
Survival Estimates of Endangered Shortnose Sturgeon (Acipenser brevirostrum Lesueur, 1818) from Geographically Disjunct Population Segments



Abstract
1. Introduction
2. Materials and Methods
2.1. Study Sites
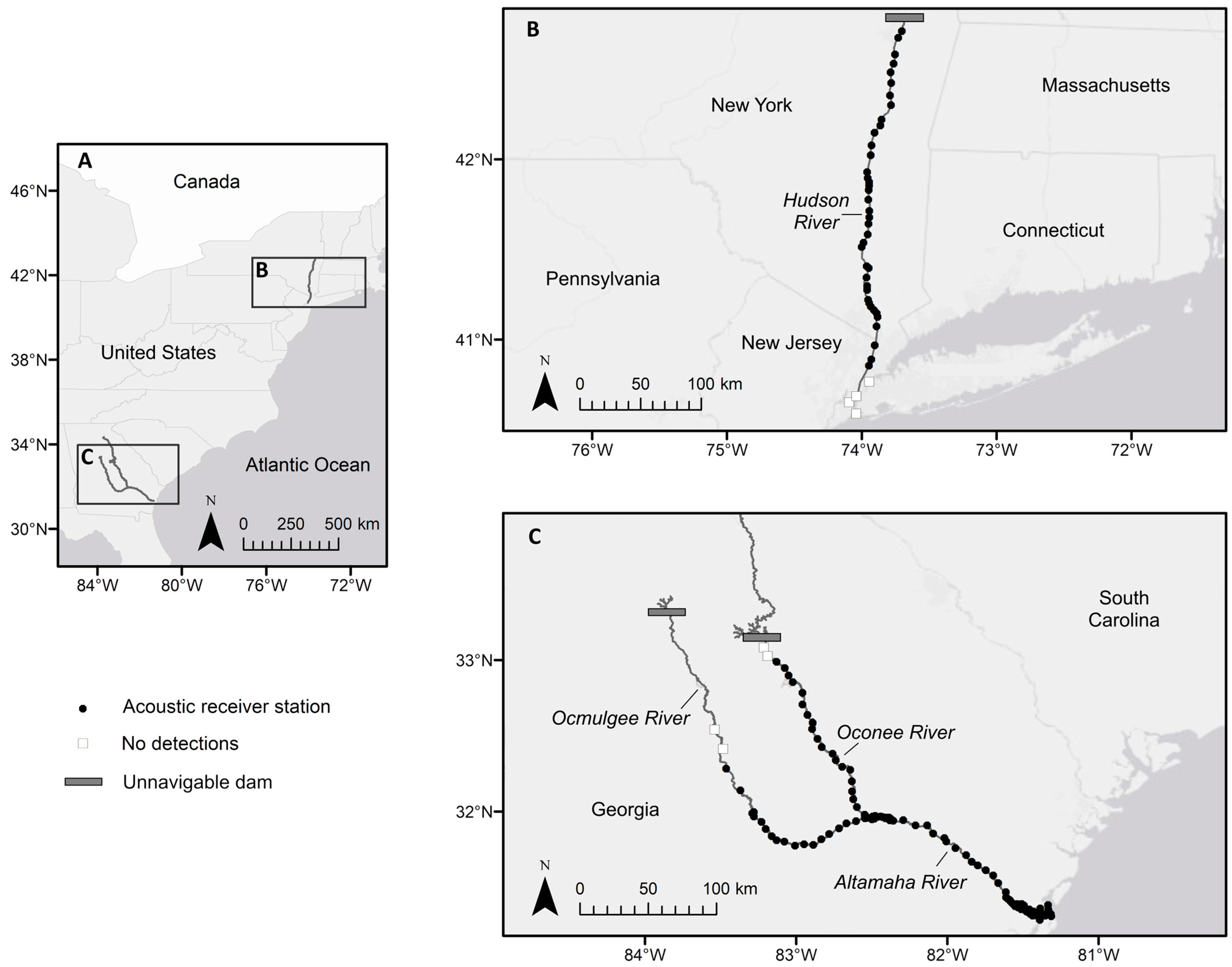
2.1.1. Hudson River
2.1.2. Altamaha River
2.2. Acoustic Telemetry Monitoring
2.3. Bayesian Multistate Capture–Recapture
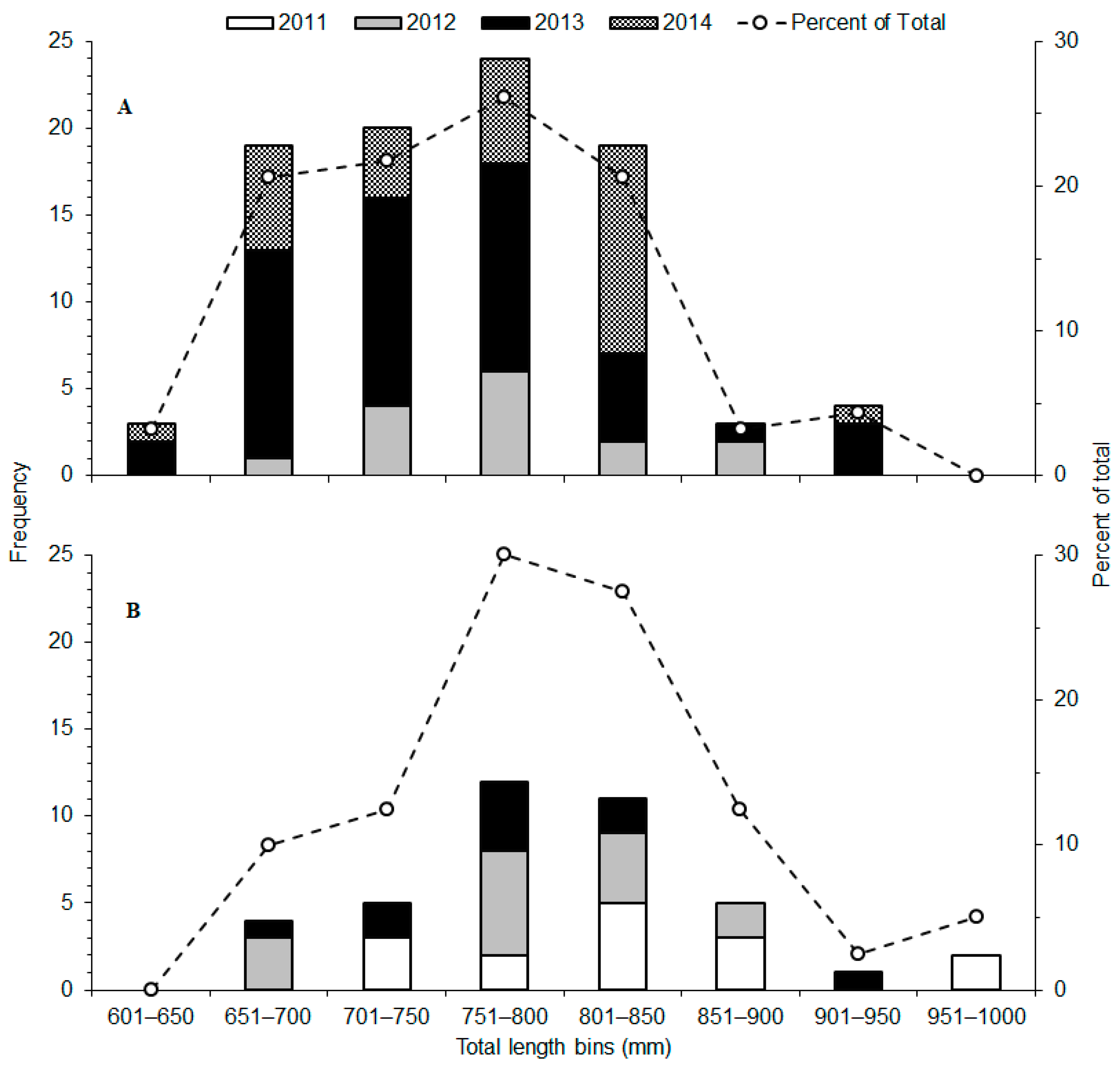
3. Results
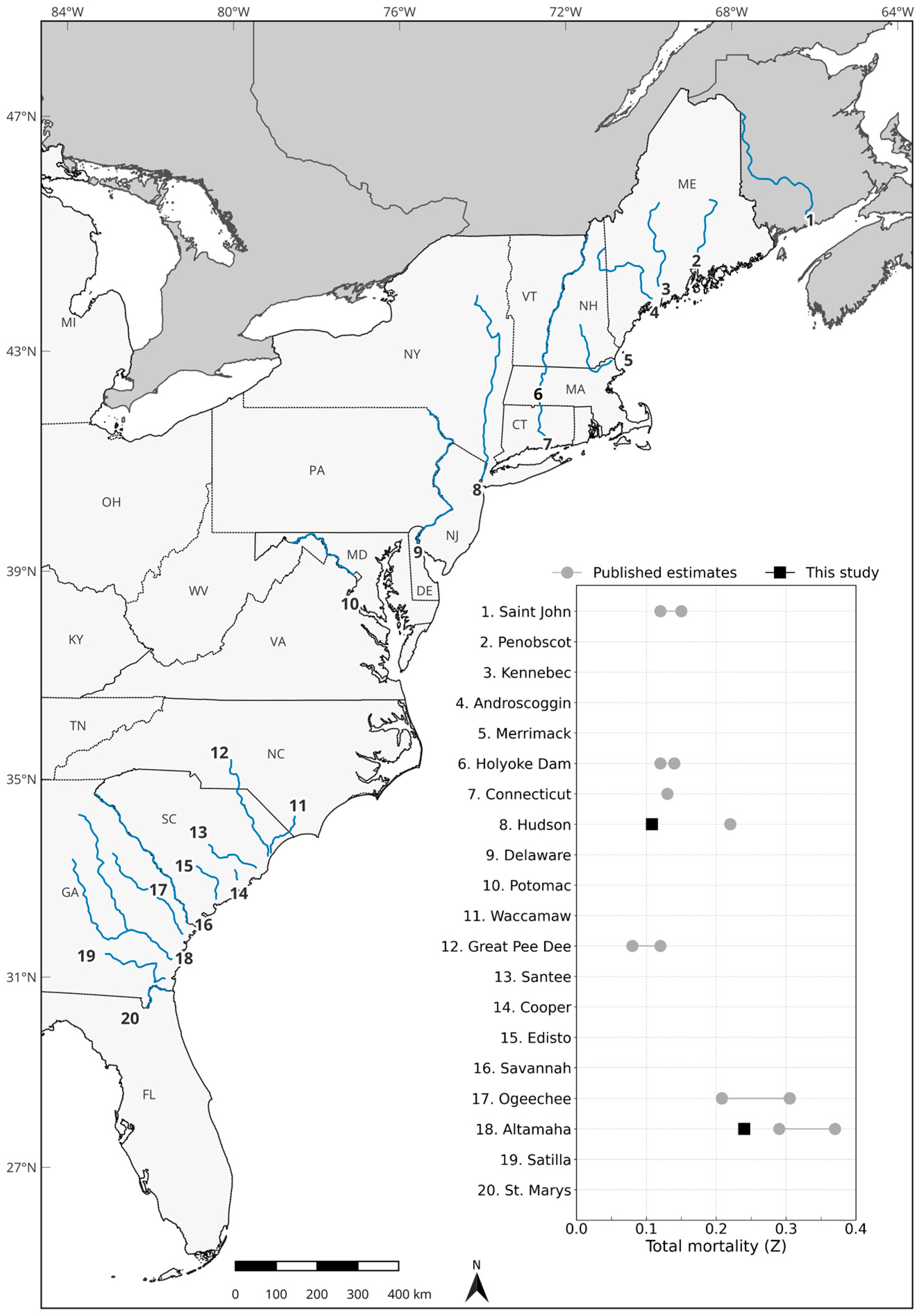
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CI | Bayesian credibility interval |
| DIC | Deviance information criterion |
| ESA | Endangered Species Act |
| FL | Fork length |
| MCMC | Markov chain Monte Carlo |
| TL | Total length |
References
- Birstein, V.J. Sturgeons and paddlefishes: Threatened fishes in need of conservation. Conserv. Biol. 1993, 7, 773–787. [Google Scholar] [CrossRef]
