
Effects of Co-Feeding Dry and Live Feed from the Onset of Exogenous Feeding on Red Seabream Pagrus major Larviculture and Pre-Growing


 ,
,
Abstract
1. Introduction
2. Materials and Methods
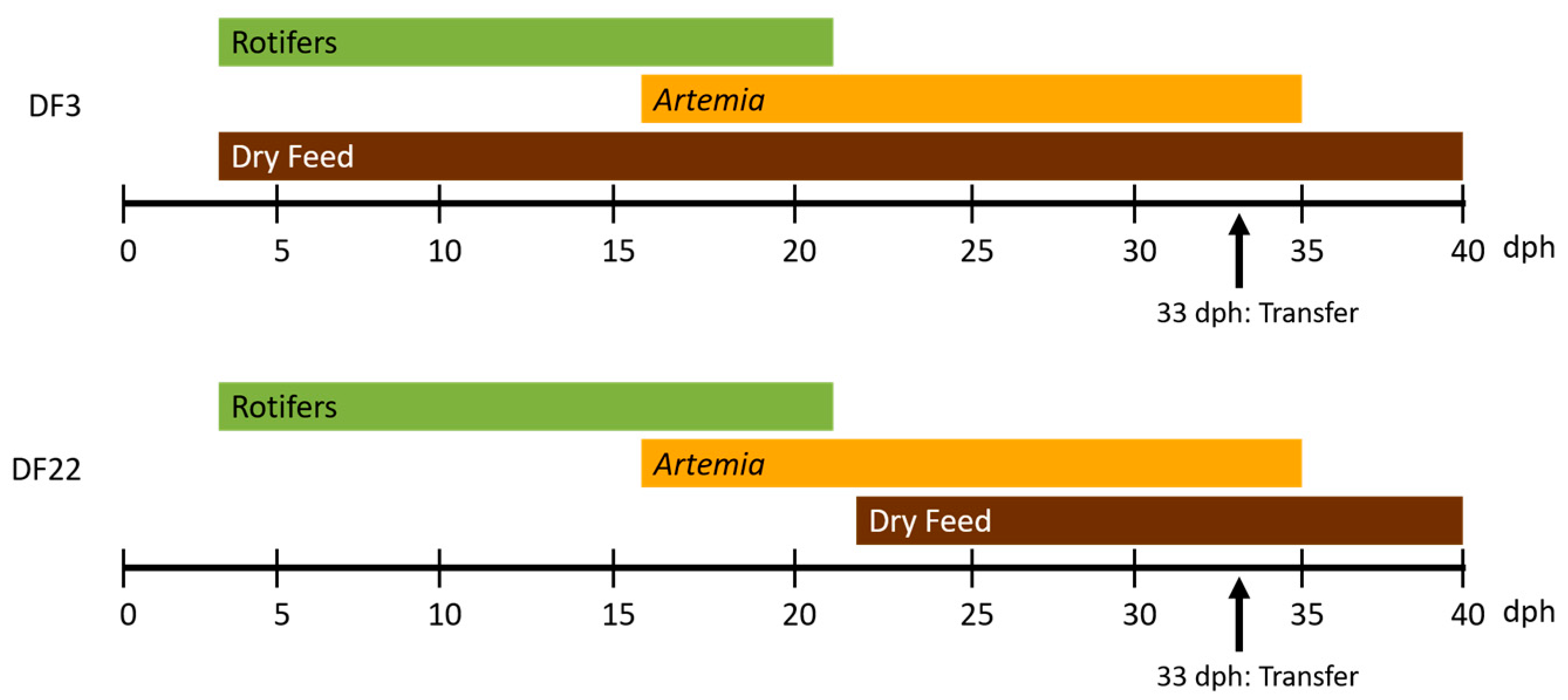
2.1. Experimental Design
2.1.1. Trial 1 (Hatchery Rearing, Hr)
2.1.2. First Grading at Hatchery
2.1.3. Trial 2 (Laboratory Rearing, Lr)
2.2. Analyses and Calculations
2.2.1. Trial 1 (Hatchery Rearing, Hr)
2.2.2. Information Obtained upon First Grading
- -
- Survival (%) = [(final number of survived larvae)/(initial number of stocked larvae)] × 100;
- -
- Big larvae (%) = [(number of Big larvae)/(number of survived larvae)] × 100;
- -
- Small larvae (%) = [(number of Small larvae)/(number of survived larvae)] × 100;
- -
- Mean weight of Big and Small larvae (group weighted).
