
Dietary Supplementation of an Organic Acid-Based Feed Attractant in Juvenile Largemouth Bass (Micropterus salmoides): Effects on Growth, Morphohistology, and Oxidative Stress

 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Ethics
2.2. Diet Preparation
2.3. Fish Rearing
2.4. Sample Collection
2.5. Performance and Nutritional Content Measurement
2.6. Analysis of Biochemical Indices
2.7. Analysis of Antioxidant Capacity
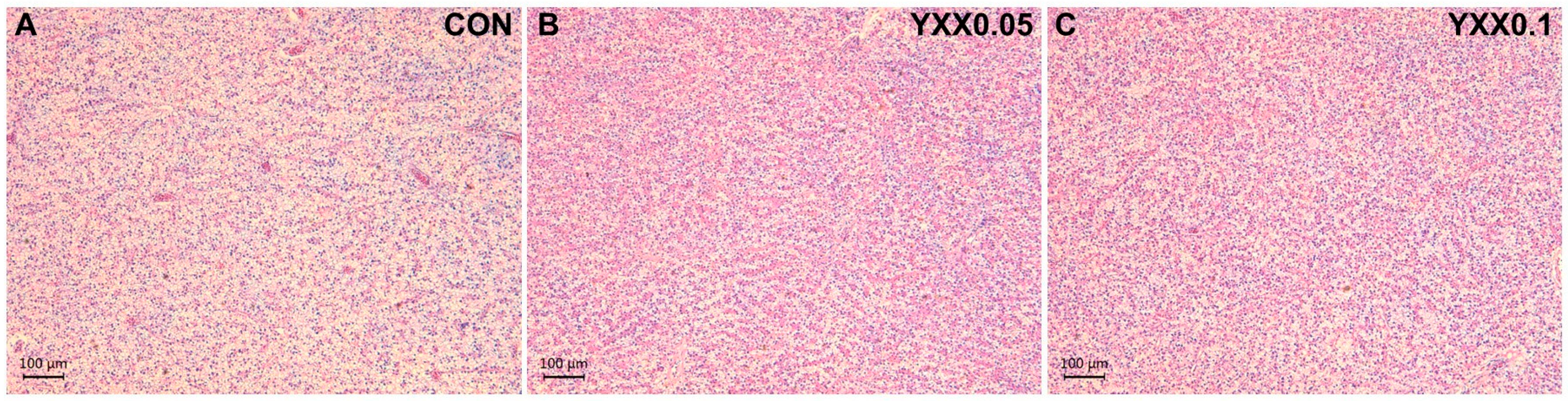
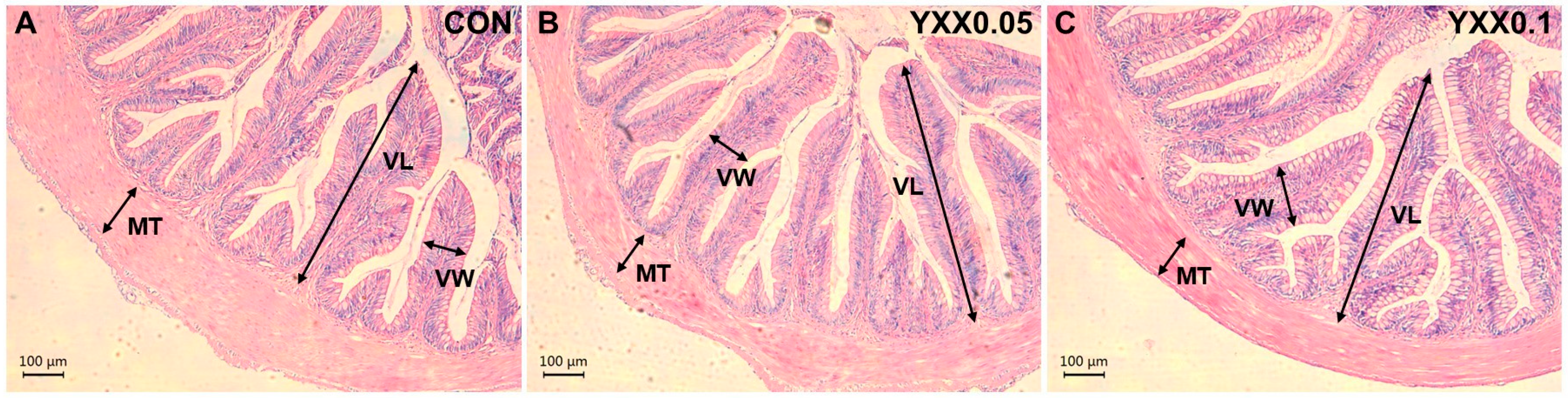
2.8. Histopathological Analysis of Liver and Intestine
2.9. Statistical Analysis
3. Results
3.1. The Effects of Fishy Attractant on Growth Performance and Organ Indexes
3.2. The Effects of Fish Attractant on Serum Biochemistry and Tissue Morphology
3.3. The Effects of Fish Attractant on Oxidative Stress in the Liver
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Carr, W.E. The molecular nature of chemical stimuli in the aquatic environment. In Sensory Biology of Aquatic Animals; Springer: Berlin/Heidelberg, Germany, 1988; pp. 3–27. [Google Scholar]
- Abdel-Daim, M.M.; Abdelkhalek, N.K.; Hassan, A.M. Antagonistic activity of dietary allicin against deltamethrin-induced oxidative damage in freshwater Nile tilapia; Oreochromis niloticus. Ecotoxicol. Environ. Saf. 2015, 111, 146–152. [Google Scholar] [CrossRef] [PubMed]
- Ajiboye, O.; Yakubu, A.; Simpa, J.; Balogun, S. Effect of garlic-supplemented diets on growth response, survival, nutrient utilization and body composition of monosex Tilapia zillii. World J. Fish Mar. Sci. 2016, 8, 115–122. [Google Scholar]
- Ismail, T.; Hegazi, E.; Dawood, M.A.; Nassef, E.; Bakr, A.; Paray, B.A.; Van Doan, H. Using of betaine to replace fish meal with soybean or/and corn gluten meal in nile tilapia (Oreochromis niloticus) diets: Histomorphology, growth, fatty acid, and glucose-related gene expression traits. Aquac. Rep. 2020, 17, 100376. [Google Scholar] [CrossRef]
- Sun, H.; Jiang, W.-D.; Wu, P.; Liu, Y.; Jiang, J.; Yang, Q.-H.; Kuang, S.-Y.; Tang, L.; Zhou, X.-Q.; Feng, L. Betaine supplementations enhance the intestinal immunity of on-growing grass carp (Ctenopharyngodon idella): Partly related to TOR and NF-κB signaling pathways. Aquaculture 2020, 518, 734846. [Google Scholar] [CrossRef]
- JP, D. Mixture interactions of glutamate and betaine in single squid olfactory receptor neurons. J. Comp. Physiol. 2000, 186, 57–67. [Google Scholar]
- Aaqillah-Amr, M.; Hidir, A.; Azra, M.; Ahmad-Ideris, A.; Abualreesh, M.; Noordiyana, M.; Ikhwanuddin, M. Use of pelleted diets in commercially farmed decapods during juvenile stages: A review. Animals 2021, 11, 1761. [Google Scholar] [CrossRef]
