
Increased CO2 Concentration Mitigates the Impact of Nitrite on Zebrafish (Danio rerio) Liver and Gills


 and
and 
Abstract
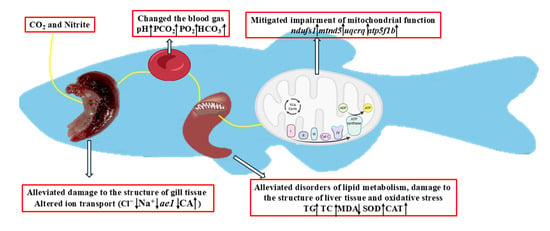
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Zebrafish Rearing and Exposure Experiments
2.3. Blood Gas and Hb/MetHb Quantification
2.4. Histopathological Analysis
2.5. Biochemical Indicator Assay
2.6. RNA Extraction and qPCR
2.7. Statistical Analyses
3. Results
3.1. Nitrite Accumulation in Liver and Gills
3.2. Hematological and Blood Gas Parameters
3.2.1. Hb and MetHb Levels in Blood
3.2.2. Blood Gas Analysis
3.3. Effects of Nitrite Co-Treatment with Carbon Dioxide on Gills
3.3.1. Morphological Changes in Gill Tissues
3.3.2. Ion Transport Changes in Gills
3.3.3. Changes in rMO2
3.4. Effects of Nitrite Co-Treatment with Carbon Dioxide on Liver
3.4.1. Morphological Changes in Liver Tissue
3.4.2. Oxidative-Antioxidant Parameter Analysis
3.4.3. Profiles of Genes Related to Mitochondrial Respiratory Chain
3.4.4. Lipid Metabolism Analysis in the Liver
4. Discussion
4.1. Increased CO2 Concentration Mitigates Toxic Effects of Nitrite on Gills
4.2. The Toxic Effect of Nitrite on the Liver Was Mitigated by the Increase in CO2 Concentration
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
Appendix A.1
Appendix A.2
Appendix A.3

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Accession No. | Primer Sequences (from 5′ to 3′) | Amplification Efficiency (%) |
|---|---|---|---|
| ae1 | NM_198338 | F: GGAGGACTTCTTGCGGATAAG | 108.4 |
| R: CCTATGACGAGAACTGGTTGAG | |||
| nbc1 | XM_009295136 | F: AGGAAACAGCCAGATGGATAAA | 109.9 |
| R: GAAGAGCGAGTGCAGAGAAA | |||
| nhe1 | NM_001113480 | F: GGTCCTGTATCACCTGTTTGAG | 109.4 |
| R: CCACAGACACCACGAAGAAA | |||
| nka | NM_131686 | F: TGTGCTTTCTGCTGTCGTGA | 102.9 |
| R: ATAACCAAGGCTTGCTGGGG | |||
| nhe3b | NM_001113479.1 | F: TCACTGTCATTCTGCAGGGTA | 97.9 |
| R: GGGTCAGATCGGACTGCG | |||
| nkcc | XM_021467734 | F: CTGCGAGAGGGACTTGATATTT | 92.6 |
| R: CTCCATCCGAGTCTTTGCTTAT | |||
| ndufs1 | NM_001007765.1 | F: GAAGATGTCCTTACGCGTGT | 101.8 |
| R: TTCAGCAGGTCCTTCAGAGC | |||
| mtnd5 | NP_059341 | F: TTCAGCAGGTCCTTCAGAGC | 110 |
| R: AGATGGGAGGTGGTTATTGC | |||
| mtcyb | NP_059343 | F: TAGACAATGCAACCCTTACA | 101.4 |
| R: CGGTTTCATGGAGAAATAGC | |||
| uqcrq | NM_001002495.3 | F: ATTTGTGGATTCGATTTAGG | 104.4 |
| R: ATGATTGCCCCATGTGTAGG | |||
| atp5f1b | NM_001024429.2 | F: CTCAATGCCCTGGAAGTAGC | 108.3 |
| R: GAACCTTCTGACCACGAACC | |||