2.2.3. Trial 2 (Laboratory Rearing, Lr)
2.3. Statistics
3. Results
3.1. Trial 1 (Hatchery Rearing, Hr)
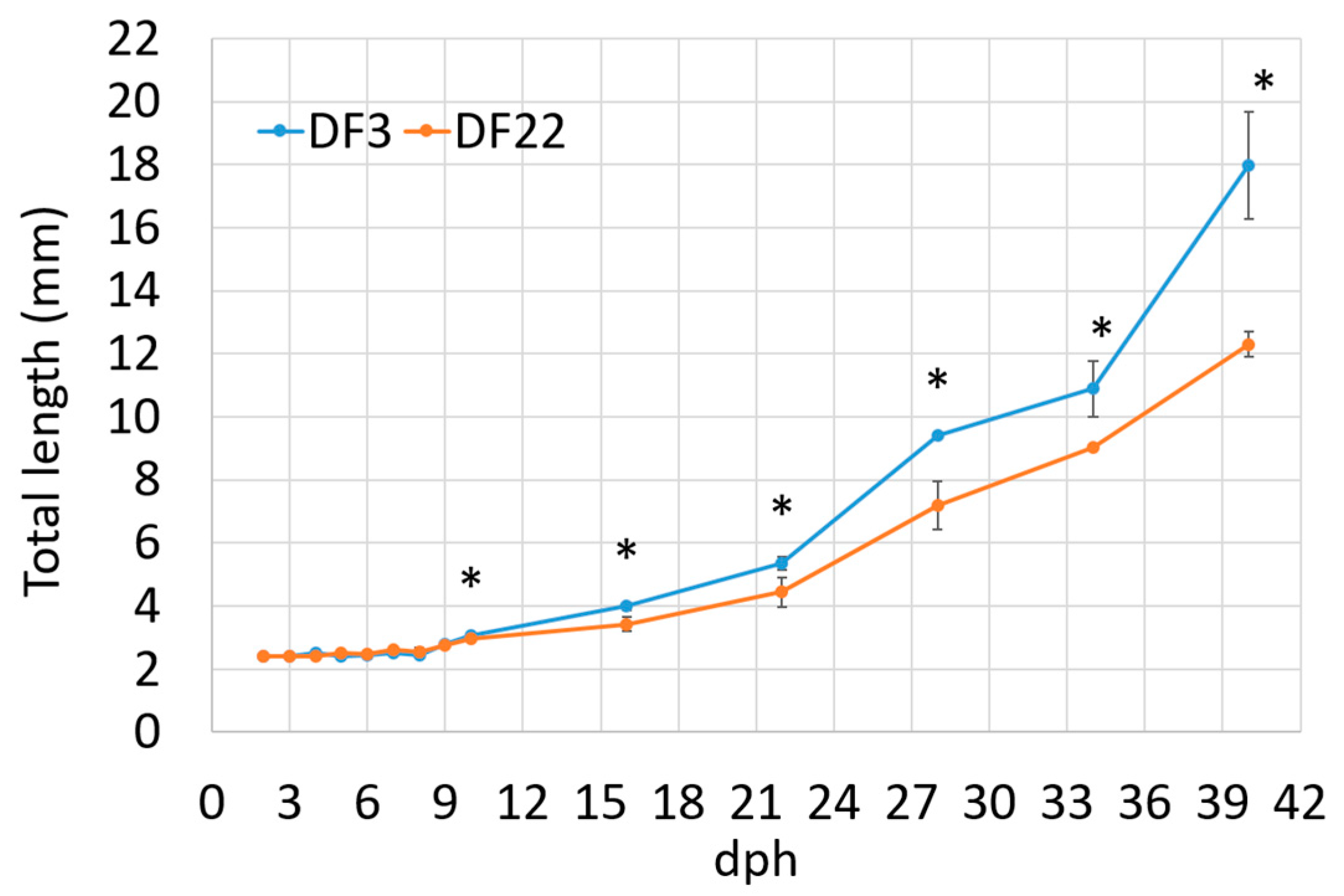
3.1.1. Larvae Growth
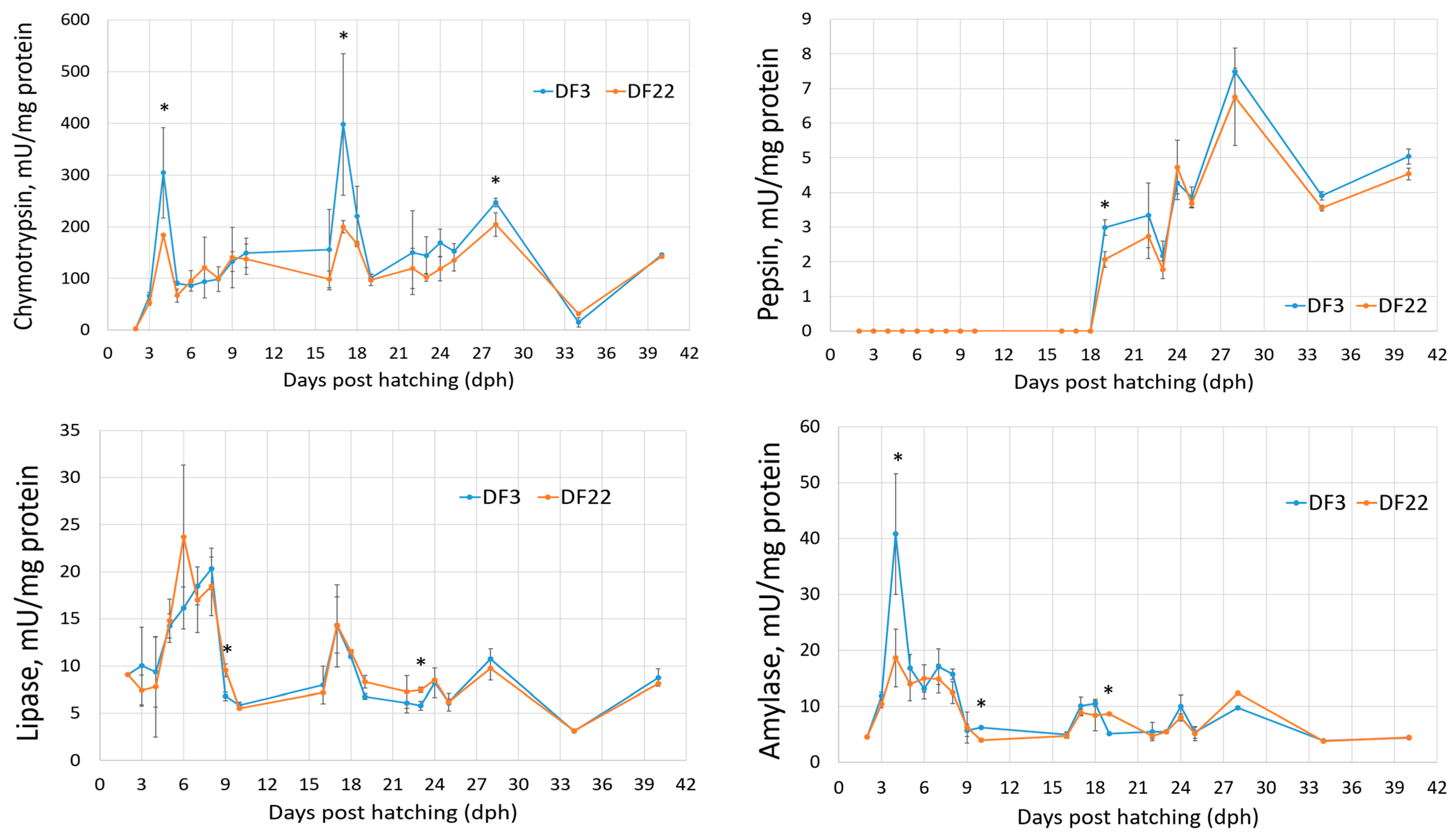
3.1.2. Digestive Enzymes
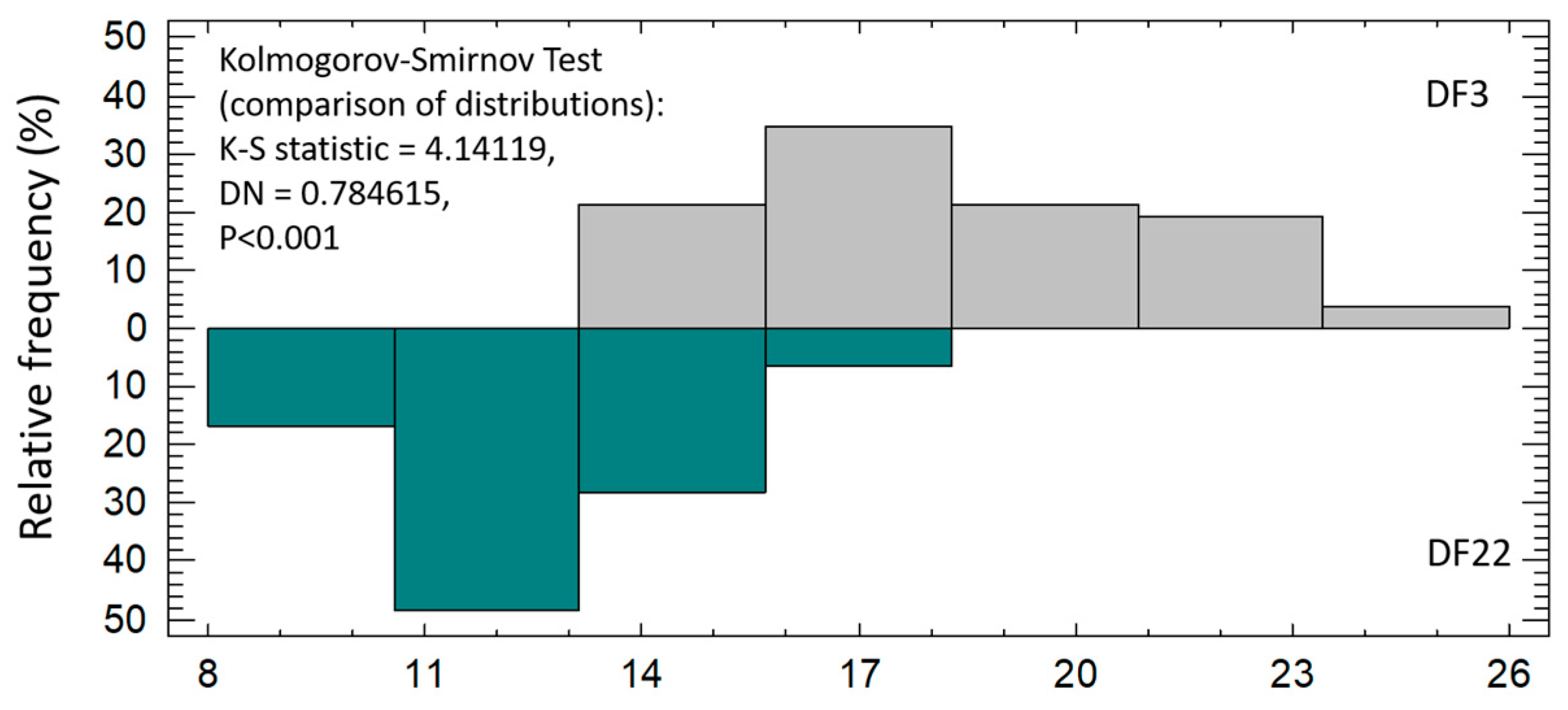
3.1.3. Hatchery Data Obtained on 1st Grading (53 dph)
3.2. Trial 2 (Laboratory Rearing, Lr)
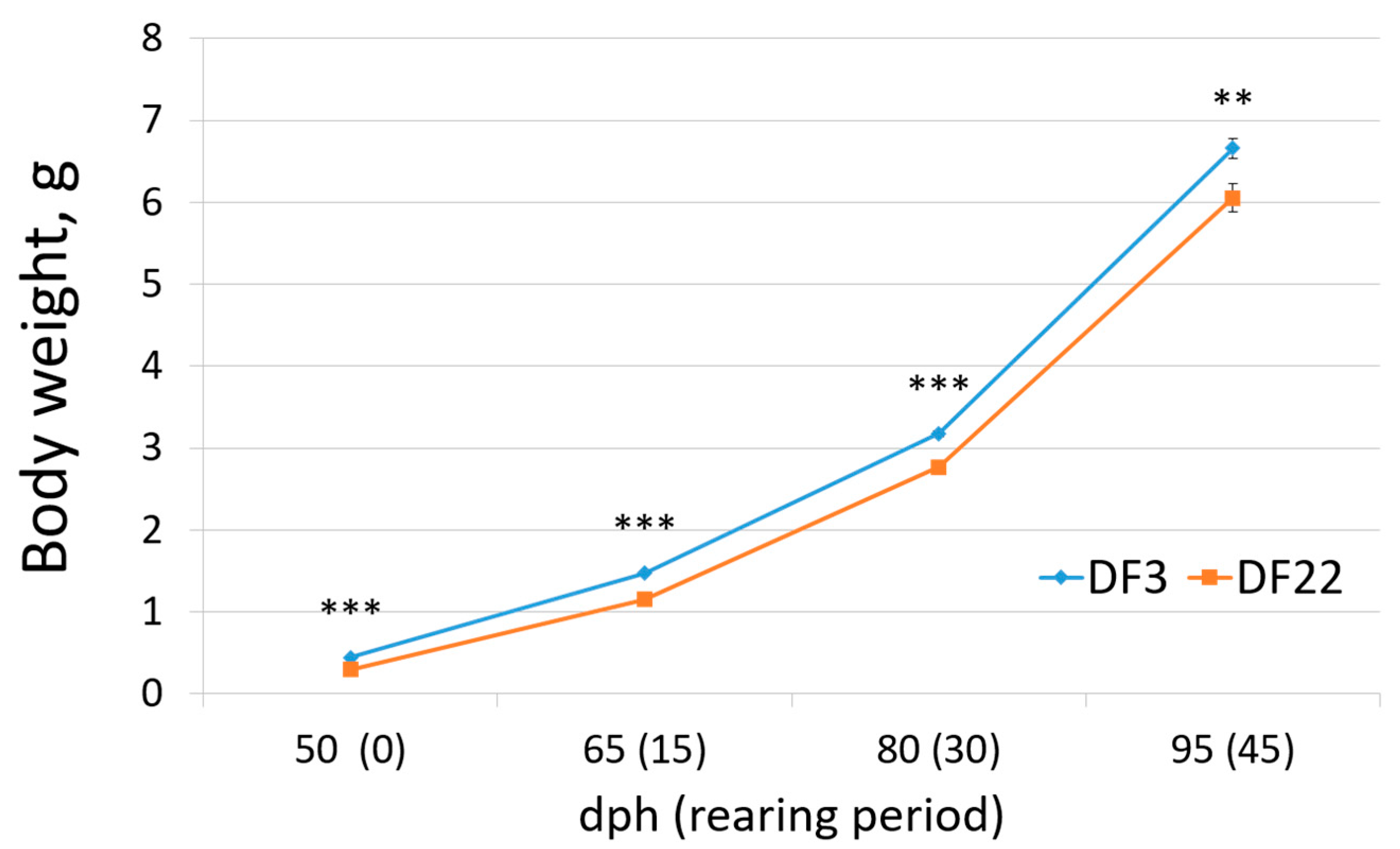
3.2.1. Growth Performance
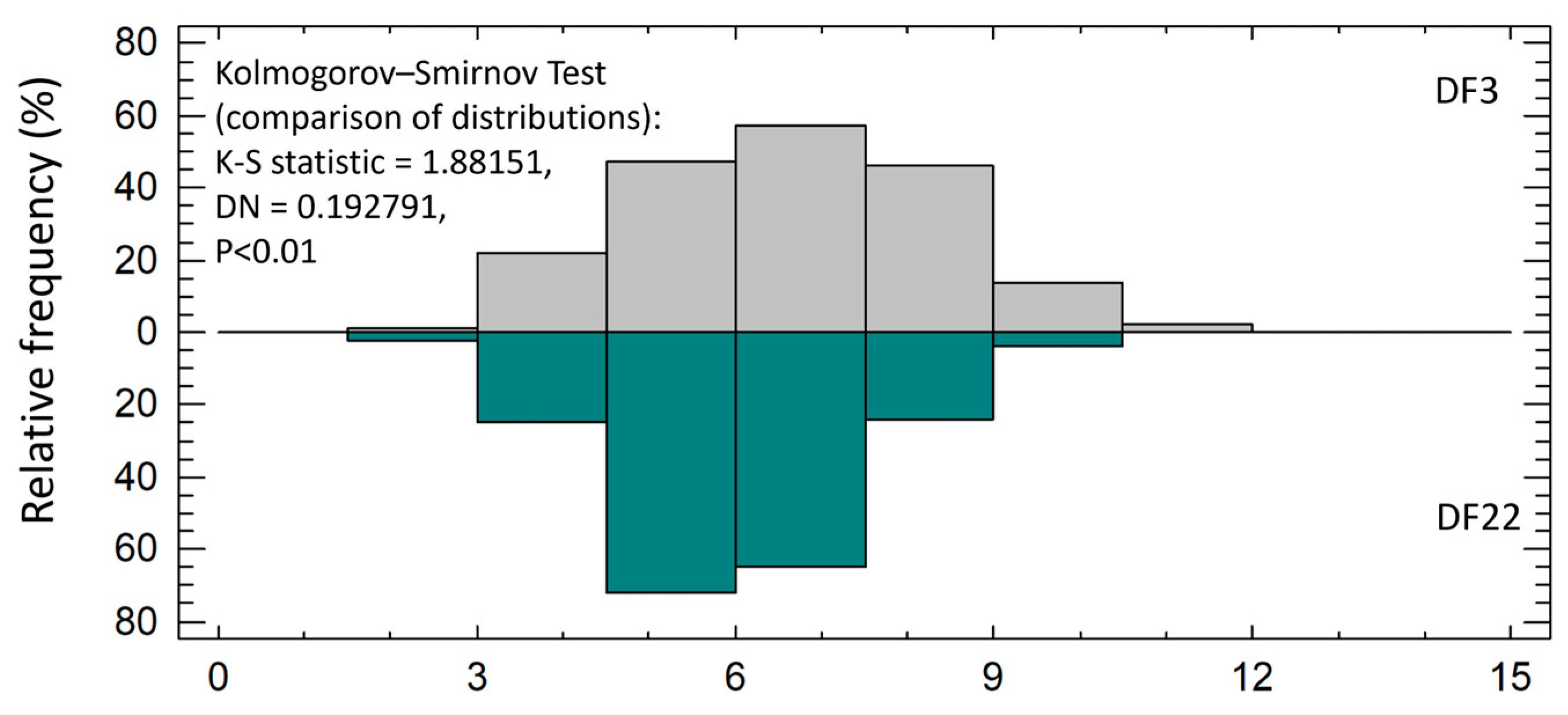
3.2.2. Phenotypical Deformities in Lr
4. Discussion
4.1. Larval Growth During Hr
4.2. Digestive Enzymes
4.3. Post-Larvae Growth During Lr
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Basurco, B.; Lovatelli, A.; Garcia, B. Current Status of Sparidae Aquaculture. In Sparidae: Biology and Aquaculture of Gilthead Sea Bream and Other Species; Pavlidis, M.A., Mylonas, C.C., Eds.; Wiley-Blackwell: Chichester, UK, 2011; pp. 1–50. ISBN 978-1-4051-9772-4. [Google Scholar]
- HAPO. Aquaculture in Greece: Aquaculture Annual Report 2024; Hellenic Aquaculture Producers Organisation: Athens, Greece, 2024. [Google Scholar]
- Rønnestad, I.; Yufera, M.; Ueberschar, B.; Ribeiro, L.; Sæle, Ø.; Boglione, C. Feeding Behaviour and Digestive Physiology in Larval Fish: Current Knowledge, and Gaps and Bottlenecks in Research. Rev. Aquac. 2013, 5, S59–S98. [Google Scholar] [CrossRef]
- Hagiwara, A.; Gallardo, W.G.; Assavaaree, M.; Kotani, T.; de Araujo, A.B. Live Food Production in Japan: Recent Progress and Future Aspects. Aquaculture 2001, 200, 111–127. [Google Scholar] [CrossRef]
- Yufera, M.; Conceicao, L.E.C.; Battaglene, S.; Fushimi, H.; Kotani, T. Early Development and Metabolism. In Sparidae: Biology and Aquaculture of Gilthead Sea Bream and Other Species; Pavlidis, M.A., Mylonas, C.C., Eds.; Wiley-Blackwell: Chichester, UK, 2011; pp. 133–155. ISBN 978-1-4051-9772-4. [Google Scholar]
- Hamre, K. Nutrient Profiles of Rotifers (Brachionus Sp.) and Rotifer Diets from Four Different Marine Fish Hatcheries. Aquaculture 2016, 450, 136–142. [Google Scholar] [CrossRef]
- Hart, P.; Purser, J. Weaning of Hatchery-Reared Greenback Flounder (Rhombosolea tapirina Gunther) from Live to Artificial Diets: Effects of Age and Duration of the Changeover Period. Aquaculture 1996, 145, 171–181. [Google Scholar] [CrossRef]
- Cahu, C.; Zambonino Infante, J. Substitution of Live Food by Formulated Diets in Marine Fish Larvae. Aquaculture 2001, 200, 161–180. [Google Scholar] [CrossRef]