- Vladykov, V.D.; Greeley, J.R. Order Acipenseroidei. In Fishes of the Western North Atlantic. Part 3: Soft-Rayed Bony Fishes; Olsen, V.H., Ed.; Yale University: New Haven, CT, USA, 1963; pp. 24–60. [Google Scholar]
- Kynard, B. Life history, latitudinal patterns, and status of the shortnose sturgeon, Acipenser brevirostrum. Environ. Biol. Fishes 1997, 48, 319–334. [Google Scholar] [CrossRef]
- Kynard, B.; Bolden, S.; Kieffer, M.; Collins, M.; Brundage, H.; Hilton, E.J.; Litvak, M.; Kinnison, M.T.; King, T.; Peterson, D. Life history and status of shortnose sturgeon (Acipenser brevirostrum LeSeur, 1818). J. Appl. Ichthyol. 2016, 32, 208–248. [Google Scholar] [CrossRef]
- Dadswell, M.J. Biology and population characteristics of the shortnose sturgeon, Acipenser brevirostrum LeSueur 1818 (Osteichthyes: Acipenseridae), in the Saint John River estuary, New Brunswick, Canada. Can. J. Zool. 1979, 57, 2186–2210. [Google Scholar] [CrossRef]
- Rochard, E.; Castelnaud, G.; Lepage, M. Sturgeons (Pisces: Acipenseridae): Threats and prospects. J. Fish Biol. 1990, 37, 123–132. [Google Scholar] [CrossRef]
- Cooke, D.W.; Leach, S.D. Implications of a migration impediment on shortnose sturgeon spawning. N. Am. J. Fish. Manag. 2004, 24, 1460–1468. [Google Scholar] [CrossRef]
- Shortnose Sturgeon Status Review Team. A Biological Assessment of Shortnose Sturgeon (Acipenser brevirostrum). Report to National Marine Fisheries Service, Northeast Regional Office. 2010. Available online: https://repository.library.noaa.gov/view/noaa/17811 (accessed on 1 June 2022).