| mtatp8 | NP_059335 | F: ATGCCTCAGCTTAATCCAAAA | 109.9 |
| R: ATCAACTTGAGTTGGGTCATTAG | |||
| gapdh | NM_001115114 | F:CTGGTGACCCGTGCTGCTT | 98.1 |
| R: TTTGCCGCCTTCTGCCTTA | |||
| fasn | XM_009306806 | F: GAGAAAGCTTGCCAAACAGG | 90.7 |
| R: GAGGGTCTTGCAGGAGACAG | |||
| acaca | NM_001271308 | F: GGACGGACCCTTGCACAATA | 108.4 |
| R: CCTCTGCAGGTCGATACGTC | |||
| atgl | XM_005174256 | F: ACACACTTACACCGCGTGAT | 108.8 |
| R: AGCACGTTTTCTCCATCCGT | |||
| hsla | NM_001316725 | F: AGGTAAGCAAAGGTTGTCCGA | 102.9 |
| R: TTCATGACCCCCAACAGACG | |||
| cpt1aa | NM_001044854 | F: TCTACCTGAGAGGTCGTGGG | 89.9 |
| R: TGACGTTTCCTGCTCTTGCT |
| Group | Time | pH | Dissolved Oxygen (mg/L) | Temperature (°C) | Nitrite (mg/L) | CO2 (mg/L) | HCO3− Content (mg/L) |
|---|---|---|---|---|---|---|---|
| Control | 12 h | 7.85 ± 0.08 | 6.61 ± 0.48 | 25.5 ± 0.17 | 0.01 ± 0 | 1.02 ± 0.06 | 166.79 ± 0.94 |
| Nitrite | 12 h | 7.78 ± 0.06 | 6.89 ± 0.02 | 25.57 ± 0.25 | 1.88 ± 0.03 | 1.02 ± 0.03 | 166.38 ± 2.54 |
| Nitrite-2.5 CO2 | 12 h | 7.59 ± 0.04 | 6.91 ± 0.06 | 25.60 ± 0.10 | 1.88 ± 0.01 | 2.18 ± 0.07 | 170.86 ± 1.22 |
| Nitrite-5 CO2 | 12 h | 7.43 ± 0.04 | 6.66 ± 0.13 | 25.37 ± 0.06 | 1.9 ± 0.01 | 4.47 ± 0.11 | 183.47 ± 1.27 |
| Nitrite-10 CO2 | 12 h | 7.10 ± 0.05 | 6.53 ± 0.11 | 25.50 ± 0.26 | 1.91 ± 0.03 | 8.54 ± 0.07 | 201.16 ± 5.12 |
| Control | 24 h | 7.79 ± 0.05 | 5.84 ± 0.06 | 25.58 ± 0.07 | 0.01 ± 0 | 1.07 ± 0.01 | 167.4 ± 0.94 |
| Nitrite | 24 h | 7.80 ± 0.05 | 5.79 ± 0.12 | 25.33 ± 0.06 | 1.78 ± 0.03 | 1.00 ± 0.03 | 166.58 ± 1.06 |
| Nitrite-2.5 CO2 | 24 h | 7.62 ± 0.08 | 5.94 ± 0.12 | 25.53 ± 0.12 | 1.81 ± 0.02 | 1.77 ± 0.10 | 173.5 ± 3.13 |
| Nitrite-5 CO2 | 24 h | 7.39 ± 0.03 | 5.88 ± 0.14 | 25.40 ± 0.10 | 1.83 ± 0.01 | 3.30 ± 0.10 | 183.67 ± 2.20 |
| Nitrite-10 CO2 | 24 h | 7.01 ± 0.05 | 5.69 ± 0.18 | 25.37 ± 0.06 | 1.85 ± 0.03 | 6.06 ± 0.15 | 202.18 ± 3.07 |
Appendix B


References
- FAO. The State of World Fisheries and Aquaculture 2018-Meeting the Sustainable Development Goals; Food and Agriculture Organization of the United Nations: Rome, Italy, 2018. [Google Scholar]
- Manan, H.; Zhong, J.M.H.; Kasan, N.A.; Suratman, S.; Ikhwanuddin, M. Carbon dioxide flux from intensive aquaculture shrimp farming applying biofloc system of Setiu Terengganu, Malaysia. Aquaculture 2019, 509, 52–58. [Google Scholar] [CrossRef]
- Grosell, M.; Jensen, F.B. Uptake and effects of nitrite in the marine teleost fish Platichthys flesus. Aquat. Toxicol. 2000, 50, 97–107. [Google Scholar] [CrossRef]
