
Characterization and Analysis of the Role of Corazonin in Regulating Ovarian Development in the Mud Crab Scylla paramamosain



Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. RNA Isolation and Molecular Cloning
2.3. Expression Profiles of Sp-Crz and Sp-CrzR in the Female Mud Crab
2.4. Localization of Sp-Crz in the Eyestalk Ganglion by Immunohistochemistry
2.5. In Vitro Effect of Sp-Crz on Sp-Vg and Sp-VgR Expression
2.6. In Vivo Effect of Sp-Crz on the Ovarian Development of the Mud Crab
2.7. Determination of Estradiol (E2) Concentration in Hemolymph
2.8. Statistical Analysis
3. Results
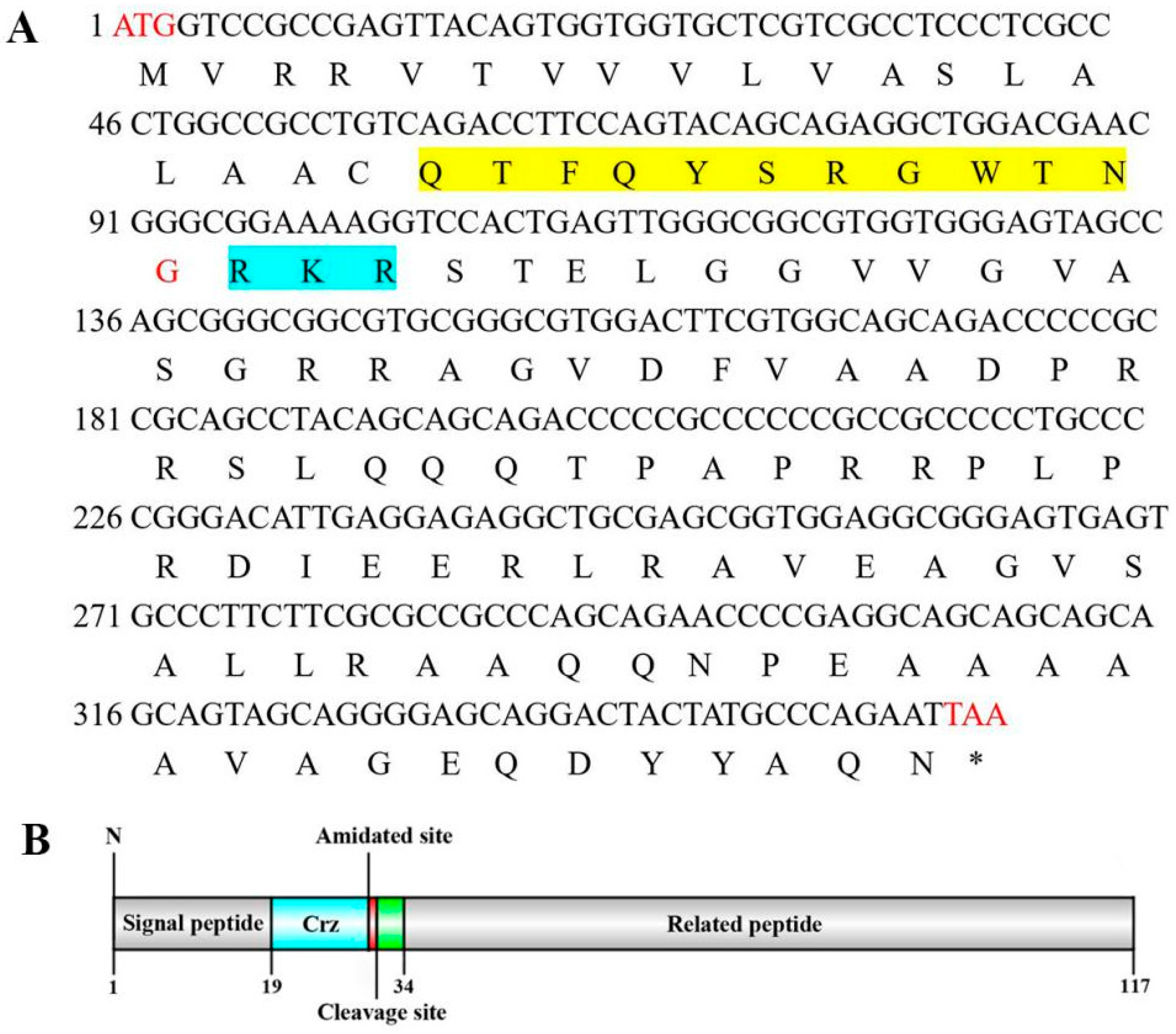
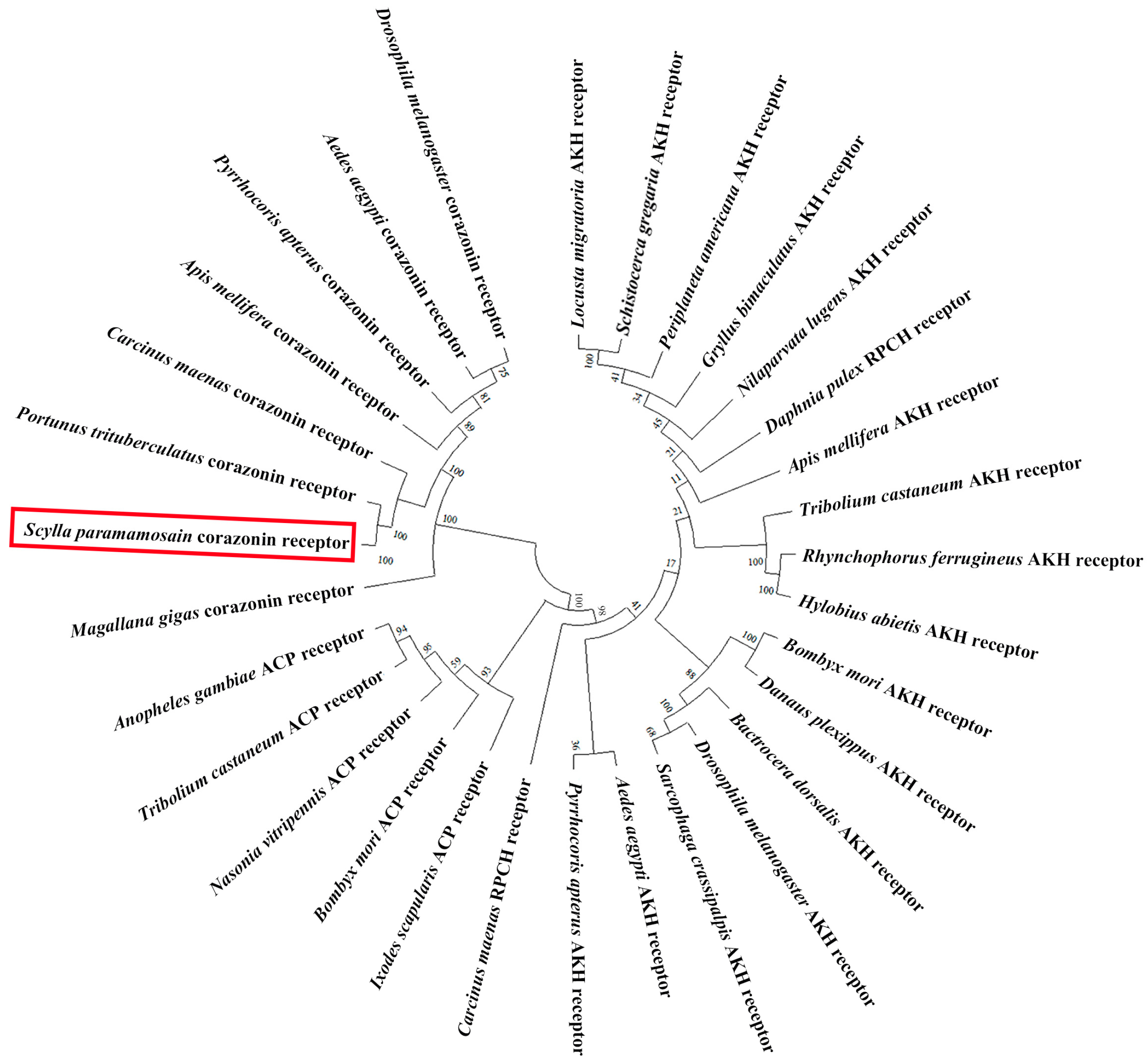
3.1. Cloning and Sequence Analysis of Sp-Crz and Its Receptor
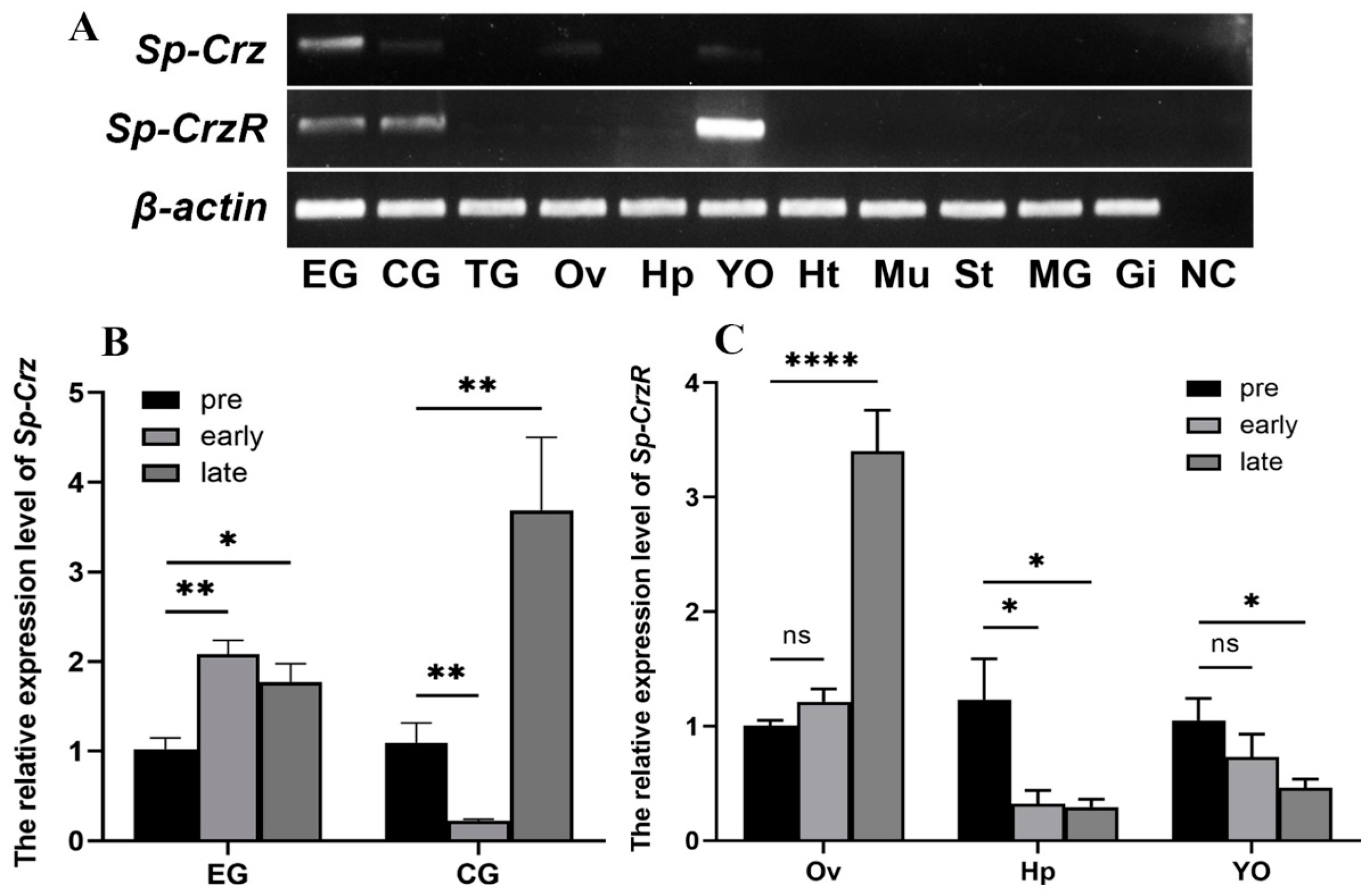
3.2. Expression Profiles of Sp-Crz and Sp-CrzR in Female S. paramamosain
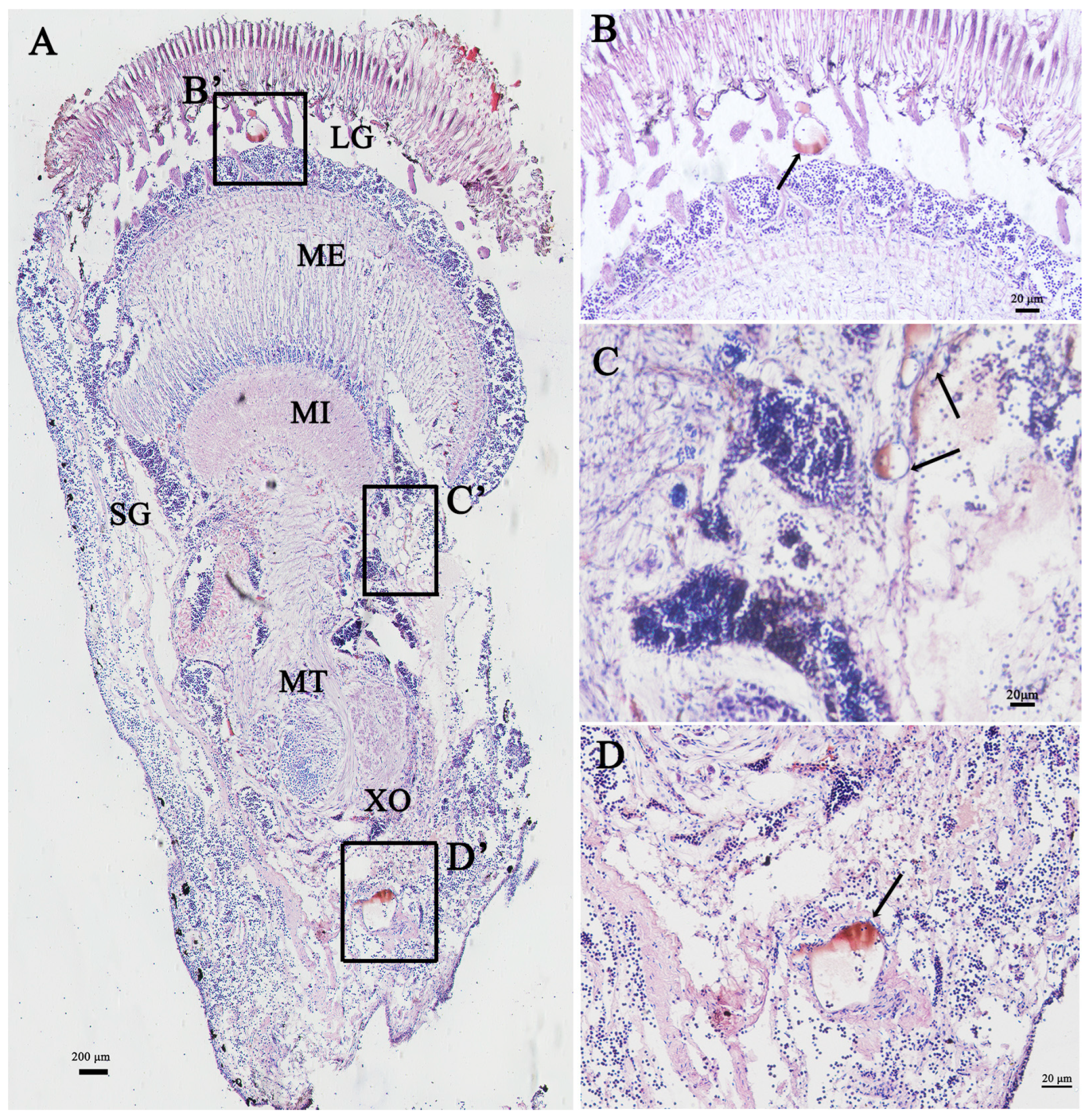
3.3. Localization of Sp-Crz in the Eyestalk Ganglion of S. paramamosain
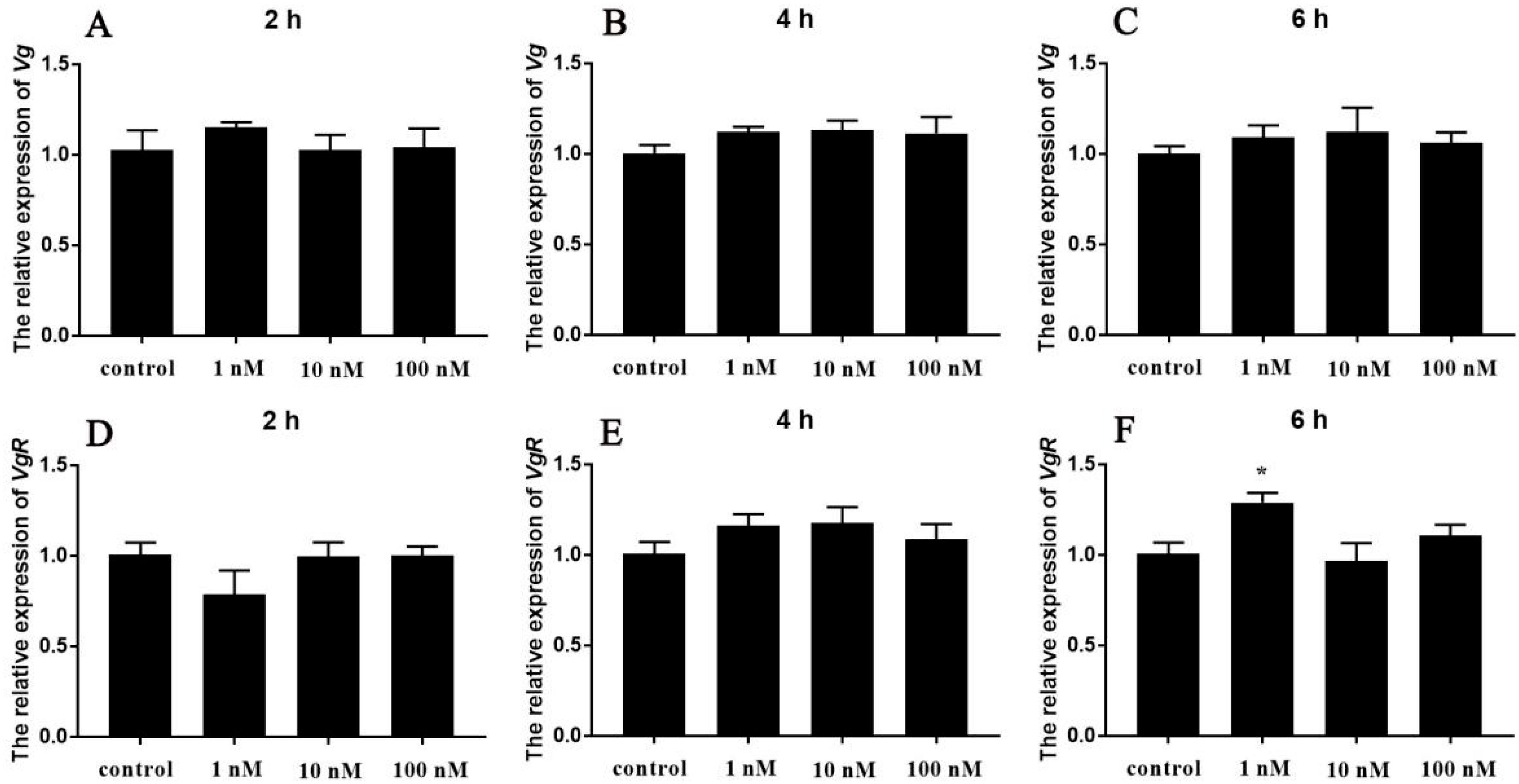
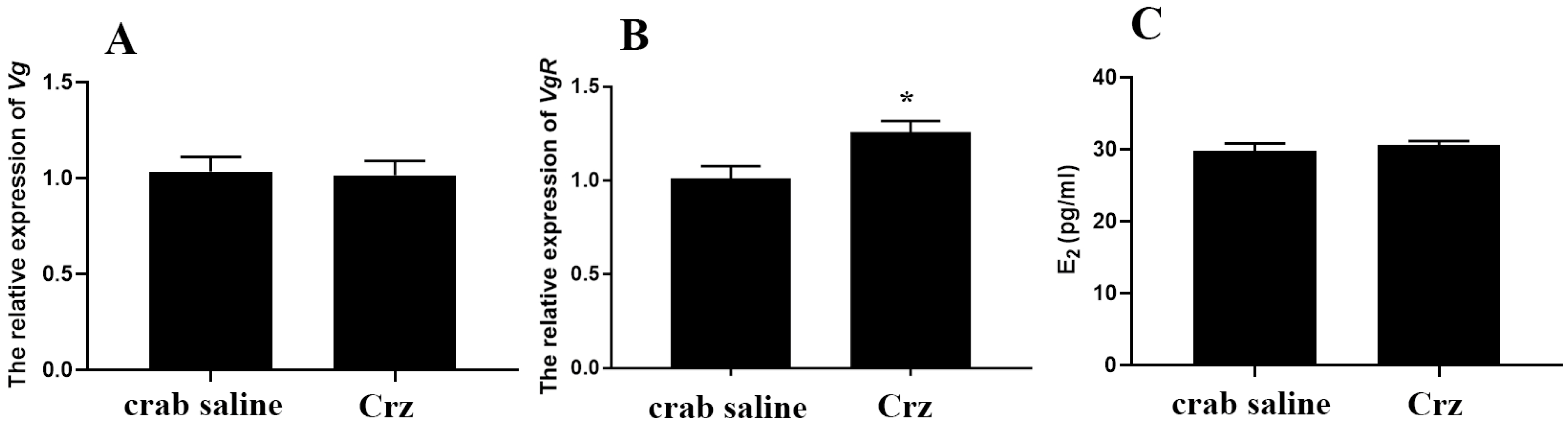
3.4. Effects of Sp-Crz on Vitellogenesis of S. paramamosain
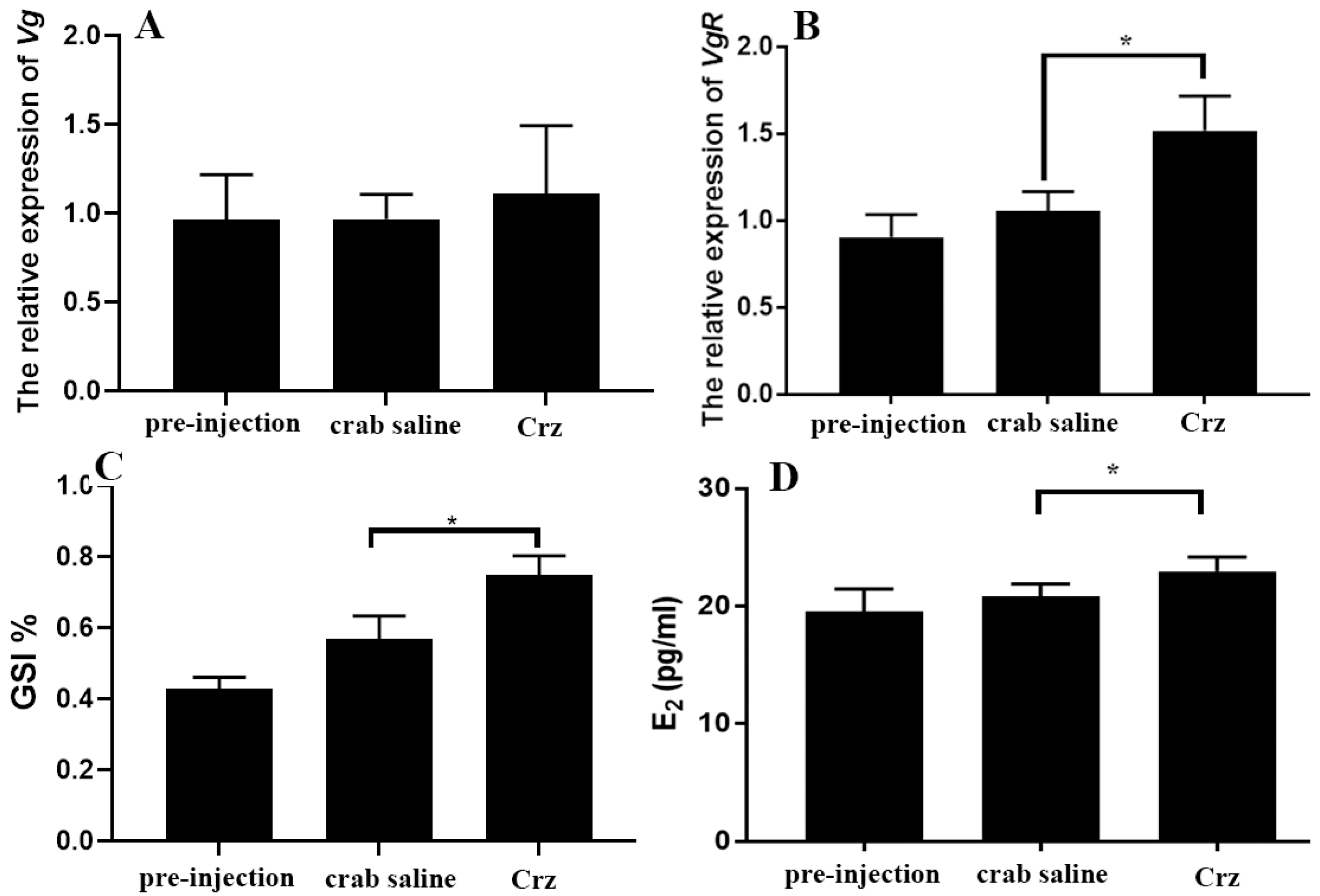
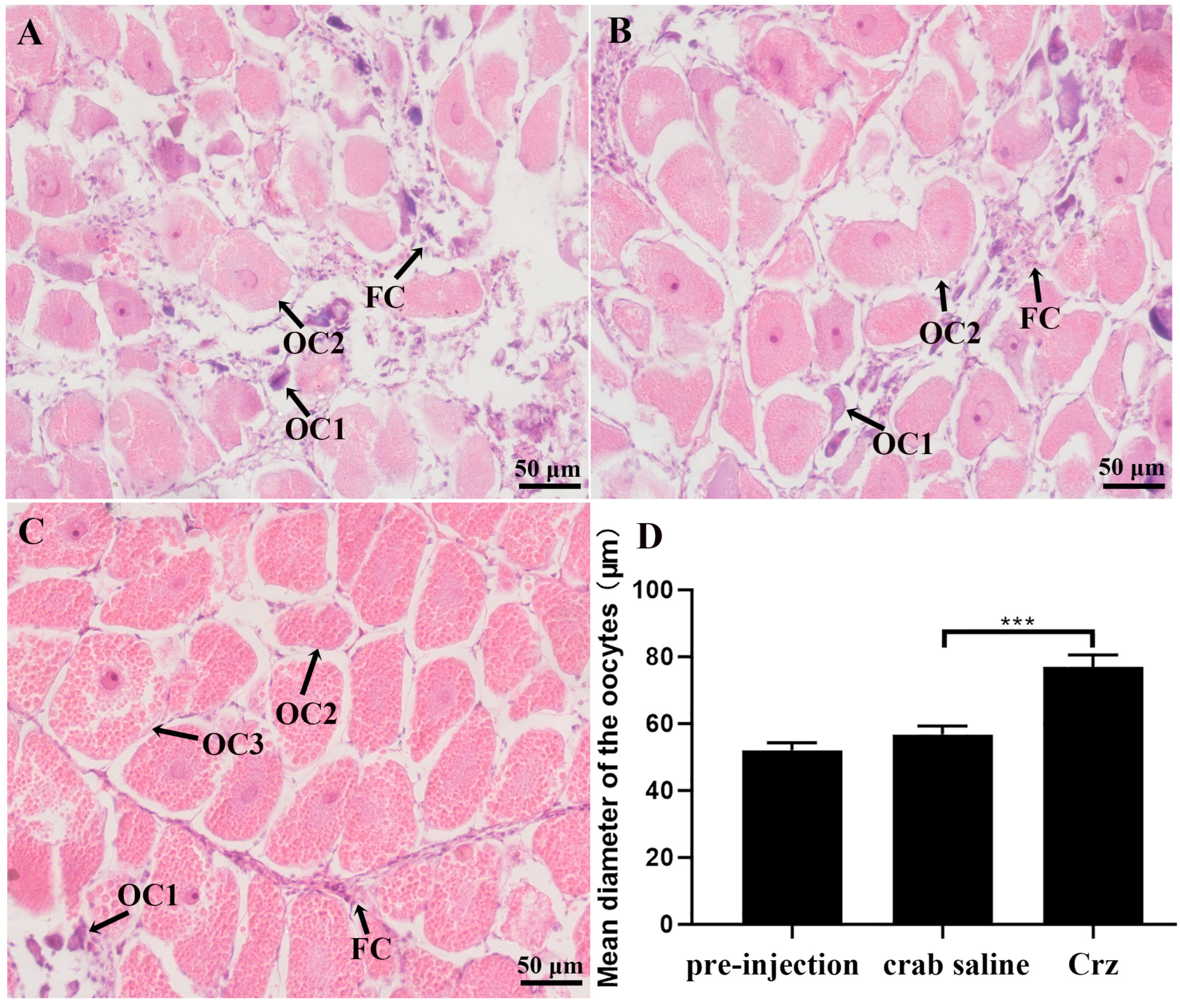
3.5. In Vivo Effects of Sp-Crz on S. paramamosain Ovarian Development
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Primer Sequence (5′-3′) | Application |
|---|---|---|
| Sp-Crz-OF | ATGGTGGCGGGGATCTCGTCTACCG | ORF validation |
| Sp-Crz-OR | TCACAGCTGTGTGTCCAACTCGCGG | ORF validation |
| Sp-Crz-F | AAAGTGTTCCGCAGTGAGC | RT-PCR |
| Sp-Crz-R | GTAGGGAGTCCAGCAGATGA | RT-PCR |
| Sp-CrzR-F | TCATCTGCTGGACTCCCTAC | RT-PCR |
| Sp-CrzR-R | TTCGTGCGACCTTCTGGT | RT-PCR |
| RV-M | GAGCGCATAACAATTTCACACA | RT-PCR |
| M13-47 | CGCCAGGGTTTTCCCAGTCACG | RT-PCR |
| Sp-Crz-qF | GTGAGTGCCCTTCTTCGC | qPCR |
| Sp-Crz-qR | GCAGTTTGGATGTTGTTTCG | qPCR |
| Sp-CrzR-qF | GGCAGCGTCCACCAGAA | qPCR |
| Sp-CrzR-qR | GCCCAAGCAAGCCAGAG | qPCR |
| Sp-Vg-qF | CGCAACCGCCACTGAAGAT | qPCR |
| Sp-Vg-qR | CCACCATGCTGCTCACGACT | qPCR |
| Sp-VgR-qF | TTCTATACCAGGCCACTACC | qPCR |
| Sp-VgR-qR | TTTTCACTCCAAGCACACTC | qPCR |
| β-actin-qF | GAGCGAGAAATCGTTCGTGAC | qPCR |