- Hameed, A.; Majeed, W.; Naveed, M.; Ramzan, U.; Bordiga, M.; Hameed, M.; Ur Rehman, S.; Rana, N. Success of aquaculture industry with new insights of using insects as feed: A review. Fishes 2022, 7, 395. [Google Scholar] [CrossRef]
- Öz, M.; Inanan, B.E.; Karasahin, T.; Dikel, S. Effects of glutamine on growth performance, nutrient content, fatty acid profile, and blood parameters of rainbow trout (Oncorhynchus mykiss). J. Fish Biol. 2024, 104, 1213–1222. [Google Scholar] [CrossRef]
- Ramos, A.R.P.; Campelo, D.A.V.; da Silva Carneiro, C.L.; Zuanon, J.A.S.; da Matta, S.L.P.; Furuya, W.M.; Salaro, A.L. Optimal dietary L-glutamine level improves growth performance and intestinal histomorphometry of juvenile giant trahira (Hoplias lacerdae), a Neotropical carnivorous fish species. Aquaculture 2022, 547, 737469. [Google Scholar] [CrossRef]
- Morais, S. The physiology of taste in fish: Potential implications for feeding stimulation and gut chemical sensing. Rev. Fish. Sci. Aquac. 2017, 25, 133–149. [Google Scholar] [CrossRef]
- Chowdhury, M.A.K.; Song, H.; Liu, Y.; Bunod, J.-D.; Dong, X.-H. Effects of microencapsulated organic acid and their salts on growth performance, immunity, and disease resistance of Pacific White Shrimp Litopenaeus vannamei. Sustainability 2021, 13, 7791. [Google Scholar] [CrossRef]
- Nunes, A.J.; Sá, M.V.; Andriola-Neto, F.F.; Lemos, D. Behavioral response to selected feed attractants and stimulants in Pacific white shrimp, Litopenaeus vannamei. Aquaculture 2006, 260, 244–254. [Google Scholar] [CrossRef]
- Alam, S.; Afzal, G.; Siddique, A.B.; Afzal, M.; Shahid, M.; Ramzan, A.; Iqbal, Z.; Ali, H.M.; Ahsan, H.; Hussain, R. Essential Oil-Based Functional Feeds for Promoting Growth in Aquaculture Species. Complement. Altern. Med. Essent. Oils 2024, 199–206. [Google Scholar] [CrossRef]
- Onomu, A.J.; Okuthe, G.E. The role of functional feed additives in enhancing aquaculture sustainability. Fishes 2024, 9, 167. [Google Scholar] [CrossRef]
- Dawood, M.A.; El Basuini, M.F.; Yilmaz, S.; Abdel-Latif, H.M.; Alagawany, M.; Kari, Z.A.; Abdul Razab, M.K.A.; Hamid, N.K.A.; Moonmanee, T.; Van Doan, H. Exploring the roles of dietary herbal essential oils in aquaculture: A review. Animals 2022, 12, 823. [Google Scholar] [CrossRef]
- Wang, J.; Deng, L.; Chen, M.; Che, Y.; Li, L.; Zhu, L.; Chen, G.; Feng, T. Phytogenic feed additives as natural antibiotic alternatives in animal health and production: A review of the literature of the last decade. Anim. Nutr. 2024, 17, 244–264. [Google Scholar] [CrossRef]
- Qu, K.; He, G.; Shi, M.; Chen, X.; Zhu, W.; Chen, Z.; Tan, B.; Xie, S. Effects of compound feed attractants on the growth rate, feed consumption, intestinal histology, protein synthesis, and immune response of black tiger shrimp (Penaeus monodon). Anim. Feed Sci. Technol. 2024, 311, 115952. [Google Scholar] [CrossRef]