- Jensen, F.B. Nitrite disrupts multiple physiological functions in aquatic animals. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2003, 135, 9–24. [Google Scholar] [CrossRef] [PubMed]
- Ha, N.T.K.; Huong, D.T.T.; Phuong, N.T.; Bayley, M.; Jensen, F.B. Impact and tissue metabolism of nitrite at two acclimation temperatures in striped catfish (Pangasianodon hypophthalmus). Aquat. Toxicol. 2019, 212, 154–161. [Google Scholar] [CrossRef]
- Yang, L.P.; Guo, H.H.; Kuang, Y.; Yang, H.; Zhang, X.; Tang, R.; Li, D.P.; Li, L. Neurotoxicity induced by combined exposure of microcystin-LR and nitrite in male zebrafish (Danio rerio): Effects of oxidant-antioxidant system and neurotransmitter system. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2021, 253, 109248. [Google Scholar] [CrossRef]
- Huong, D.T.T.; Gam, L.T.H.; Lek, S.; Ut, V.N.; Phuong, N.T. Effects of nitrite at different temperatures on physiological parameters and growth in clown knifefish (Chitala ornata, Gray 1831). Aquaculture 2020, 521, 735060. [Google Scholar] [CrossRef]
- Zhang, M.Z.; Yin, X.L.; Li, M.; Wang, R.X.; Qian, Y.X.; Hong, M.L. Effect of nitrite exposure on haematological status, oxidative stress, immune response and apoptosis in yellow catfish (Pelteobagrus fulvidraco). Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2020, 238, 108867. [Google Scholar] [CrossRef]
- Yang, H.; Ou-Yang, K.; He, Y.; Wang, X.Y.; Wang, L.M.; Yang, Q.; Li, D.P.; Li, L. Nitrite induces hepatic glucose and lipid metabolism disorders in zebrafish through mitochondrial dysfunction and ERs response. Aquat. Toxicol. 2024, 273, 107015. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.H.; Su, Y.L.; Ma, H.L.; Deng, Y.Q.; Feng, J.; Chen, X.L.; Jie, Y.K.; Guo, Z.X. Effect of nitrite exposure on oxidative stress, DNA damage and apoptosis in mud crab (Scylla paramamosain). Chemosphere 2019, 239, 124668. [Google Scholar] [CrossRef]
- Gao, X.Q.; Fei, F.; Huo, H.H.; Huang, B.; Meng, X.S.; Zhang, T.; Liu, B.L. Effect of acute exposure to nitrite on physiological parameters, oxidative stress, and apoptosis in Takifugu rubripes. Ecotoxicol. Environ. Saf. 2019, 188, 109878. [Google Scholar] [CrossRef]
- Ministry of Agriculture and Rural Affairs of the People’s Republic of China. Water Quality Standard for Fisheries (GB 11607-89). National Environmental Protection Agency, Beijing. 1989. Available online: https://www.mee.gov.cn/ywgz/fgbz/bz/bzwb/shjbh/shjzlbz/199003/t19900301_66502.shtml (accessed on 14 April 2025).