- Mozes, N.; Papandroulakis, N.; Vergara, J.M.; Biswas, A.; Takii, K.; Ntatsopoulos, A. Production Systems. In Sparidae: Biology and Aquaculture of Gilthead Sea Bream and Other Species; Pavlidis, M.A., Mylonas, C.C., Eds.; Blackwell Publishing: Chichester, UK, 2011; pp. 169–198. ISBN 978-1-4051-9772-4. [Google Scholar]
- Hamre, K.; Yufera, M.; Rønnestad, I.; Boglione, C.; Conceicao, L.E.C.; Izquierdo, M. Fish Larval Nutrition and Feed Formulation: Knowledge Gaps and Bottlenecks for Advances in Larval Rearing. Rev. Aquac. 2013, 5, S26–S58. [Google Scholar] [CrossRef]
- Fernandez-Diaz, C.; Yufera, M. Detecting Growth in Gilthead Seabream, Sparus aurata L., Larvae Fed Microcapsules. Aquaculture 1997, 153, 93–102. [Google Scholar] [CrossRef]
- Fosse, P.J.; da Cruz Mattos, D.; Demier Cardoso, L.; Costa Radael, M.; Fosse Filho, J.; Vazquez Vidal, M.J. Duration of Co-Feeding on the Nishikigoi Cyprinus carpio Larvae during Weaning from Live to Inert Food in an Indoor System. Cienc. Rural 2018, 48, e20170579. [Google Scholar] [CrossRef]
- Khoa, T.N.D.; Waqalevu, V.; Honda, A.; Shiozaki, K.; Kotani, T. Comparative Study on Early Digestive Enzyme Activity and Expression in Red Sea Bream (Pagrus major) Fed on Live Feed and Micro-Diet. Aquaculture 2020, 519, 734721. [Google Scholar] [CrossRef]
- Chepkirui-Boit, V.; Ngugi, C.C.; Bowman, J.; Oyoo-Okoth, E.; Rasowo, J.; Mugo-Bundi, J.; Cherop, L. Growth Performance, Survival, Feed Utilization and Nutrient Utilization of African Catfish (Clarias gariepinus) Larvae Co-Fed Artemia and a Micro-Diet Containing Freshwater Atyid Shrimp (Caridina nilotica) during Weaning. Aquac. Nutr. 2011, 17, e82–e89. [Google Scholar] [CrossRef]
- Karakatsouli, N.; Batzina, A.; Ntomalis, K.; Panopoulos, S.; Coli, A.; Geropanagioti, E.; Anastasiadou, C.; Rati, M.; Bantounas, S. Co-Feeding Dry and Live Feed in First-Feeding Gilthead Seabream: Effects on Functional Development of the Digestive System, Larvae and Postlarvae Performance. Aquac. Nutr. 2021, 27, 2555–2566. [Google Scholar] [CrossRef]
- Kestemont, P.; Xueliang, X.; Hamza, N.; Maboudou, J.; Imorou Toko, I. Effect of Weaning Age and Diet on Pikeperch Larviculture. Aquaculture 2007, 264, 197–204. [Google Scholar] [CrossRef]
- Canavate, J.P.; Fernandez-Diaz, C. Influence of Co-Feeding Larvae with Live and Inert Diets on Weaning the Sole Solea senegalensis onto Commercial Dry Feeds. Aquaculture 1999, 174, 255–263. [Google Scholar] [CrossRef]
- Mai, M.G.; Engrola, S.; Morais, S.; Portella, M.C.; Verani, J.R.; Dinis, M.T.; Conceicao, L.E.C. Co-Feeding of Live Feed and Inert Diet from First-Feeding Affects Artemia Lipid Digestibility and Retention in Senegalese Sole (Solea senegalensis) Larvae. Aquaculture 2009, 296, 284–291. [Google Scholar] [CrossRef]