| β-actin-qR | GGAAGGAAGGCTGGAAGAGAG | qPCR |
References
- Hauser, F.; Cazzamali, G.; Williamson, M.; Blenau, W.; Grimmelikhuijzen, C.J.P. A review of neurohormone GPCRs present in the fruitfly Drosophila melanogaster and the honey bee. Apis mellifera. Prog. Neurobiol. 2006, 80, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Johnson, E.C. Post-genomic approaches to resolve neuropeptide signaling in Drosophila. In Invertebrate Neuropeptides and Hormones: Basic Knowledge and Recent Advances; Transworld Research Network: Trivandrum, India, 2006; pp. 179–224. [Google Scholar]
- Nässel, D.R. Neuropeptides in the nervous system of Drosophila and other insects: Multiple roles as neuromodulators and neurohormones. Prog. Neurobiol. 2002, 68, 1–84. [Google Scholar] [CrossRef] [PubMed]
- Taghert, P.H.; Veenstra, J.A. Drosophila neuropeptide signaling. Adv. Genet. 2003, 49, 1–65. [Google Scholar] [CrossRef] [PubMed]
- Veenstra, J.A. Isolation and structure of corazonin, a cardioactive peptide from the American cockroach. FEBS Lett. 1989, 250, 231–234. [Google Scholar] [CrossRef]
- Hua, Y.-J.; Ishibashi, J.; Saito, H.; Tawfik, A.I.; Sakakibara, M.; Tanaka, Y.; Derua, R.; Waelkens, E.; Baggerman, G.; De Loof, A.; et al. Identification of [Arg7] corazonin in the silkworm, Bombyx mori and the cricket, Gryllus bimaculatus, as a factor inducing dark color in an albino strain of the locust, Locusta migratoria. J. Insect Physiol. 2000, 46, 853–860. [Google Scholar] [CrossRef]
- Tanaka, S.; Pener, M.P. A neuropeptide controlling the dark pigmentation in color polymorphism of the migratory locust, Locusta migratoria. J. Insect Physiol. 1994, 40, 997–1005. [Google Scholar] [CrossRef]
- Kim, Y.J.; Spalovská-Valachová, I.; Cho, K.-H.; Zitnanova, I.; Park, Y.; Adams, M.E.; Žitňan, D. Corazonin receptor signaling in ecdysis initiation. Proc. Natl. Acad. Sci. USA 2004, 101, 6704–6709. [Google Scholar] [CrossRef]
- Belmont, M.; Cazzamali, G.; Williamson, M.; Hauser, F.; Grimmelikhuijzen, C.J.P. Identification of four evolutionarily related G protein-coupled receptors from the malaria mosquito Anopheles gambiae. Biochem. Biophys. Res. Commun. 2006, 344, 160–165. [Google Scholar] [CrossRef]