- Buřič, M.; Blahovec, J.; Kouřil, J. Feasibility of open recirculating system in temperate climate—A case study. Aquac. Res. 2016, 47, 1156–1167. [Google Scholar] [CrossRef]
- Kamstra, A.; Span, J.; Weerd, J.V. The acute toxicity and sublethal effects of nitrite on growth and feed utilization of European eel, Anguilla (L.). Aquac. Res. 1996, 27, 903–911. [Google Scholar] [CrossRef]
- Tacon, A.; Cody, J.; Conquest, L.; Divakaran, S.; Forster, I.; Decamp, O. Effect of culture system on the nutrition and growth performance of Pacific white shrimp Litopenaeus vannamei (Boone) fed different diets. Aquac. Nutr. 2002, 8, 121–137. [Google Scholar] [CrossRef]
- Yang, P.; Lai, D.F.; Yang, H.; Tong, C. Carbon dioxide dynamics from sediment, sediment-water interface and overlying water in the aquaculture shrimp ponds in subtropical estuaries, southeast China. J. Environ. Manag. 2019, 236, 224–235. [Google Scholar] [CrossRef] [PubMed]
- Gorle, J.M.R.; Terjesen, B.F.; Mota, V.C.; Summerfelt, S. Water velocity in commercial RAS culture tanks for Atlantic salmon smolt production. Aquac. Eng. 2018, 81, 89–100. [Google Scholar] [CrossRef]
- Summerfelt, S.; Bebak-Williams, J.; Tsukuda, S. Controlled systems: Water reuse and recirculation. Fish Hatch. Manag. 2001, 40, 285–395. [Google Scholar]
- Foucreau, N.; Cottin, D.; Piscart, C.; Hervant, F. Physiological and metabolic responses to rising temperature in Gammarus pulex (Crustacea) populations living under continental or Mediterranean climates. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2014, 168, 69–75. [Google Scholar] [CrossRef]
- Sundin, J.; Jutfelt, F. Effects of elevated carbon dioxide on male and female behavioral lateralization in a temperate goby. Roy. Soc. Open. Sci. 2018, 5, 171550. [Google Scholar] [CrossRef]
- Nasuchon, N.; Yagi, M.; Kawabata, Y.; Gao, K.; Ishimatsu, A. Escape responses of the Japanese anchovy Engraulis japonicus under elevated temperature and CO2 conditions. Fish Sci. 2016, 82, 435–444. [Google Scholar] [CrossRef]
- Guo, T.; Wang, Y.F.; Li, J.Y.; Guo, X.Y.; Xu, S.H.; Han, H.W.; Yu, J.C.; Li, J.; Liu, Q. Accumulated CO2 affects growth, acid-base regulation and ion balance of turbot (Scophthalmus maximus) in a recirculating aquaculture system. Aquaculture 2023, 578, 740070. [Google Scholar] [CrossRef]
- Mota, V.C.; Nilsen, T.O.; Gerwins, J.; Gallo, M.; Ytteborg, E.; Baeverfjord, G.; Kolarevic, J.; Summerfelt, S.T.; Terjesen, B.F. The effects of carbon dioxide on growth performance, welfare, and health of Atlantic salmon post-smolt (Salmo salar) in recirculating aquaculture systems. Aquaculture 2019, 498, 578–586. [Google Scholar] [CrossRef]
- Sadoul, B.; Friggens, N.; Valotaire, C.; Labbé, L.; Colson, V.; Prunet, P.; Leguen, I. Physiological and behavioral flexibility to an acute CO2 challenge, within and between genotypes in rainbow trout. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2017, 209, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Evans, D.H.; Piermarini, P.M.; Choe, K.P. The multifunctional fish gill: Dominant site of gas exchange, osmoregulation, acid-base regulation, and excretion of nitrogenous waste. Physiol. Rev. 2005, 85, 97–177. [Google Scholar] [CrossRef]