- Engrola, S.; Figueira, L.; Conceicao, L.E.C.; Gavaia, P.J.; Ribeiro, L.; Dinis, M.T. Co-Feeding in Senegalese Sole Larvae with Inert Diet from Mouth Opening Promotes Growth at Weaning. Aquaculture 2009, 288, 264–272. [Google Scholar] [CrossRef]
- Engrola, S.; Mai, M.; Dinis, M.T.; Conceicao, L.E.C. Co-Feeding of Inert Diet from Mouth Opening Does Not Impair Protein Utilization by Senegalese Sole (Solea senegalensis) Larvae. Aquaculture 2009, 287, 185–190. [Google Scholar] [CrossRef]
- Engrola, S.; Dinis, M.T.; Conceicao, L.E.C. Senegalese Sole Larvae Growth and Protein Utilization Is Depressed When Co-Fed High Levels of Inert Diet and Artemia since First Feeding. Aquac. Nutr. 2010, 16, 457–465. [Google Scholar] [CrossRef]
- Wang, Y.; Hu, M.; Cao, L.; Wang, W. Effects of Daphnia (Moina micrura) plus Chlorella (Chlorella pyrenoidosa) or Microparticle Diets on Growth and Survival of Larval Loach (Misgurnus anguillicaudatus). Aquac. Int. 2008, 16, 361–368. [Google Scholar] [CrossRef]
- Wang, Y.; Hu, M.; Wang, W.; Cao, L. Effects on Growth and Survival of Loach (Misgurnus anguillicaudatus) Larvae When Co-Fed on Live and Microparticle Diets. Aquac. Res. 2009, 40, 385–394. [Google Scholar] [CrossRef]
- Giri, S.S.; Sahoo, S.K.; Sahu, B.B.; Sahu, A.K.; Mohanty, S.N.; Mukhopadhyay, P.K.; Ayyappan, S. Larval Survival and Growth in Wallago attu (Bloch and Schneider): Effects of Light, Photoperiod and Feeding Regimes. Aquaculture 2002, 213, 151–161. [Google Scholar] [CrossRef]
- Marinho, Y.F.; Oliveira, C.Y.B.; Mendes, L.E.M.; Santos, I.R.A.; Dias, J.A.R.; Ândrade, M.; Lopes, Y.V.A.; Azevedo, J.W.J.; Lourenço, C.B.; Moura, R.S.T.; et al. Co-Feeding Using Live Food and Feed as First Feeding for the Small Catfish Trachelyopterus galeatus (Linnaeus 1766). Arq. Bras. Med. Veterinária E Zootec. 2024, 76, 323–332. [Google Scholar] [CrossRef]
- Sweetman, J.W. Larviculture of Mediterranean Marine Fish Species: Current Status and Future Trends. J. World Aquac. Soc. 1992, 23, 330–337. [Google Scholar] [CrossRef]
- López-Alvarado, J.; Kanazawa, A. Effect of Dietary Protein Sources in Microdiets on Feeding Behavior and Growth of Red Sea Bream, Pagrus major, During Weaning and Metamorphosis. J. Appl. Aquac. 1997, 7, 53–66. [Google Scholar] [CrossRef]
- El-Sayed, H.S.; Ghonim, A.Z.; El-Khodary, G.M.; El-Sheikh, M.A.; Khairy, H.M. Application of Enriched Cyclops Abyssorum divergens with Mixed Algal Diet Compared to Artemia franciscana for Improving Larval Growth and Body Composition of Dicentrarchus labrax. Aquac. Rep. 2021, 20, 100715. [Google Scholar] [CrossRef]
