increasing Temperature: Insights from the In Vitro Model of Maraena Whitefish. Fishes. 2025; 10(7):352. https://doi.org/10.3390/fishes10070352
Chicago/Turabian StyleTönißen, Katrin, Julia Brenmoehl, Heike Wanka, and Bianka Grunow. 2025. "Morphological and Metabolic Adaptations to Increasing Temperature: Insights from the In Vitro Model of Maraena Whitefish" Fishes 10, no. 7: 352. https://doi.org/10.3390/fishes10070352
APA StyleTönißen, K., Brenmoehl, J., Wanka, H., & Grunow, B. (2025). Morphological and Metabolic Adaptations to Increasing Temperature: Insights from the In Vitro Model of Maraena Whitefish. Fishes, 10(7), 352. https://doi.org/10.3390/fishes10070352