- Perry, S.; Gilmour, K. Acid–base balance and CO2 excretion in fish: Unanswered questions and emerging models. Respir. Physiol. Neurobiol. 2006, 154, 199–215. [Google Scholar] [CrossRef] [PubMed]
- Hwang, P.P. Ion uptake and acid secretion in zebrafish (Danio rerio). J. Exp. Biol. 2009, 212, 1745–1752. [Google Scholar] [CrossRef]
- Sundin, J.; Morgan, R.; Finnøen, M.H.; Dey, A.; Sarkar, K.; Jutfelt, F. On the observation of wild zebrafish (Danio rerio) in India. Zebrafish 2019, 16, 546–553. [Google Scholar] [CrossRef]
- Tarafder, P.K.; Rathore, D. Spectrophotometric determination of nitrite in water. Analyst 1988, 113, 1073–1076. [Google Scholar] [CrossRef]
- Guo, H.H.; Lin, W.; Wu, X.Y.; Wang, L.K.; Zhang, D.D.; Li, L.; Li, D.P.; Tang, R.; Yang, L.P.; Qiu, Y.M. Survival strategies of Wuchang bream (Megalobrama amblycephala) juveniles for chronic ammonia exposure: Antioxidant defense and the synthesis of urea and glutamine. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2020, 230, 108707. [Google Scholar] [CrossRef]
- Ni, H.; Peng, L.; Gao, X.; Ji, H.; Ma, J.Y.; Li, Y.; Jiang, S. Effects of medermycin on adult zebrafish (Danio rerio): Acute toxicity, tissue damage and oxidative stress. Ecotoxicol. Environ. Saf. 2019, 168, 249–259. [Google Scholar] [CrossRef]
- Gam, L.T.H.; Jensen, F.B.; Huong, D.T.T.; Phuong, N.T.; Bayley, M. The effects of elevated environmental CO2 on nitrite uptake in the air-breathing clown knifefish, Chitala ornata. Aquatic. Toxicol. 2018, 196, 124–131. [Google Scholar] [CrossRef]
- Kocour Kroupová, H.; Valentová, O.; Svobodová, Z.; Šauer, P.; Máchová, J. Toxic effects of nitrite on freshwater organisms: A review. Rev. Aquac. 2016, 10, 525–542. [Google Scholar] [CrossRef]
- Matsche, M.A.; Markin, E.L.; Donaldson, E.M.; Hengst, A.M.; Lazur, A. Effect of chloride on nitrite-induced methemoglobinemia in Atlantic sturgeon, Acipenser oxyrinchus (Mitchill). J. Fish Dis. 2012, 35, 873–885. [Google Scholar] [CrossRef]
- Jensen, F.B.; Koldkjær, P.; Bach, A. Anion uptake and acid-base and ionic effects during isolated and combined exposure to hypercapnia and nitrite in the freshwater crayfish, Astacus. J. Comp. Physiol. B 2000, 170, 489–495. [Google Scholar] [CrossRef]
- Gattinoni, L.; Pesenti, A.; Matthay, M.A. Understanding blood gas analysis. Intensive Care Med. 2017, 44, 91–93. [Google Scholar] [CrossRef] [PubMed]
- Hvas, M.; Damsgaard, C.D.; Gam, L.T.H.; Huong, D.T.T.; Jensen, F.B.; Bayley, M. The effect of environmental hypercapnia and size on nitrite toxicity in the striped catfish (Pangasianodon hypophthalmus). Aquat. Toxicol. 2016, 176, 151–160. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.Y.; Shieh, L.W.; Chen, J.C. Changes in hemolymph oxyhemocyanin, acid-base balance, and electrolytes in Marsupenaeus japonicus under combined ammonia and nitrite stress. Aquat. Toxicol. 2013, 130, 132–138. [Google Scholar] [CrossRef]
- Lin, Y.; Miao, L.H.; Pan, W.J.; Huang, X.; Dengu, J.M.; Zhang, W.X.; Ge, X.P.; Liu, B.; Ren, M.C.; Zhou, Q.L.; et al. Effect of nitrite exposure on the antioxidant enzymes and glutathione system in the liver of bighead carp, Aristichthys nobilis. Fish Shellfish Immunol. 2018, 76, 126–132. [Google Scholar] [CrossRef]
- Shen, C.; Cao, S.; Mohsen, M.; Li, X.S.; Wang, L.; Lu, K.L.; Zhang, C.X.; Song, K. Effects of chronic nitrite exposure on hematological parameters, oxidative stress and apoptosis in spotted seabass (Lateolabrax maculatus) reared at high temperature. Aquac. Rep. 2024, 35, 102022. [Google Scholar] [CrossRef]