- Bain, M.B.; Haley, N.; Peterson, D.L.; Arend, K.K.; Mills, K.E.; Sullivan, P.J. Recovery of a US endangered fish. PLoS ONE 2007, 2, e168. [Google Scholar] [CrossRef]
- Altenritter, M.N.; Zydlewski, G.B.; Kinnison, M.T.; Zydlewski, J.D.; Wippelhauser, G.S. Understanding the basis of shortnose sturgeon (Acipenser brevirostrum) partial migration in the Gulf of Maine. Can. J. Fish. Aquat. Sci. 2018, 75, 464–473. [Google Scholar] [CrossRef]
- Fernandes, S.J.; Zydlewski, G.B.; Zydlewski, J.D.; Wippelhauser, G.S.; Kinnison, M.T. Seasonal distribution and movements of shortnose sturgeon and Atlantic Sturgeon in the Penobscot River estuary, Maine. Trans. Am. Fish. Soc. 2010, 139, 1436–1449. [Google Scholar] [CrossRef]
- King, T.L.; Henderson, A.P.; Kynard, B.E.; Kieffer, M.C.; Peterson, D.L.; Aunins, A.W.; Brown, B.L. A nuclear DNA perspective on delineating evolutionarily significant lineages in polyploids: The case of the endangered shortnose sturgeon (Acipenser brevirostrum). PLoS ONE 2014, 9, e102784. [Google Scholar] [CrossRef]
- Fleming, J.E.; Bryce, T.D.; Kirk, J.P. Age, growth, and status of shortnose sturgeon in the lower Ogeechee River, Georgia. Proc. Annu. Conf. SEAFWA 2003, 57, 80–91. [Google Scholar]
- Bain, M.B. Atlantic and shortnose sturgeons of the Hudson River: Common and divergent life history attributes. Environ. Biol. Fishes 1997, 48, 347–358. [Google Scholar] [CrossRef]
- O’herron, J.C.; Able, K.W.; Hastings, R.W. Movements of shortnose sturgeon (Acipenser brevirostrum) in the Delaware River. Estuaries 1993, 16, 235. [Google Scholar] [CrossRef]
- Ingram, E.C.; Peterson, D.L.; Fox, A.G. Abundance of endangered shortnose sturgeon (Acipenser brevirostrum) in the Altamaha River in Georgia. Fish. Bull. 2020, 118, 198–204. [Google Scholar] [CrossRef]
- Peterson, D.L.; Bednarski, M.S. Abundance and size structure of shortnose sturgeon in the Altamaha River, Georgia. Trans. Am. Fish. Soc. 2013, 142, 1444–1452. [Google Scholar] [CrossRef]
- Jager, H.I.; Peterson, D.L.; Farrae, D.; Bevelhimer, M.S. A population model to assess influences on the viability of the shortnose sturgeon population in the Ogeechee River, Georgia. Trans. Am. Fish. Soc. 2013, 142, 731–746. [Google Scholar] [CrossRef]
- Boreman, J. Sensitivity of North American sturgeons and paddlefish to fishing mortality. Environ. Biol. Fishes 1997, 48, 399–405. [Google Scholar] [CrossRef]
- Bahn, R.A.; Fleming, J.E.; Peterson, D.L. Bycatch of shortnose sturgeon in the commercial American shad fishery of the Altamaha River, Georgia. N. Am. J. Fish. Manag. 2012, 32, 557–562. [Google Scholar] [CrossRef]
- Collins, M.R.; Rogers, S.G.; Smith, T.I.J. Bycatch of sturgeons along the southern Atlantic Coast of the USA. N. Am. J. Fish. Manag. 1996, 16, 24–29. [Google Scholar] [CrossRef]
- Collins, M.R.; Rogers, S.G.; Smith, T.I.J.; Moser, M.L. Primary factors affecting sturgeon populations in the southeastern United States: Fishing mortality and degradation of essential habitats. Bull. Mar. Sci. 2000, 66, 917–928. [Google Scholar]
- Robson, D.S.; Chapman, D.G. Catch curves and mortality rates. Trans. Am. Fish. Soc. 1961, 90, 181–189. [Google Scholar] [CrossRef]
- Pauly, D. On the interrelationships between natural mortality, growth parameters, and mean environmental temperature in 175 fish stocks. J. Cons. Cons. Perm. Int. Pour Explor. Mer. 1980, 39, 175–192. [Google Scholar] [CrossRef]