- Kawamura, G.; Yong, A.S.-K.; Au, H.-L.; Doison, A.; Ooi, S.-Y.; Lim, L.-S.L.-S. Malaysian herbs as feeding attractants and enhancers for the giant freshwater prawn (Macrobrachium rosenbergii) and the whiteleg shrimp (Litopenaeus vannamei). Borneo J. Mar. Sci. Aquac. (BJoMSA) 2019, 3, 57–67. [Google Scholar] [CrossRef]
- Fry, J.P.; Mailloux, N.A.; Love, D.C.; Milli, M.C.; Cao, L. Feed conversion efficiency in aquaculture: Do we measure it correctly? Environ. Res. Lett. 2018, 13, 024017. [Google Scholar] [CrossRef]
- Van Rijn, J. Waste treatment in recirculating aquaculture systems. Aquac. Eng. 2013, 53, 49–56. [Google Scholar] [CrossRef]
- Dawood, M.A.; Koshio, S.; Esteban, M.Á. Beneficial roles of feed additives as immunostimulants in aquaculture: A review. Rev. Aquac. 2018, 10, 950–974. [Google Scholar] [CrossRef]
- Barad, R.R.; Verma, D.K.; Yusufzai, S.; Shrivastava, V.; Ram, A.R. Herbal Feed Additives: Natural Boost for Aquatic Health and Growth. In Sustainable Feed Ingredients and Additives for Aquaculture Farming: Perspectives from Africa and Asia; Springer: Berlin/Heidelberg, Germany, 2024; pp. 405–431. [Google Scholar]
- He, R.; Lei, B.; Su, Y.; Wang, A.; Cui, K.; Shi, X.; Chen, X. Effectiveness of eugenol as an anesthetic for adult spotted sea bass (Lateolabrax maculatus). Aquaculture 2020, 523, 735180. [Google Scholar] [CrossRef]
- Feldsine, P.; Abeyta, C.; Andrews, W.H. AOAC International methods committee guidelines for validation of qualitative and quantitative food microbiological official methods of analysis. J. AOAC Int. 2002, 85, 1187–1200. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.; Liu, Y.; Qiu, D.; Yi, M.; Li, X.; Li, Y. Effects of feeding rate and frequency on growth performance, digestion and nutrients balances of A tlantic salmon (Salmo salar) in recirculating aquaculture systems (RAS). Aquac. Res. 2016, 47, 176–188. [Google Scholar] [CrossRef]
- Ahmadifar, E.; Pourmohammadi Fallah, H.; Yousefi, M.; Dawood, M.A.; Hoseinifar, S.H.; Adineh, H.; Yilmaz, S.; Paolucci, M.; Doan, H.V. The gene regulatory roles of herbal extracts on the growth, immune system, and reproduction of fish. Animals 2021, 11, 2167. [Google Scholar] [CrossRef]
- Faheem, M.; Rao, Z.A.; Liaqat, I.; Hoseinifar, S.H.; Maneepitaksanti, W.; Van Doan, H. Bio-active components in medicinal plants: A mechanistic review of their effects on fish growth and physiological parameters–A review. Ann. Anim. Sci. 2022, 22, 1127–1149. [Google Scholar] [CrossRef]
- Abwao, J.; Jung’a, J.; Barasa, J.E.; Kyule, D.; Opiyo, M.; Awuor, J.F.; Ogello, E.; Munguti, J.M.; Keya, G.A. Selective breeding of Nile tilapia, Oreochromis niloticus: A strategy for increased genetic diversity and sustainable development of aquaculture in Kenya. J. Appl. Aquac. 2023, 35, 237–256. [Google Scholar] [CrossRef]