- Guimaraes, D.A.; Portella, R.d.L.; Kamga, P.C.; Tanus-Santos, J.E.; Shiva, S.S. Nitrite Modulates Mitochondrial Function Through PKA Activation and AKAP121 Expression Under Normoxia. Free Radic. Biol. Med. 2015, 87, S77–S78. [Google Scholar] [CrossRef]
- Schopfer, F.J.; Riobo, N.A.; Carreras, M.C.; Alvarez, B.; Radi, R.; Boveris, A.; Cadenas, E.; Poderoso, J.J. Oxidation of ubiquinol by peroxynitrite: Implications for protection of mitochondria against nitrosative damage. Biochem. J. 2000, 349, 35–42. [Google Scholar] [CrossRef]
- Gupta, K.J.; Lee, C.P.; Lee, C.P.; Ratcliffe, R.G. Nitrite protects mitochondrial structure and function under hypoxia. Plant Cell Physiol. 2016, 58, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.Z.; Li, M.; Wang, R.X.; Qian, Y.X. Effects of acute ammonia toxicity on oxidative stress, immune response and apoptosis of juvenile yellow catfish Pelteobagrus fulvidraco and the mitigation of exogenous taurine. Fish Shellfish Immunol. 2018, 79, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Jia, R.; Han, C.; Lei, J.L.; Liu, B.L.; Huang, B.; Huo, H.H.; Yin, S.T. Effects of nitrite exposure on haematological parameters, oxidative stress and apoptosis in juvenile turbot (Scophthalmus maximus). Aquatic. Toxicol. 2015, 169, 1–9. [Google Scholar] [CrossRef]
- Pflaum, R.; Howick, L.C. Spectrophotometric Determination of Potassium with Sodium Tetraphenylborate. Anal. Chem. 1956, 28, 1542–1544. [Google Scholar] [CrossRef]
- Johnson, T.B.; Douglass, I.B. The action of chlorine on thiocyanates. J. Am. Chem. Soc. 1939, 61, 2548–2550. [Google Scholar] [CrossRef]
- Kumar, A.; Chapoteau, E.; Czech, B.; Gebauer, C.; Chimenti, M.; Raimondo, O. Chromogenic ionophore-based methods for spectrophotometric assay of sodium and potassium in serum and plasma. Clin. Chem. 1988, 34, 1709–1712. [Google Scholar] [CrossRef]
- Nägele, U.; Hägele, E.; Sauer, G.; Wiedemann, E.; Lehmann, P.; Wahlefeld, A.; Gruber, W. Reagent for the enzymatic determination of serum total triglycerides with improved lipolytic efficiency. Clin. Chem. Lab. Med. 1984, 22, 165–174. [Google Scholar] [CrossRef][Green Version]
- Janero, D.R. Malondialdehyde and thiobarbituric acid-reactivity as diagnostic indices of lipid peroxidation and peroxidative tissue injury. Free Radic. Biol. Med. 1990, 9, 515–540. [Google Scholar] [CrossRef]
- Peskin, A.V.; Winterbourn, C.C. A microtiter plate assay for superoxide dismutase using a water-soluble tetrazolium salt (WST-1). Clin. Chim. Acta 2000, 293, 157–166. [Google Scholar] [CrossRef]
- Aebi, H. [13] Catalase in vitro. Meth. Enzymol. 1984, 105, 121–126. [Google Scholar] [CrossRef]
- Nørby, J.G. [11] Coupled assay of Na+-K+-ATPase activity. Meth. Enzymol. 1988, 156, 116–119. [Google Scholar] [CrossRef]
- Chen, Z.; Sun, C. Contect of ATP in RBC detect by CPK. J. Pract. Med. Tech. 2002, 12, 908–909. Available online: https://kns.cnki.net/kcms2/article/abstract?v=EKYfHJ8l29jp96NxBk2yeai3-uQbS3ebhgfLeaD1yxXHUmGV7ZA3xQeOyJ90sXLP5goepa4Bkvm27jRBw_zAA6a_yTiuXxFgf68c0sTnz-64aZ6DEZk1sFQrfwF1FUwd9hobG3LSCqvi5oCxXmOFsSDSbTGS_eZjotWuPb39I4Yt0qpgSnXTzJApY7aMXGB1&uniplatform=NZKPT&language=CHS (accessed on 14 April 2025).