- Lorenzen, K. The relationship between body weight and natural mortality in juvenile and adult fish: A comparison of natural ecosystems and aquaculture. J. Fish Biol. 1996, 49, 627–642. [Google Scholar] [CrossRef]
- Then, A.Y.; Hoenig, J.M.; Hall, N.G.; Hewitt, D.A. Evaluating the predictive performance of empirical estimators of natural mortality rate using information on over 200 fish species. ICES J. Mar. Sci. 2015, 72, 82–92. [Google Scholar] [CrossRef]
- Brownscombe, J.W.; Lédée, E.J.I.; Raby, G.D.; Struthers, D.P.; Gutowsky, L.F.G.; Nguyen, V.M.; Young, N.; Stokesbury, M.J.W.; Holbrook, C.M.; Brenden, T.O.; et al. Conducting and interpreting fish telemetry studies: Considerations for researchers and resource managers. Rev. Fish Biol. Fish. 2019, 29, 369–400. [Google Scholar] [CrossRef]
- Kahn, J.E.; Hager, C.; Watterson, J.C.; Mathies, N.; Deacy, A.; Hartman, K.J. Population and sex-specific survival estimates for Atlantic Sturgeon: Addressing detection probability and tag loss. Aquat. Biol. 2023, 32, 1–12. [Google Scholar] [CrossRef]
- Hightower, J.E.; Loeffler, M.; Post, W.C.; Peterson, D.L. Estimated survival of subadult and adult Atlantic Sturgeon in four river basins in the southeastern United States. Mar. Coast. Fish. 2015, 7, 514–522. [Google Scholar] [CrossRef]
- Melnychuk, M.C.; Dunton, K.J.; Jordaan, A.; McKown, K.A.; Frisk, M.G. Informing conservation strategies for the endangered Atlantic Sturgeon using acoustic telemetry and multi-state mark–recapture models. J. Appl. Ecol. 2016, 54, 914–925. [Google Scholar] [CrossRef]
- Lindley, S.T.; Moser, M.L.; Erickson, D.L.; Belchik, M.; Welch, D.W.; Rechisky, E.L.; Kelly, J.T.; Heublein, J.; Klimley, A.P. Marine migration of North American green sturgeon. Trans. Am. Fish. Soc. 2008, 137, 182–194. [Google Scholar] [CrossRef]
- Rudd, M.B.; Ahrens, R.N.M.; Pine III, W.E.; Bolden, S.K. Empirical, spatially explicit natural mortality and movement rate estimates for the threatened Gulf Sturgeon (Acipenser oxyrinchus desotoi). Can. J. Fish. Aquat. Sci. 2014, 71, 1407–1417. [Google Scholar] [CrossRef]
- Colborne, S.F.; Hayden, T.A.; Holbrook, C.M.; Hondorp, D.W.; Krueger, C.C. Lake sturgeon (Acipenser fulvescens) annual adult survival estimated from acoustic telemetry. J. Gt. Lakes Res. 2021, 47, 1814–1823. [Google Scholar] [CrossRef]
- Ingram, E.C.; Peterson, D.L. Seasonal movements of shortnose sturgeon (Acipenser brevirostrum) in the Altamaha River, Georgia. River Res. Appl. 2018, 34, 873–882. [Google Scholar] [CrossRef]
- Coch, N.K.; Bokuniewicz, H.J. Oceanographic and geologic framework of the Hudson System. Northeast. Geol. 1986, 8, 96–108. [Google Scholar]
- Olsen, C.R.; Simpson, H.J.; Peng, T.-H.; Bopp, R.F.; Trier, R.M. Sediment mixing and accumulation rate effects on radionuclide depth profiles in Hudson Estuary sediments. J. Geophys. Res. Oceans 1981, 86, 11020–11028. [Google Scholar] [CrossRef]
- de Vries, M.P.; Weiss, L.A. Salt-Front Movement in the Hudson River Estuary, New York—Simulations by One-Dimensional Flow and Solute-Transport Models; Water-Resources Investigations Report 99–4024; U.S. Geological Survey: Reston, VA, USA, 2001; 69p. Available online: https://pubs.er.usgs.gov/publication/wri994024 (accessed on 1 June 2022).