- Gospocic, J.; Shields, E.J.; Glastad, K.M.; Lin, Y.; Penick, C.A.; Yan, H.; Mikheyev, A.S.; Linksvayer, T.A.; Garcia, B.A.; Berger, S.L.; et al. The neuropeptide corazonin controls social behavior and caste identity in ants. Cell 2017, 170, 748–759.e12. [Google Scholar] [CrossRef]
- Tu, S.; Ge, F.; Han, Y.; Wang, M.; Xie, X.; Zhu, D. Putative role of corazonin in the ovarian development of the swimming crab Portunus trituberculatus. Front. Mar. Sci. 2022, 9, 976754. [Google Scholar] [CrossRef]
- Verleyen, P.; Baggerman, G.; Mertens, I.; Vandersmissen, T.; Huybrechts, J.; Lommel, A.V.; Loof, A.D.; Schoofs, L. Cloning and characterization of a third isoform of corazonin in the honey bee Apis mellifera. Peptides 2006, 27, 493–499. [Google Scholar] [CrossRef] [PubMed]
- Predel, R.; Neupert, S.; Russell, W.K.; Scheibner, O.; Nachman, R.J. Corazonin in insects. Peptides 2007, 28, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Hansen, K.K.; Stafflinger, E.; Schneider, M.; Hauser, F.; Cazzamali, G.; Williamson, M.; Kollmann, M.; Schachtner, J.; Grimmelikhuijzen, C.J.P. Discovery of a novel insect neuropeptide signaling system closely related to the insect adipokinetic hormone and corazonin hormonal systems. J. Biol. Chem. 2010, 285, 10736–10747. [Google Scholar] [CrossRef]
- Alexander, J.L.; Oliphant, A.; Wilcockson, D.C.; Audsley, N.; Down, R.E.; Lafont, R.; Webster, S.G. Functional characterization and signaling systems of corazonin and red pigment concentrating hormone in the green shore crab, Carcinus maenas. Front. Neurosci. 2018, 11, 752. [Google Scholar] [CrossRef]
- Minh Nhut, T.; Mykles, D.L.; Elizur, A.; Ventura, T. Ecdysis triggering hormone modulates molt behaviour in the redclaw crayfish Cherax quadricarinatus, providing a mechanistic evidence for conserved function in molt regulation across Pancrustacea. Gen. Comp. Endocrinol. 2020, 298, 113556. [Google Scholar] [CrossRef]