- Lainé, J.; Beattie, M.; LeBel, D. Simultaneous Kinetic Determinations of Lipase, Chymotrypsin, Trypsin, Elastase, and Amylase on the Same Microtiter Plate. Pancreas 1993, 8, 383–386. [Google Scholar] [CrossRef]
- Anson, M.L. The Estimation of Pepsin, Trypsin, Papain, and Cathepsin with Hemoglobin. J. Gen. Physiol. 1938, 22, 79–89. [Google Scholar] [CrossRef]
- Somogyi, M. Modifications of Two Methods for the Assay of Amylase. Clin. Chem. 1960, 6, 23–35. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.-J.; Hwang, J.M.; Kim, S.I. A Colorimetric Microplate Assay Method for High Throughput Analysis of Lipase Activity. J. Biochem. Mol. Biol. 2003, 36, 417–420. [Google Scholar] [CrossRef]
- Bradford, M.M. Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Khoa, T.N.D.; Waqalevu, V.; Honda, A.; Shiozaki, K.; Kotani, T. Early Ontogenetic Development, Digestive Enzymatic Activity and Gene Expression in Red Sea Bream (Pagrus major). Aquaculture 2019, 512, 734283. [Google Scholar] [CrossRef]
- Sasaki, T.; Matsui, H.; Kuwabara, Y.; Yokoyama, S.; Ishikawa, M.; Kotani, T. Euryhaline Copepod Pseudodiaptomus inopinus Changed the Prey Preference of Red Sea Bream Pagrus major Larvae. Fish. Sci. 2024, 90, 281–294. [Google Scholar] [CrossRef]
- Andrade, C.A.P.; Nascimento, F.; Conceicao, L.E.C.; Linares, F.; Lacuisse, M.; Dinis, M.T. Red Porgy, Pagrus pagrus, Larvae Performance and Nutritional Condition in Response to Different Weaning Regimes. J. World Aquac. Soc. 2012, 43, 321–334. [Google Scholar] [CrossRef]
- Andrade, C.A.P.; Nascimento, F.; Noguiera, N.; Pimenta, F.; Dinis, M.T.; Narciso, L. Allometric Growth in Red Porgy Larvae: Developing Morphological Indices for Mesocosm Semi-Intensive Culture. N. Am. J. Aquac. 2013, 75, 42–49. [Google Scholar] [CrossRef]
- Andrade, C.A.P.; Soares, F.; Ribeiro, L.; Roo, F.; Socorro, J.; Dinis, M.T. Morphological, Histological, Histochemical and Behavioral Aspects during Early Development of Red Porgy Pagrus pagrus L. Reared in Mesocosm. Turk. J. Fish. Aquat. Sci. 2015, 15, 137–148. [Google Scholar] [CrossRef]
- Mihelakakis, A.; Yoshimatsu, T.; Tsolkas, C. Spawning in Captivity and Early Life History of Cultured Red Porgy, Pagrus pagrus. Aquaculture 2001, 199, 333–352. [Google Scholar] [CrossRef]
- Papandroulakis, N.; Kentouri, M.; Divanach, P. Biological Performance of Red Porgy (Pagrus pagrus) Larvae under Intensive Rearing Conditions with the Use of an Automated Feeding System. Aquac. Int. 2004, 12, 191–203. [Google Scholar] [CrossRef]