- Salama, A.J. Effects of different feeding frequency on the growth, survival and feed conversion ratio of the Asian sea bass Lates calcarifer juveniles reared under hypersaline seawater of the Red Sea. Aquac. Res. 2008, 39, 561–567. [Google Scholar] [CrossRef]
- Yue, Y.; Chen, M.; Bao, X.; Yu, Y.; Shi, W.; Kumkhong, S.; Liu, Y.; Yang, Y.; Yu, H. Effects of three feed attractants on the growth performance and meat quality of the largemouth bass (Micropterus salmoides). Front. Mar. Sci. 2022, 9, 1029969. [Google Scholar] [CrossRef]
- Peng, K.; Huang, W.; Zhao, H.; Sun, Y.; Chen, B. Dietary condensed tannins improved growth performance and antioxidant function but impaired intestinal morphology of Litopenaeus vannamei. Aquac. Rep. 2021, 21, 100853. [Google Scholar] [CrossRef]
- Abdel-Latif, H.M.; Abdel-Tawwab, M.; Dawood, M.A.; Menanteau-Ledouble, S.; El-Matbouli, M. Benefits of dietary butyric acid, sodium butyrate, and their protected forms in aquafeeds: A review. Rev. Fish. Sci. Aquac. 2020, 28, 421–448. [Google Scholar] [CrossRef]
- Feng, H.; Peng, D.; Liang, X.-F.; Li, J.; Luo, H.; Tang, S.; Chai, F. Intracerebroventricular injection with octanoic acid activates hypothalamic fatty acid sensing systems and regulates appetite in Chinese perch Siniperca chuatsi. Fish. Sci. 2022, 88, 83–90. [Google Scholar] [CrossRef]
- Jensen, M.M.; Christensen, M.S.; Høy, C.-E. Intestinal absorption of octanoic, decanoic, and linoleic acids: Effect of triglyceride structure. Ann. Nutr. Metab. 1994, 38, 104–116. [Google Scholar] [CrossRef] [PubMed]
- Bhuiyan, M.S.; Ellett, F.; Murray, G.L.; Kostoulias, X.; Cerqueira, G.M.; Schulze, K.E.; Mahamad Maifiah, M.H.; Li, J.; Creek, D.J.; Lieschke, G.J. Acinetobacter baumannii phenylacetic acid metabolism influences infection outcome through a direct effect on neutrophil chemotaxis. Proc. Natl. Acad. Sci. USA 2016, 113, 9599–9604. [Google Scholar] [CrossRef]
- Ng, W.K.; Koh, C.B. The utilization and mode of action of organic acids in the feeds of cultured aquatic animals. Rev. Aquac. 2017, 9, 342–368. [Google Scholar] [CrossRef]
- Rasidi, R.; Pamungkas, W.; Handajani, H.; Puspaningsih, D.; Taqwa, F.H.; Hartami, P. Sustaining Aquaculture: Organic Acid as Feed Additives in Aquaculture. In Sustainable Feed Ingredients and Additives for Aquaculture Farming: Perspectives from Africa and Asia; Springer: Berlin/Heidelberg, Germany, 2024; pp. 481–500. [Google Scholar]
- Hancz, C. Feed efficiency, nutrient sensing and feeding stimulation in aquaculture: A review. Acta Agrar. Kaposváriensis 2020, 24, 35–54. [Google Scholar] [CrossRef]
- Lin, X. Optimizing Animal Energy Metabolism: Nutritional Strategies in Agricultural Production. Int. J. Mol. Zool. 2025, 14. [Google Scholar] [CrossRef]