- Blumberg, A.F.; Hellweger, F.L. Hydrodynamics of the Hudson River estuary. Am. Fish. Soc. Symp. 2006, 51, 9–28. [Google Scholar]
- Seekell, D.A.; Pace, M.L. Climate change drives warming in the Hudson River estuary, New York (USA). J. Environ. Monit. 2011, 13, 2321. [Google Scholar] [CrossRef]
- Dynesius, M.; Nilsson, C. Fragmentation and flow regulation of river systems in the northern third of the world. Science 1994, 266, 753–762. [Google Scholar] [CrossRef]
- Sheldon, J.E.; Alber, M. A Comparison of residence time calculations using simple compartment models of the Altamaha River estuary, Georgia. Estuaries 2002, 25, 1304–1317. [Google Scholar] [CrossRef]
- Pendleton, R.M.; Standley, C.R.; Higgs, A.L.; Kenney, G.H.; Sullivan, P.J.; Sethi, S.A.; Harris, B.P. Acoustic telemetry and benthic habitat mapping inform the spatial ecology of shortnose sturgeon in the Hudson River, New York, USA. Trans. Am. Fish. Soc. 2019, 148, 35–47. [Google Scholar] [CrossRef]
- Scott, W.B.; Crossman, E.J. Freshwater Fishes of Canada, 1st ed.; Fisheries Research Board of Canada: Ottawa, ON, Canada, 1973. [Google Scholar]
- Dadswell, M.J.; Taubert, B.D.; Squiers, T.S.; Marchette, D.; Buckley, J. Synopsis of Biological Data on Shortnose Sturgeon, Acipenser brevirostrum LeSueur 1818; FAO Fish Synopsis 140; National Oceanic and Atmospheric Administration: Washington, DC, USA, 1984. [Google Scholar]
- Boone, S.S.; Hernandez, S.M.; Camus, A.C.; Peterson, D.L.; Jennings, C.A.; Shelton, J.L.; Divers, S.J. Evaluation of four suture materials for surgical incision closure in Siberian sturgeon. Trans. Am. Fish. Soc. 2013, 142, 649–659. [Google Scholar] [CrossRef]
- Breece, M.W.; Higgs, A.L.; Fox, D.A. Spawning intervals, timing, and riverine habitat use of adult Atlantic Sturgeon in the Hudson River. Trans. Am. Fish. Soc. 2021, 150, 528–537. [Google Scholar] [CrossRef]
- Ingram, E.C.; Peterson, D.L. Annual spawning migrations of adult Atlantic Sturgeon in the Altamaha River, Georgia. Mar. Coast. Fish. Dyn. Manag. Ecosyst. Sci. 2016, 8, 595–606. [Google Scholar] [CrossRef]
- Hightower, J.E.; Harris, J.E. Estimating fish mortality rates using telemetry and multistate models. Fisheries 2017, 42, 210–219. [Google Scholar] [CrossRef]
- Kéry, M.; Schaub, M. Bayesian Population Analysis Using WinBUGS: A Hierarchical Perspective; Academic Press: Cambridge, MA, USA, 2012. [Google Scholar]
- King, R. A review of Bayesian state-space modelling of capture-recapture-recovery data. Interface Focus 2012, 2, 190–204. [Google Scholar] [CrossRef]
- Gimenez, O.; Rossi, V.; Choquet, R.; Dehais, C.; Doris, B.; Varella, H.; Vila, J.-P.; Pradel, R. State-space modelling of data on marked individuals. Ecol. Model. 2007, 206, 431–438. [Google Scholar] [CrossRef]
- Arnason, A.N. The estimation of population size, migration rates and survival in a stratified population. Res. Popul. Ecol. 1973, 15, 1–8. [Google Scholar] [CrossRef]
- Kendall, W.L.; Nichols, J.D. On the use of secondary capture-recapture samples to estimate temporary emigration and breeding proportions. J. Appl. Stat. 1995, 22, 751–762. [Google Scholar] [CrossRef]
- White, G.C.; Kendall, W.L.; Barker, R.J. Multistate survival models and their extensions in Program MARK. J. Wildl. Manag. 2006, 70, 1521–1529. [Google Scholar] [CrossRef]
- Plummer, M. JAGS: A program for analysis of Bayesian graphical models using Gibbs sampling. In Proceedings of the 3rd International Workshop on Distributed Statistical Computing, Vienna, Austria, 20–22 March 2003; pp. 1–10. [Google Scholar]