- Darias, M.J.; Ortiz-Delgado, J.B.; Sarasquete, C.; Martinez-Rodriguez, G.; Yufera, M. Larval Organogenesis of Pagrus pagrus L., 1758 with Special Attention to the Digestive System Development. Histol. Histopathol. 2007, 22, 753–768. [Google Scholar] [CrossRef]
- Suzer, C.; Kamaci, H.O.; Coban, D.; Saka, S.; Firat, K.; Ozkara, B.; Ozkara, A. Digestive Enzyme Activity of the Red Porgy (Pagrus pagrus, L.) during Larval Development under Culture Conditions. Aquac. Res. 2007, 38, 1778–1785. [Google Scholar] [CrossRef]
- Uematsu, K.; Kitano, M.; Morita, M.; Iijima, N. Presence and Ontogeny of Intestinal and Pancreatic Phospholipase A2-like Proteins in the Red Sea Bream, Pagrus major. An Immunocytochemical Study. Fish Physiol. Biochem. 1992, 9, 427–438. [Google Scholar] [CrossRef]
- Roo, F.J.; Socorro, J.; Izquierdo, M.S.; Caballero, M.J.; Hernández-Cruz, C.M.; Fernández, A.; Fernández-Palacios, H. Development of Red Porgy Pagrus pagrus Visual System in Relation with Changes in the Digestive Tract and Larval Feeding Habits. Aquaculture 1999, 179, 499–512. [Google Scholar] [CrossRef]
- Darias, M.J.; Murray, H.M.; Gallant, J.W.; Douglas, S.E.; Yúfera, M.; Martínez-Rodríguez, G. The Spatiotemporal Expression Pattern of Trypsinogen and Bile Salt-Activated Lipase during the Larval Development of Red Porgy (Pagrus pagrus, Pisces, Sparidae). Mar. Biol. 2007, 152, 109–118. [Google Scholar] [CrossRef]
- Darias, M.J.; Murray, H.M.; Gallant, J.W.; Douglas, S.E.; Yúfera, M.; Martínez-Rodríguez, G. Ontogeny of Pepsinogen and Gastric Proton Pump Expression in Red Porgy (Pagrus pagrus): Determination of Stomach Functionality. Aquaculture 2007, 270, 369–378. [Google Scholar] [CrossRef]
- Sánchez-Amaya, M.I.; Ortiz-Delgado, J.B.; García-López, Á.; Cárdenas, S.; Sarasquete, C. Larval Ontogeny of Redbanded Seabream Pagrus auriga Valenciennes, 1843 with Special Reference to the Digestive System. A Histological and Histochemical Approach. Aquaculture 2007, 263, 259–279. [Google Scholar] [CrossRef]
- Cahu, C.; Rønnestad, I.; Grangier, V.; Zambonino Infante, J.L. Expression and Activities of Pancreatic Enzymes in Developing Sea Bass Larvae (Dicentrarchus labrax) in Relation to Intact and Hydrolyzed Dietary Protein; Involvement of Cholecystokinin. Aquaculture 2004, 238, 295–308. [Google Scholar] [CrossRef]
- Waqalevu, V.; Honda, A.; Dossou, S.; Khoa, T.N.D.; Matsui, H.; Mzengereza, K.; Liu, H.; Ishikawa, M.; Shiozaki, K.; Kotani, T. Effect of Oil Enrichment on Brachionus plicatilis Rotifer and First Feeding Red Sea Bream (Pagrus major) and Japanese Flounder (Paralichthys olivaceus). Aquaculture 2019, 510, 73–83. [Google Scholar] [CrossRef]