- Api, A.; Belmonte, F.; Belsito, D.; Botelho, D.; Bruze, M.; Burton, G., Jr.; Buschmann, J.; Dagli, M.; Date, M.; Dekant, W. RIFM fragrance ingredient safety assessment, isovaleric acid, CAS Registry Number 503-74-2. Food Chem Toxicol. 2019, 130, 110570. [Google Scholar] [CrossRef]
- de Figueirêdo Urach, B.; de Souza Ramos, A.P.; Luz, J.R.; de Magalhães Júnior, F.O.; Schorer, M.; Braga, L.G.T. Sodium butyrate improves the performance of juvenile tilapia, Oreochromis niloticus (Linneaus, 1758). Res. Soc. Dev. 2020, 9, e551997535. [Google Scholar] [CrossRef]
- Meng, X.; Cai, H.; Li, H.; You, F.; Jiang, A.; Hu, W.; Li, K.; Zhang, X.; Zhang, Y.; Chang, X. Clostridium butyricum-fermented Chinese herbal medicine enhances the immunity by modulating the intestinal microflora of largemouth bass (Micropterus salmoides). Aquaculture 2023, 562, 738768. [Google Scholar] [CrossRef]
- Ragland, S.A.; Criss, A.K. From bacterial killing to immune modulation: Recent insights into the functions of lysozyme. PLoS Pathog. 2017, 13, e1006512. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Wei, C.; Li, Y.; Wang, M.; Mao, Y.; Tian, X. A Comparative Study on Effects of Three Butyric Acid-Producing Additives on the Growth Performance, Non-specific Immunity, and Intestinal Microbiota of the Sea Cucumber Apostichopus japonicus. Aquac. Nutr. 2024, 2024, 6973951. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Shu, H.; Kou, Y.; Dang, H.; Ai, C. Beneficial effects of the butanoic acid derivative tributyrin on the growth, immunity and intestinal health of large mouth bass (Micropterus salmoides). Aquaculture 2024, 590, 741007. [Google Scholar] [CrossRef]
- Kalas, M.A.; Chavez, L.; Leon, M.; Taweesedt, P.T.; Surani, S. Abnormal liver enzymes: A review for clinicians. World J. Hepatol. 2021, 13, 1688. [Google Scholar] [CrossRef]
- Peng, D.; Peng, B.; Li, J.; Zhang, Y.; Luo, H.; Xiao, Q.; Tang, S.; Liang, X.-F. Effects of three feed attractants on the growth, biochemical indicators, lipid metabolism and appetite of Chinese perch (Siniperca chuatsi). Aquac. Rep. 2022, 23, 101075. [Google Scholar] [CrossRef]
- Deepa, K.; Purushothaman, M.; Vasanthakumar, P.; Sivakumar, K. Butyric acid as an antibiotic substitute for broiler chicken–A review. Adv. Anim. Vet. Sci. 2018, 6, 63–69. [Google Scholar] [CrossRef]
- Abdeldayem, F.A.; Lestingi, A.; Abol-Ela, S.S.; Alagawany, M.; Ismail, T.A.; Mostafa, N.G.; El-Shall, N.A. Application of butyric acid as a feed additive for improving quail performance and health. Poult. Sci. 2024, 103, 104109. [Google Scholar] [CrossRef]
- Yohana, M.A.; Ray, G.W.; Yang, Q.; Shiyu, K.; Tan, B.; Wu, J.; Mao, M.; Feng, L. Comprehensive analysis of butyric acid impact on immunology, histopathology, gene expression, and metabolomic responses in pacific shrimp experiencing cold stress. Comp. Biochem. Physiol. Part D Genom. Proteom. 2024, 52, 101293. [Google Scholar] [CrossRef]