- Su, Y.; Yajima, M. R2jags: A Package for Running JAGS from R, 2014. Available online: http://CRAN.R-project.org/package=R2jags (accessed on 1 June 2022).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Nelson, T.R.; Powers, S.P. Estimates of red drum mortality via acoustic telemetry. Mar. Coast. Fish. 2020, 12, 78–97. [Google Scholar] [CrossRef]
- Calvert, A.M.; Bonner, S.J.; Jonsen, I.D.; Flemming, J.M.; Walde, S.J.; Taylor, P.D. A hierarchical Bayesian approach to multi-state mark–recapture: Simulations and applications. J. Appl. Ecol. 2009, 46, 610–620. [Google Scholar] [CrossRef]
- Gelman, A.; Rubin, D.B. Inference from iterative simulation using multiple sequences. Stat. Sci. 1992, 7, 457–472. [Google Scholar] [CrossRef]
- Spiegelhalter, D.J.; Best, N.G.; Carlin, B.P.; Van Der Linde, A. Bayesian measures of model complexity and fit. J. R. Stat. Soc. Ser. B Stat. Methodol. 2002, 64, 583–639. [Google Scholar] [CrossRef]
- Hooten, M.B.; Hobbs, N.T. A guide to Bayesian model selection for ecologists. Ecol. Monogr. 2015, 85, 3–28. [Google Scholar] [CrossRef]
- Gelman, A.; Carlin, J.B.; Stern, H.S.; Dunson, D.B.; Vehtari, A.; Rubin, D.B. Bayesian Data Analysis, 3rd ed.; Chapman & Hall/CRC Texts in Statistical Science; CRC Press: Boca Raton, FL, USA, 2013. [Google Scholar]
- Dovel, W.L. The Endangered Shortnose Sturgeon of the Hudson Estuary: Its Life History and Vulnerability to the Activities of Man; U.S. Department of Energy: Washington, DC, USA, 1981. [Google Scholar]
- Kruschke, J.K. Bayesian analysis reporting guidelines. Nat. Hum. Behav. 2021, 5, 1282–1291. [Google Scholar] [CrossRef] [PubMed]
- Shortnose Sturgeon Recovery Team. Final Recovery Plan for the Shortnose Sturgeon (Acipenser brevirostrum); National Marine Fisheries Service: Silver Spring, MD, USA, 1998. [Google Scholar]
- Perry, R.W.; Castro-Santos, T.; Holbrook, C.M.; Sandford, B.P. Using mark-recapture models to estimate survival from telemetry data. In Telemetry Techniques: A User Guide for Fisheries Research; Adams, N.S., Beeman, J.W., Eiler, J.H., Eds.; American Fisheries Society: Bethesda, MD, USA, 2012; pp. 453–475. [Google Scholar]
- Morris, W.F.; Doak, D.F. Quantitative Conservation Biology: Theory and Practice of Population Viability Analysis; Sinauer Associates: Sunderland, MA, USA, 2003. [Google Scholar]
- Taubert, B.D. Reproduction of shortnose sturgeon (Acipenser brevirostrum) in Holyoke Pool, Connecticut River, Massachusetts. Copeia 1980, 1980, 114–117. [Google Scholar] [CrossRef]
- Woodland, R.J.; Secor, D.H. Year-class strength and recovery of endangered shortnose sturgeon in the Hudson River, New York. Trans. Am. Fish. Soc. 2007, 136, 72–81. [Google Scholar] [CrossRef]
- Peterson, D.L.; Farrae, D.J. Evidence of metapopulation dynamics in shortnose sturgeon in the southern part of their range. Trans. Am. Fish. Soc. 2011, 140, 1540–1546. [Google Scholar] [CrossRef]
- DeVries, R.J. Population Dynamics, Movements, and Spawning Habitat of the Shortnose Sturgeon, Acipenser Brevirostrum, in the Altamaha River System, Georgia. Master’s Thesis, University of Georgia, Athens, GA, USA, 2006. [Google Scholar]
- Ziegeweid, J.R.; Jennings, C.A.; Peterson, D.L.; Black, M.C. Effects of salinity, temperature, and weight on the survival of young-of-year shortnose sturgeon. Trans. Am. Fish. Soc. 2008, 137, 1490–1499. [Google Scholar] [CrossRef]