- Chan, K.M.; Delfert, D.; Junger, K. A direct colorimetric assay for Ca2+-stimulated ATPase activity. Anal. Biochem. 1986, 157, 375–380. [Google Scholar] [CrossRef]
- Narayana, B.; Sunil, K. A spectrophotometric method for the determination of nitrite and nitrate. Eur. J. Ageing Cancer 2009, 4, 204–214. [Google Scholar]
- Wind, T.C.; Messenger, M.P.; Thompson, D.; Selby, P.J.; Banks, R.E. Measuring carbonic anhydrase IX as a hypoxia biomarker: Differences in concentrations in serum and plasma using a commercial enzyme-linked immunosorbent assay due to influences of metal ions. Ann. Clin. Biochem. 2011, 48, 112–120. [Google Scholar] [CrossRef]
- Rahman, I.; Kode, A.; Biswas, S.K. Assay for quantitative determination of glutathione and glutathione disulfide levels using enzymatic recycling method. Nat. Protoc. 2006, 1, 3159–3165. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]










| Group | Time | pH | Dissolved Oxygen (mg/L) | Temperature (°C) |
|---|---|---|---|---|
| Control | 1–14th day | 7.85 ± 0.03 | 7.93 ± 0.11 | 25.55 ± 0.19 |
| Nitrite | 1–14th day | 7.79 ± 0.04 | 7.93 ± 0.06 | 25.58 ± 0.19 |
| Nitrite-2.5 CO2 | 1–14th day | 7.62 ± 0.03 | 7.79 ± 0.10 | 25.50 ± 0.12 |
| Nitrite-5 CO2 | 1–14th day | 7.43 ± 0.05 | 7.68 ± 0.12 | 25.50 ± 0.18 |
| Nitrite-10 CO2 | 1–14th day | 7.02 ± 0.04 | 7.60 ± 0.18 | 25.40 ± 0.08 |
| Control | 15–28th day | 7.82 ± 0.02 | 8.06 ± 0.05 | 25.51 ± 0.19 |
| Nitrite | 15–28th day | 7.78 ± 0.02 | 8.02 ± 0.03 | 25.35 ± 0.13 |
| Nitrite-2.5 CO2 | 15–28th day | 7.61 ± 0.04 | 7.94 ± 0.06 | 25.40 ± 0.14 |
| Nitrite-5 CO2 | 15–28th day | 7.44 ± 0.04 | 7.83 ± 0.07 | 25.43 ± 0.10 |
| Nitrite-10 CO2 | 15–28th day | 6.99 ± 0.09 | 7.69 ± 0.07 | 25.50 ± 0.12 |
| Group | Time | pH | P CO2 mmHg | P O2 mmHg | c(HCO3−) (mmol/L) | TCO2 (mmol/L) | sO2 (%) | Glu (mg/dL) |
|---|---|---|---|---|---|---|---|---|
| Control (0–0.1 mg/L Nitrite, 0–1 mg/L CO2) | 14th day | 6.84 ± 0.14 a | 11.67 ± 0.90 a | 28.33 ± 2.52 a | 1.87 ± 0.12 a | 7.13 ± 0.35 a | 47.67 ± 6.66 d | 22.00 ± 1.00 a |
| Nitrite (2 mg/L Nitrite, 0–1 mg/L CO2) | 6.96 ± 0.07 ab | 12.67 ± 0.21 ab | 29.67 ± 2.52 ab | 2.8 ± 0.17 b | 8.27 ± 0.15 b | 27.33 ± 2.08 a | 45.33 ± 2.08 d | |