- Li, L.; Gao, M.; Yang, N.; Ai, L.; Guo, L.; Xue, X.; Sheng, Z. Trimethyltin chloride induces apoptosis and DNA damage via ROS/NF-κB in grass carp liver cells causing immune dysfunction. Fish Shellfish Immunol. 2023, 142, 109082. [Google Scholar] [CrossRef]
- Bian, D.-D.; Zhang, X.; Zhu, X.-R.; Tang, W.-H.; Peng, Q.; Chen, Y.-H.; Wang, G.; Zhang, D.-Z.; Tang, B.-P.; Liu, Q.-N. The Nrf2-Keap1/ARE signaling pathway in aquatic animals. Int. J. Biol. Macromol. 2025, 308, 142595. [Google Scholar] [CrossRef]
- Zhao, M.; Zhang, Z.; Liu, Y.; Zhang, W.; Gong, Y.; Tang, Y.; Chen, F.; Zhang, J.; Liu, G.; Zhang, H. Effects of supplemental octanoate on hepatic lipid metabolism, serum biochemical indexes, antioxidant capacity and inflammation-related genes expression of large yellow croaker (Larimichthys crocea) fed with high soybean oil diet. Front. Immunol. 2023, 14, 1162633. [Google Scholar] [CrossRef]
- Aalamifar, H.; Soltanian, S.; Vazirzadeh, A.; Akhlaghi, M.; Morshedi, V.; Gholamhosseini, A.; Torfi Mozanzadeh, M. Dietary butyric acid improved growth, digestive enzyme activities and humoral immune parameters in Barramundi (Lates calcarifer). Aquac. Nutr. 2020, 26, 156–164. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Item | Experimental Diets | ||
|---|---|---|---|
| CON | YXX0.05 | YXX0.1 | |
| BLU fish meal | 35.00 | 35.00 | 35.00 |
| Chicken meal | 15.00 | 15.00 | 15.00 |
| Corn gluten meal | 5.00 | 5.00 | 5.00 |
| Soy protein concentrate | 14.00 | 14.00 | 14.00 |
| Wheat starch | 10.00 | 10.00 | 10.00 |
| Soybean meal | 9.00 | 9.00 | 9.00 |
| Soybean oil | 3.00 | 3.00 | 3.00 |
| Fish oil | 3.00 | 3.00 | 3.00 |
| Soy lecithin oil | 1.00 | 1.00 | 1.00 |
| VC | 0.20 | 0.20 | 0.20 |
| Sea bass premix | 1.00 | 1.00 | 1.00 |
| Calcium dihydrogen phosphate | 2.00 | 2.00 | 2.00 |
| Sodium alginate | 1.00 | 1.00 | 1.00 |
| Choline chloride | 0.50 | 0.50 | 0.50 |
| Microcrystalline cellulose | 0.30 | 0.25 | 0.20 |
| Fishy attractant | 0.00 | 0.05 | 0.10 |
| Proximate compositions | |||
| Crude protein | 49.72 | 49.72 | 49.72 |
| Crude lipid | 12.86 | 12.86 | 12.86 |
| Index | Group | ||
|---|---|---|---|
| CON | YXX0.05 | YXX0.1 | |
| FBW (g/fish) | 81.66 ± 1.32 | 83.41 ± 1.02 | 83.91 ± 1.21 |
| FI (g/fish) | 62.18 ± 1.08 b | 65.34 ± 1.21 a | 65.68 ± 0.69 a |
| SGR (%/d) | 2.74 ± 0.03 | 2.77 ± 0.02 | 2.74 ± 0.03 |
| FCR | 0.94 ± 0.01 | 0.94 ± 0.01 | 0.95 ± 0.01 |
| WGR (%) | 416.23 ± 8.35 | 427.29 ± 6.55 | 417.64 ± 7.37 |
| HSI (%) | 1.59 ± 0.02 | 1.52 ± 0.04 | 1.51 ± 0.07 |
| VSI (%) | 9.95 ± 0.15 a | 9.59 ± 0.07 b | 9.59 ± 0.07 b |
| SR (%) | 98.67 ± 2.31 | 94.67 ± 2.31 | 97.33 ± 2.31 |