- Sargent, J.; McEvoy, L.; Estevez, A.; Bell, G.; Bell, M.; Henderson, J.; Tocher, D. Lipid Nutrition of Marine Fish during Early Development: Current Status and Future Directions. Aquaculture 1999, 179, 217–229. [Google Scholar] [CrossRef]
- Tocher, D.R.; Bendiksen, E.Å.; Campbell, P.J.; Bell, J.G. The Role of Phospholipids in Nutrition and Metabolism of Teleost Fish. Aquaculture 2008, 280, 21–34. [Google Scholar] [CrossRef]
- Foscarini, R. A Review: Intensive Farming Procedure for Red Sea Bream (Pagrus major) in Japan. Aquaculture 1988, 72, 191–246. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DF3 | DF22 | |||||
|---|---|---|---|---|---|---|
| dph | Rotifers 1, mil/m3/day | Artemia 2, mil/m3/day | Dry Feed 3, g/m3/day | Rotifers, mil/m3/day | Artemia, mil/m3/day | Dry Feed, g/m3/day |
| 1 | ||||||
| 2 | ||||||
| 3 | 6.0 | 6.7 | 6.0 | |||
| 4 | 46.0 | 8.3 | 46.0 | |||
| 5 | 14.5 | 8.3 | 14.5 | |||
| 6 | 16.0 | 8.3 | 16.0 | |||
| 7 | 15.0 | 9.9 | 15.0 | |||
| 8 | 26.0 | 10.0 | 26.0 | |||
| 9 | 32.0 | 23.2 | 32.0 | |||
| 10 | 34.0 | 16.5 | 34.0 | |||
| 11 | 34.0 | 16.6 | 34.0 | |||
| 12 | 32.0 | 16.7 | 32.0 | |||
| 13 | 30.0 | 20.0 | 30.0 | |||
| 14 | 42.0 | 20.0 | 42.0 | |||
| 15 | 30.0 | 25.0 | 30.0 | |||
| 16 | 38.0 | 0.4 | 25.0 | 38.0 | 0.4 | |
| 17 | 26.0 | 0.8 | 25.0 | 26.0 | 0.8 | |
| 18 | 30.0 | 1.6 | 25.0 | 30.0 | 1.6 | |
| 19 | 30.0 | 3.0 | 25.0 | 30.0 | 3.0 | |
| 20 | 32.0 | 3.8 | 25.0 | 32.0 | 3.8 | |
| 21 | 32.0 | 4.3 | 25.0 | 32.0 | 4.3 | |
| 22 | 3.8 | 25.0 | 3.8 | 25.0 | ||
| 23 | 4.1 | 25.0 | 4.1 | 25.0 | ||
| 24 | 4.9 | 25.0 | 4.9 | 25.0 | ||
| 25 | 6.0 | 25.0 | 6.0 | 25.0 | ||
| 26 | 4.8 | 25.0 | 4.8 | 25.0 | ||
| 27 | 7.2 | 25.0 | 7.2 | 25.0 | ||
| 28 | 7.4 | 25.0 | 7.4 | 25.0 | ||
| 29 | 7.7 | 25.0 | 7.7 | 25.0 | ||
| 30 | 7.4 | 25.0 | 7.4 | 25.0 | ||
| 31 | 7.0 | 33.3 | 7.0 | 33.3 | ||
| 32 | 7.0 | 33.3 | 7.0 | 33.3 | ||
| 33 | 4.0 | 26.3 | 4.0 | 26.3 | ||
| 34 | 5.0 | 55.6 | 5.0 | 55.6 | ||
| 35 | 5.0 | 55.6 | 5.0 | 55.6 | ||
| 36 | 55.6 | 55.6 | ||||
| 37 | 55.6 | 55.6 | ||||
| 38 | 55.6 | 55.6 | ||||
| 39 | 55.6 | 55.6 | ||||
| 40 | 55.6 | 55.6 |
| DF3 | DF22 | Significance Level (p) | |
|---|---|---|---|
| Growth Performance | |||
| Total length, 40 dph (mm) | 17.98 ± 1.690 b | 12.31 ± 0.393 a | * |
| CVTL, 40 dph | 12.3 ± 1.05 | 15.2 ± 4.08 | ns |
| SGRTL, 3–40 dph | 5.41 ± 0.253 b | 4.42 ± 0.076 a | * |
| TGCTL, 3–40 dph | 1.71 ± 0.110 b | 1.30 ± 0.030 a | * |
| Grading data (53 dph) | |||
| Survival (%) | 28.6 ± 0.82 | 30.9 ± 6.07 | ns |