| Nitrite (2 mg/L)-CO2 (2.5 mg/L) | 7.01 ± 0.01 b | 13.87 ± 0.64 bc | 32.67 ± 2.52 bc | 3.77 ± 0.40 c | 9.23 ± 0.31 c | 33.33 ± 2.08 ab | 35.00 ± 2.65 c | |
| Nitrite (2 mg/L)-CO2 (5 mg/L) | 7.07 ± 0.02 bc | 14.57 ± 0.78 cd | 35.00 ± 1.00 cd | 4.83 ± 0.25 d | 9.90 ± 0.36 c | 37.33 ± 1.53 bc | 34.33 ± 1.53 c | |
| Nitrite (2 mg/L)-CO2 (10 mg/L) | 7.17 ± 0.04 c | 15.67 ± 0.64 d | 37.33 ± 1.53 d | 5.90 ± 0.36 e | 12.2 ± 0.60 d | 43.00 ± 3.61 cd | 26.67 ± 2.08 b | |
| Control (0–0.1 mg/L Nitrite, 0–1 mg/L CO2) | 28th day | 6.83 ± 0.06 a | 12.27 ± 1.77 a | 28.33 ± 6.51 a | 1.93 ± 0.15 a | 8.27 ± 0.40 a | 49.00 ± 4.00 c | 26.33 ± 4.16 a |
| Nitrite (2 mg/L Nitrite, 0–1 mg/L CO2) | 6.95 ± 0.05 b | 13.2 ± 1.56 ab | 30.33 ± 4.16 ab | 2.73 ± 0.32 a | 9.63 ± 0.15 b | 32.67 ± 3.06 a | 48.67 ± 2.52 c | |
| Nitrite (2 mg/L)-CO2 (2.5 mg/L) | 6.97 ± 0.08 b | 15.27 ± 0.32 bc | 35.67 ± 1.15 bc | 4.30 ± 0.85 b | 9.87 ± 0.55 b | 40.67 ± 5.03 b | 33.33 ± 3.79 b | |
| Nitrite (2 mg/L)-CO2 (5 mg/L) | 7.11 ± 0.01 b | 16.07 ± 0.96 c | 38.00 ± 0 c | 5.67 ± 0.23 c | 12.27 ± 0.59 c | 45.33 ± 1.15 bc | 34.67 ± 1.53 b | |
| Nitrite (2 mg/L)-CO2 (10 mg/L) | 7.16 ± 0.03 b | 17.33 ± 0.87 c | 40.33 ± 2.31 c | 6.10 ± 0.70 c | 13.5 ± 0.26 d | 45.00 ± 2.65 bc | 31.00 ± 2.65 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Tang, Y.; Yang, H.; He, Y.; Ou-Yang, K.; Wang, L.; Zhang, Q.; Li, D.; Li, L. Increased CO2 Concentration Mitigates the Impact of Nitrite on Zebrafish (Danio rerio) Liver and Gills. Fishes 2025, 10, 205. https://doi.org/10.3390/fishes10050205
Wang X, Tang Y, Yang H, He Y, Ou-Yang K, Wang L, Zhang Q, Li D, Li L. Increased CO2 Concentration Mitigates the Impact of Nitrite on Zebrafish (Danio rerio) Liver and Gills. Fishes. 2025; 10(5):205. https://doi.org/10.3390/fishes10050205
Chicago/Turabian StyleWang, Xinyu, Yao Tang, Hui Yang, Ya He, Kang Ou-Yang, Liangmou Wang, Qian Zhang, Dapeng Li, and Li Li. 2025. "Increased CO2 Concentration Mitigates the Impact of Nitrite on Zebrafish (Danio rerio) Liver and Gills" Fishes 10, no. 5: 205. https://doi.org/10.3390/fishes10050205
APA StyleWang, X., Tang, Y., Yang, H., He, Y., Ou-Yang, K., Wang, L., Zhang, Q., Li, D., & Li, L. (2025). Increased CO2 Concentration Mitigates the Impact of Nitrite on Zebrafish (Danio rerio) Liver and Gills. Fishes, 10(5), 205. https://doi.org/10.3390/fishes10050205