- Tawfik, A.I.; Tanaka, S.; De Loof, A.; Schoofs, L.; Baggerman, G.; Waelkens, E.; Derua, R.; Milner, Y.; Yerushalmi, Y.; Pener, M.P. Identification of the gregarization-associated dark-pigmentotropin in locusts through an albino mutant. Proc. Natl. Acad. Sci. USA 1999, 96, 7083–7087. [Google Scholar] [CrossRef]
- Tanaka, Y.; Hua, Y.J.; Roller, L.; Tanaka, S. Corazonin reduces the spinning rate in the silkworm, Bombyx mori. J. Insect Physiol. 2002, 48, 707–714. [Google Scholar] [CrossRef]
- Kapan, N.; Lushchak, O.V.; Luo, J.; Nässel, D.R. Identified peptidergic neurons in the Drosophila brain regulate insulin-producing cells, stress responses and metabolism by coexpressed short neuropeptide F and corazonin. Cell. Mol. Life Sci. 2012, 69, 4051–4066. [Google Scholar] [CrossRef]
- Tayler, T.D.; Pacheco, D.A.; Hergarden, A.C.; Murthy, M.; Anderson, D.J. A neuropeptide circuit that coordinates sperm transfer and copulation duration in Drosophila. Proc. Natl. Acad. Sci. USA 2012, 109, 20697–20702. [Google Scholar] [CrossRef]
- Sha, K. Functional Analysis of Corazonin and Its Receptor in Drosophila melanogaster. Ph.D. Thesis, The University of Tennessee, Knoxville, TN, USA, 2013. [Google Scholar]
- Oyeyinka, A.; Kansal, M.; O’Sullivan, S.M.; Gualtieri, C.; Smith, Z.M.; Vonhoff, F.J. Corazonin neurons contribute to dimorphic ethanol sedation sensitivity in Drosophila melanogaster. Front. Neural Circuits 2022, 16, 702901. [Google Scholar] [CrossRef]
- Lee, G.; Kim, K.M.; Kikuno, K.; Wang, Z.; Choi, Y.J.; Park, J.H. Developmental regulation and functions of the expression of the neuropeptide corazonin in Drosophila melanogaster. Cell Tissue Res. 2008, 331, 659–673. [Google Scholar] [CrossRef] [PubMed]
- Hou, Q.L.; Jiang, H.B.; Gui, S.H.; Chen, E.H.; Wei, D.D.; Li, H.M.; Wang, J.J.; Smagghe, G. A Role of corazonin receptor in larval-pupal transition and pupariation in the oriental fruit fly Bactrocera dorsalis (Hendel) (Diptera: Tephritidae). Front. Physiol. 2017, 8, 77. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Gu, G.X.; Teng, Z.W.; Wu, S.F.; Huang, J.; Song, Q.S.; Ye, G.Y.; Fang, Q. Identification and expression profiles of neuropeptides and their G protein-coupled receptors in the rice stem borer Chilo suppressalis. Sci. Rep. 2016, 6, 28976. [Google Scholar] [CrossRef] [PubMed]