| % Big fish | 61.1 ± 5.56 b | 9.0 ± 2.99 a | ** |
| % Small fish | 38.9 ± 5.56 a | 91.0 ± 2.99 b | ** |
| Weight (g) of Big fish | 0.72 ± 0.015 b | 0.60 ± 0.02 a | * |
| Weight (g) of Small fish | 0.47 ± 0.015 b | 0.28 ± 0.000 a | ** |
| DF3 | DF22 | Significance Level (p) | |
|---|---|---|---|
| Initial body weight (g) | 0.44 ± 0.010 b | 0.29 ± 0.010 a | *** |
| Final body weight (g) | 6.66 ± 0.116 b | 6.05 ± 0.172 a | * |
| Total length (cm) | 7.43 ± 0.011 | 7.15 ± 0.014 | ns |
| Standard length (cm) | 6.29 ± 0.022 | 6.01 ± 0.033 | ns |
| CFTL | 1.62 ± 0.011 | 1.66 ± 0.014 | ns |
| CFSL | 2.68 ± 0.022 a | 2.79 ± 0.033 b | ** |
| CV (%) | 25.1 ± 1.40 | 22.8 ± 1.44 | ns |
| WG (g/day) | 0.15 ± 0.003 b | 0.14 ± 0.004 a | * |
| SGR | 6.84 ± 0.054 | 6.87 ± 0.083 | ns |
| TGC | 1.02 ± 0.007 | 1.04 ± 0.015 | ns |
| Mortality (%) | 47.3 ± 2.62 | 45.8 ± 1.88 | ns |
| DF3 | DF22 | Significance Level (p) | |
|---|---|---|---|
| Normal | 80.8 ± 2.12 | 72.5 ± 4.54 | ns |
| Not-normal | 19.2 ± 2.12 | 27.5 ± 4.54 | ns |
| Gill operculum | 9.7 ± 1.57 | 6.2 ± 1.54 | ns |
| Fin malformation | 2.5 ± 0.84 | 3.6 ± 1.56 | ns |
| Jaw malformation | 0.9 ± 0.38 | 1.4 ± 0.44 | ns |
| Spinal cord malformation | 3.8 ± 0.63 a | 11.7 ± 1.42 b | *** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bougali, S.B.; Karakatsouli, N.; Ntomalis, K.; Kastelis, A.; Alexopoulou, V.-A.; Batzina, A.; Markakis, I. Effects of Co-Feeding Dry and Live Feed from the Onset of Exogenous Feeding on Red Seabream Pagrus major Larviculture and Pre-Growing. Fishes 2025, 10, 324. https://doi.org/10.3390/fishes10070324
Bougali SB, Karakatsouli N, Ntomalis K, Kastelis A, Alexopoulou V-A, Batzina A, Markakis I. Effects of Co-Feeding Dry and Live Feed from the Onset of Exogenous Feeding on Red Seabream Pagrus major Larviculture and Pre-Growing. Fishes. 2025; 10(7):324. https://doi.org/10.3390/fishes10070324
Chicago/Turabian StyleBougali, Sofia Brinkmann, Nafsika Karakatsouli, Kostas Ntomalis, Anemos Kastelis, Vasiliki-Anna Alexopoulou, Alkisti Batzina, and Isidoros Markakis. 2025. "Effects of Co-Feeding Dry and Live Feed from the Onset of Exogenous Feeding on Red Seabream Pagrus major Larviculture and Pre-Growing" Fishes 10, no. 7: 324. https://doi.org/10.3390/fishes10070324
APA StyleBougali, S. B., Karakatsouli, N., Ntomalis, K., Kastelis, A., Alexopoulou, V.-A., Batzina, A., & Markakis, I. (2025). Effects of Co-Feeding Dry and Live Feed from the Onset of Exogenous Feeding on Red Seabream Pagrus major Larviculture and Pre-Growing. Fishes, 10(7), 324. https://doi.org/10.3390/fishes10070324