- Pörtner, H.O.; Peck, M.A. Climate change effects on fishes and fisheries: Towards a cause-and-effect understanding. J. Fish Biol. 2010, 77, 1745–1779. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Masson-Delmotte, V., Zhai, P., Pirani, A., Connors, S.L., Péan, C., Berger, S., Caud, N., Chen, Y., Goldfarb, L., Gomis, M.I., et al., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2023. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| FL (mm) | TL (mm) | Detections | ||||||||
| River | Tag Type | Battery (d) | Release Year | No. Tagged | Mean | Range [SD] | Mean | Range [SD] | Total | Range |
| Hudson | Lotek MM-MR-16-50 | 389 | 2012 | 14 | 673 | 590–759 [49.4] | 777 | 694–871 [54.8] | 389,792 | 12,932–69,027 |
| 2013 | 47 | 647 | 545–801 [60.2] | 749 | 639–942 [69.4] | 555,945 | 110–49,517 | |||
| 2014 | 30 | 675 | 566–826 [61.0] | 765 | 633–918 [67.5] | 281,359 | 29–49,555 | |||
| All years | 91 | 661 | 545–826 [59.9] | 759 | 633–942 [66.9] | 1,227,096 | 29–69,027 | |||
| Altamaha | VEMCO V16-4H | 1248 | 2011 | 15 | 726 | 635–830 [60.6] | 828 | 732–973 [75.2] | 1,698,833 | 5266–296,130 |
| 2012 | 15 | 690 | 670–800 [52.4] | 784 | 658–897 [64.1] | 1,256,751 | 10,442–141,132 | |||
| 2013 | 10 | 681 | 574–819 [66.2] | 776 | 656–945 [79.7] | 208,916 | 8459–39,022 | |||
| All years | 40 | 701 | 574–830 [60.9] | 799 | 656–973 [74.3] | 3,164,500 | 5266–296,130 | |||
| Overall | 131 | 673 | 545–830 [62.9] | 771 | 633–973 [71.4] | 4,391,596 | 29–296,130 | |||
| Model | p | S | Fish TL Slope | Annual Survival | Mortality | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| River | No. Tagged | TL (mm) | p | S | pD | DIC | Estimate | 95% CI | Estimate | 95% CI | Estimate | 95% CI | Estimate | 95% CI | Z a | M b |
| Hudson | 91 | 633–942 | Time | Constant | 211.1 | 1137.6 | 0.602 | 0.582–0.621 | 0.991 | 0.984–0.996 | - | - | 0.897 | 0.828–0.953 | 0.108 | 0.089 |
| Time | Size | 214.0 | 1150.4 | 0.601 | 0.581–0.620 | 0.993 | 0.986–0.998 | 0.012 | −0.002–0.031 | 0.923 | 0.848–0.983 | |||||
| Altamaha | 40 | 656–973 | Time | Constant | 122.3 | 793.1 | 0.821 | 0.792–0.848 | 0.980 | 0.969–0.989 | - | - | 0.787 | 0.685–0.874 | 0.240 | 0.224 |
| Time | Size | 122.4 | 793.6 | 0.821 | 0.792–0.848 | 0.981 | 0.970–0.989 | −0.001 | −0.007–0.006 | 0.793 | 0.692–0.880 | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ingram, E.C.; Higgs, A.L.; Butler, L.; Fox, D.A.; Fox, A.G. Survival Estimates of Endangered Shortnose Sturgeon (Acipenser brevirostrum Lesueur, 1818) from Geographically Disjunct Population Segments. Fishes 2025, 10, 293. https://doi.org/10.3390/fishes10060293
Ingram EC, Higgs AL, Butler L, Fox DA, Fox AG. Survival Estimates of Endangered Shortnose Sturgeon (Acipenser brevirostrum Lesueur, 1818) from Geographically Disjunct Population Segments. Fishes. 2025; 10(6):293. https://doi.org/10.3390/fishes10060293
Chicago/Turabian StyleIngram, Evan C., Amanda L. Higgs, Liam Butler, Dewayne A. Fox, and Adam G. Fox. 2025. "Survival Estimates of Endangered Shortnose Sturgeon (Acipenser brevirostrum Lesueur, 1818) from Geographically Disjunct Population Segments" Fishes 10, no. 6: 293. https://doi.org/10.3390/fishes10060293
APA StyleIngram, E. C., Higgs, A. L., Butler, L., Fox, D. A., & Fox, A. G. (2025). Survival Estimates of Endangered Shortnose Sturgeon (Acipenser brevirostrum Lesueur, 1818) from Geographically Disjunct Population Segments. Fishes, 10(6), 293. https://doi.org/10.3390/fishes10060293