- Fredriksson, R.; Lagerström, M.C.; Lundin, L.-G.; Schiöth, H.B. The G-protein-coupled receptors in the human genome form five main families. Phylogenetic analysis, paralogon groups, and fingerprints. Mol. Pharmacol. 2003, 63, 1256–1272. [Google Scholar] [CrossRef]
- Oldham, W.M.; Hamm, H.E. Heterotrimeric G protein activation by G-protein-coupled receptors. Nat. Rev. Mol. Cell Biol. 2008, 9, 60–71. [Google Scholar] [CrossRef]
- Cazzamali, G.; Saxild, N.P.E.; Grimmelikhuijzen, C.J.P. Molecular cloning and functional expression of a Drosophila corazonin receptor. Biochem. Biophys. Res. Commun. 2002, 298, 31–36. [Google Scholar] [CrossRef]
- Yang, J.; Huang, H.; Yang, H.; He, X.; Jiang, X.; Shi, Y.; Alatangaole, D.; Shi, L.; Zhou, N. Specific activation of the G protein-coupled receptor BNGR-A21 by the neuropeptide corazonin from the silkworm, Bombyx mori, Dually Couples to the Gq and Gs Signaling Cascades. J. Biol. Chem. 2013, 288, 11662–11675. [Google Scholar] [CrossRef]
- Sha, K.; Conner, W.C.; Choi, D.Y.; Park, J.H. Characterization, expression, and evolutionary aspects of corazonin neuropeptide and its receptor from the house fly, Musca domestica (Diptera: Muscidae). Gene 2012, 497, 191–199. [Google Scholar] [CrossRef]
- Hamoudi, Z.; Lange, A.B.; Orchard, I. Identification and characterization of the corazonin receptor and possible physiological roles of the corazonin-signaling pathway in Rhodnius prolixus. Front. Neurosci. 2016, 10. [Google Scholar] [CrossRef]
- Oryan, A.; Wahedi, A.; Paluzzi, J.-P.V. Functional characterization and quantitative expression analysis of two GnRH-related peptide receptors in the mosquito, Aedes aegypti. Biochem. Biophys. Res. Commun. 2018, 497, 550–557. [Google Scholar] [CrossRef]
- Buckley, S.J.; Fitzgibbon, Q.P.; Smith, G.G.; Ventura, T. In silico prediction of the G-protein coupled receptors expressed during the metamorphic molt of Sagmariasus verreauxi (Crustacea: Decapoda) by mining transcriptomic data: RNA-seq to repertoire. Gen. Comp. Endocrinol. 2016, 228, 111–127. [Google Scholar] [CrossRef] [PubMed]