| CF (g/cm3) | 1.57 ± 0.07 b | 1.69 ± 0.01 a | 1.70 ± 0.03 a |
| Index | Group | ||
|---|---|---|---|
| CON | YXX0.05 | YXX0.1 | |
| Moisture | 67.65 ± 0.69 | 67.21 ± 0.72 | 67.28 ± 0.74 |
| Crude Fat | 6.38 ± 0.15 | 6.43 ± 0.16 | 6.39 ± 0.11 |
| Crude Protein | 18.75 ± 0.14 | 18.64 ± 0.07 | 18.79 ± 0.13 |
| Ash | 4.56 ± 0.49 | 4.52 ± 0.47 | 4.34 ± 0.44 |
| Index | Group | ||
|---|---|---|---|
| CON | YXX0.05 | YXX0.1 | |
| Villus length (μm) | 611.70 ± 116.10 | 599.60 ± 112.01 | 596.81 ± 76.09 |
| Villus width (μm) | 151.62 ± 32.75 | 137.87 ± 20.00 | 113.00 ± 20.19 |
| Muscular thickness (μm) | 134.94 ± 23.60 | 125.93 ± 32.31 | 124.79 ± 13.49 |
| Index | Group | ||
|---|---|---|---|
| CON | YXX0.05 | YXX0.1 | |
| AST (U/L) | 87.74 ± 3.42 a | 72.85 ± 3.42 b | 82.63 ± 9.07 b |
| ALT (U/L) | 8.95 ± 1.40 | 7.76 ± 0.45 | 6.40 ± 0.66 |
| LZM (U/mL) | 21.07 ± 1.66 b | 27.43 ± 1.73 a | 27.03 ± 1.01 a |
| AKP (U/mgprot) | 4.66 ± 0.57 | 5.04 ± 0.18 | 5.10 ± 0.64 |
| ACP (U/100mL) | 10.02 ± 0.27 | 10.92 ± 0.30 | 10.43 ± 1.43 |
| Index | Group | ||
|---|---|---|---|
| CON | YXX0.05 | YXX0.1 | |
| T-AOC (U/mg) | 1.04 ± 0.01 b | 1.15 ± 0.01 a | 1.14 ± 0.01 a |
| GSH-Px (U/g) | 2174 ± 29.04 | 2185.05 ± 23.48 | 2217.24 ± 78.71 |
| SOD (U/mg) | 19.81 ± 1.57 | 20.10 ± 0.40 | 21.91 ± 0.40 |
| MDA (nmol/mgprot) | 0.40 ± 0.01 a | 0.31 ± 0.01 b | 0.37 ± 0.01 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cai, G.; Li, Z.; Yu, M.; Huang, M.; Liu, P.; Tang, X.; Huang, Q.; Guo, Z.; Sun, Y. Dietary Supplementation of an Organic Acid-Based Feed Attractant in Juvenile Largemouth Bass (Micropterus salmoides): Effects on Growth, Morphohistology, and Oxidative Stress. Fishes 2025, 10, 195. https://doi.org/10.3390/fishes10050195
Cai G, Li Z, Yu M, Huang M, Liu P, Tang X, Huang Q, Guo Z, Sun Y. Dietary Supplementation of an Organic Acid-Based Feed Attractant in Juvenile Largemouth Bass (Micropterus salmoides): Effects on Growth, Morphohistology, and Oxidative Stress. Fishes. 2025; 10(5):195. https://doi.org/10.3390/fishes10050195
Chicago/Turabian StyleCai, Guohe, Zhizhong Li, Miao Yu, Mingya Huang, Pengao Liu, Xiudan Tang, Qizhe Huang, Zhixue Guo, and Yunzhang Sun. 2025. "Dietary Supplementation of an Organic Acid-Based Feed Attractant in Juvenile Largemouth Bass (Micropterus salmoides): Effects on Growth, Morphohistology, and Oxidative Stress" Fishes 10, no. 5: 195. https://doi.org/10.3390/fishes10050195
APA StyleCai, G., Li, Z., Yu, M., Huang, M., Liu, P., Tang, X., Huang, Q., Guo, Z., & Sun, Y. (2025). Dietary Supplementation of an Organic Acid-Based Feed Attractant in Juvenile Largemouth Bass (Micropterus salmoides): Effects on Growth, Morphohistology, and Oxidative Stress. Fishes, 10(5), 195. https://doi.org/10.3390/fishes10050195