- Tran, N.M.; Mykles, D.L.; Elizur, A.; Ventura, T. Characterization of G-protein coupled receptors from the blackback land crab Gecarcinus lateralis Y organ transcriptome over the molt cycle. BMC Genom. 2019, 20, 74. [Google Scholar] [CrossRef] [PubMed]
- Boerjan, B.; Verleyen, P.; Huybrechts, J.; Schoofs, L.; De Loof, A. In search for a common denominator for the diverse functions of arthropod corazonin: A role in the physiology of stress? Gen. Comp. Endocrinol. 2010, 166, 222–233. [Google Scholar] [CrossRef] [PubMed]
- Kubrak, O.I.; Lushchak, O.V.; Zandawala, M.; Nässel, D.R. Systemic corazonin signalling modulates stress responses and metabolism in Drosophila. Open Biol. 2016, 6, 160152. [Google Scholar] [CrossRef]
- Sha, K.; Choi, S.H.; Im, J.; Lee, G.G.; Loeffler, F.; Park, J.H. Regulation of ethanol-related behavior and ethanol metabolism by the corazonin neurons and corazonin receptor in Drosophila melanogaster. PLoS ONE 2014, 9, e87062. [Google Scholar] [CrossRef]
- Huang, X.; Ye, H.; Huang, H.; Yang, Y.; Gong, J. An insulin-like androgenic gland hormone gene in the mud crab, Scylla paramamosain, extensively expressed and involved in the processes of growth and female reproduction. Gen. Comp. Endocrinol. 2014, 204, 229–238. [Google Scholar] [CrossRef]
- Tan, W.; Tang, Y.; Liu, F.; Lu, L.; Liu, A.; Ye, H. Evaluation of the effect of adipokinetic hormone/corazonin-related peptide (ACP) on ovarian development in the mud crab, Scylla paramamosain. Animals 2024, 14, 3706. [Google Scholar] [CrossRef]
- Sláma, K.; Sakai, T.; Takeda, M. Effect of corazonin and crustacean cardioactive peptide on heartbeat in the adult American cockroach (Periplaneta americana). Arch. Insect Biochem. Physiol. 2006, 62, 91–103. [Google Scholar] [CrossRef]
- Roller, L.; Tanaka, Y.; Tanaka, S. Corazonin and corazonin-like substances in the central nervous system of the Pterygote and Apterygote insects. Cell Tissue Res. 2003, 312, 393–406. [Google Scholar] [CrossRef]
- Baggerman, G.; Cerstiaens, A.; De Loof, A.; Schoofs, L. Peptidomics of the larval Drosophila melanogaster central nervous system. J. Biol. Chem. 2002, 277, 40368–40374. [Google Scholar] [CrossRef]
- Chen, S.; Tang, Y.; Lu, L.; Gong, S.; Liu, A.; Ye, H. SIFa and its receptors play a possibly stimulatory role during ovarian development of the mud crab Scylla paramamosain. Front. Mar. Sci. 2024, 11, 1494264. [Google Scholar] [CrossRef]
- Ballesteros, J.A.; Shi, L.; Javitch, J.A. Structural mimicry in G protein-coupled receptors: Implications of the high-resolution structure of rhodopsin for structure-function analysis of rhodopsin-like receptors. Mol. Pharmacol. 2001, 60, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Zandawala, M.; Haddad, A.S.; Hamoudi, Z.; Orchard, I. Identification and characterization of the adipokinetic hormone/corazonin-related peptide signaling system in Rhodnius prolixus. FEBS J. 2015, 282, 3603–3617. [Google Scholar] [CrossRef]
- Simões, L.A.R.; Normann, R.S.; Chung, J.S.; Vinagre, A.S. A brief and updated introduction to the neuroendocrine system of crustaceans. Mol. Cell. Endocrinol. 2024, 590, 112265. [Google Scholar] [CrossRef]
- Xie, X.; Zhang, J.; Tu, S.; Zhou, Q.; Zhu, D. Corazonin stimulates ecdysteroid synthesis during the molting process of the swimming crab, Portunus trituberculatus. Biology 2024, 13, 630. [Google Scholar] [CrossRef]
- Zmora, N.; Trant, J.; Chan, S.M.; Chung, J.S. Vitellogenin and its messenger RNA during ovarian development in the female blue crab, callinectes sapidus: Gene expression, synthesis, transport, and Cleavage1. Biol. Reprod. 2007, 77, 138–146. [Google Scholar] [CrossRef]
- Bao, C.; Yang, Y.; Huang, H.; Ye, H. Neuropeptides in the cerebral ganglia of the mud crab, Scylla paramamosain: Transcriptomic analysis and expression profiles during vitellogenesis. Sci. Rep. 2015, 5, 17055. [Google Scholar] [CrossRef]
- Devaraj, H.; Natarajan, A. Molecular mechanisms regulating molting in a crustacean. FEBS J. 2006, 273, 839–846. [Google Scholar] [CrossRef]
- Jia, X.; Chen, Y.; Zou, Z.; Lin, P.; Wang, Y.; Zhang, Z. Characterization and expression profile of vitellogenin gene from Scylla paramamosain. Gene 2013, 520, 119–130. [Google Scholar] [CrossRef]
- Lu, L.; Wang, T.; Liu, A.; Ye, H. A single-cell atlas of crab ovary provides new insights into oogenesis in crustaceans. Adv. Sci. 2025, 12, 2409688. [Google Scholar] [CrossRef]
- Wang, T.; He, K.; Blaney, L.; Chung, J.S. 17β-Estradiol (E2) may be involved in the mode of crustacean female sex hormone (CFSH) action in the blue crab, Callinectes sapidus. Front. Endocrinol. 2022, 13, 962576. [Google Scholar] [CrossRef] [PubMed]
- Duangprom, S.; Saetan, J.; Phanaksri, T.; Songkoomkrong, S.; Surinlert, P.; Tamtin, M.; Sobhon, P.; Kornthong, N. Acceleration of ovarian maturation in the female mud crab with RNA interference of the vitellogenesis-inhibiting hormone (VIH). Front. Mar. Sci. 2022, 9, 880235. [Google Scholar] [CrossRef]
- Sumpownon, C.; Engsusophon, A.; Siangcham, T.; Sugiyama, E.; Soonklang, N.; Meeratana, P.; Wanichanon, C.; Hanna, P.J.; Setou, M.; Sobhon, P. Variation of prostaglandin E2 concentrations in ovaries and its effects on ovarian maturation and oocyte proliferation in the giant fresh water prawn, Macrobrachium rosenbergii. Gen. Comp. Endocrinol. 2015, 223, 129–138. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, S.; Liu, L.; Tang, Y.; Liu, A.; Ye, H. Characterization and Analysis of the Role of Corazonin in Regulating Ovarian Development in the Mud Crab Scylla paramamosain. Fishes 2025, 10, 329. https://doi.org/10.3390/fishes10070329
Yang S, Liu L, Tang Y, Liu A, Ye H. Characterization and Analysis of the Role of Corazonin in Regulating Ovarian Development in the Mud Crab Scylla paramamosain. Fishes. 2025; 10(7):329. https://doi.org/10.3390/fishes10070329
Chicago/Turabian StyleYang, Shiying, Liangjie Liu, Yiwei Tang, An Liu, and Haihui Ye. 2025. "Characterization and Analysis of the Role of Corazonin in Regulating Ovarian Development in the Mud Crab Scylla paramamosain" Fishes 10, no. 7: 329. https://doi.org/10.3390/fishes10070329
APA StyleYang, S., Liu, L., Tang, Y., Liu, A., & Ye, H. (2025). Characterization and Analysis of the Role of Corazonin in Regulating Ovarian Development in the Mud Crab Scylla paramamosain. Fishes, 10(7), 329. https://doi.org/10.3390/fishes10070329