
Effects of Stocking Densities on the Growth Performance and Physiology of Juvenile Piaractus brachypomus in Recirculating Aquaculture System

 , ,
, ,  , , ,
, , ,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Experiment I
2.2. Experiment II
2.3. Water Quality Analysis
2.4. Growth and Survival
- -
- Initial weight (g) (IW);
- -
- Initial total length (cm) (IL);
- -
- Final weight (g) (FW);
- -
- Final total length (cm) (FL);
- -
- Weight gain (g) (WG) = FW − IW;
- -
- Daily weight gain (g) (DWG) = (FW − IW)/ΔT, where ΔT is the duration of the experiment in days;
- -
- Specific growth rate (%/day) (SGR) = 100 (ln FW − ln IW)/ΔT, where ΔT is the duration of the experiment in days;
- -
- Final biomass (FB) (kg/m3), where all the weights of all the fish were added together and the value found was converted to kg/m3;
- -
- Feed consumption (FC) (kg) = (feed offered (g) − feed leftover (g))/100
- -
- Feed conversion rate (FCR) = consumption/(FB − IB), where IB is the initial biomass.
2.5. Hematological and Biochemical Analysis
2.6. Viscerosomatic and Hepatosomatic Indexes
- -
- Viscerosomatic Index (VSI) (%) = 100 × ((weight of total viscera (g)/body weight (g)).
- -
- Hepatosomatic Index (HSI) (%) = 100 × (weight of liver/body weight).
2.7. Statistical Analysis
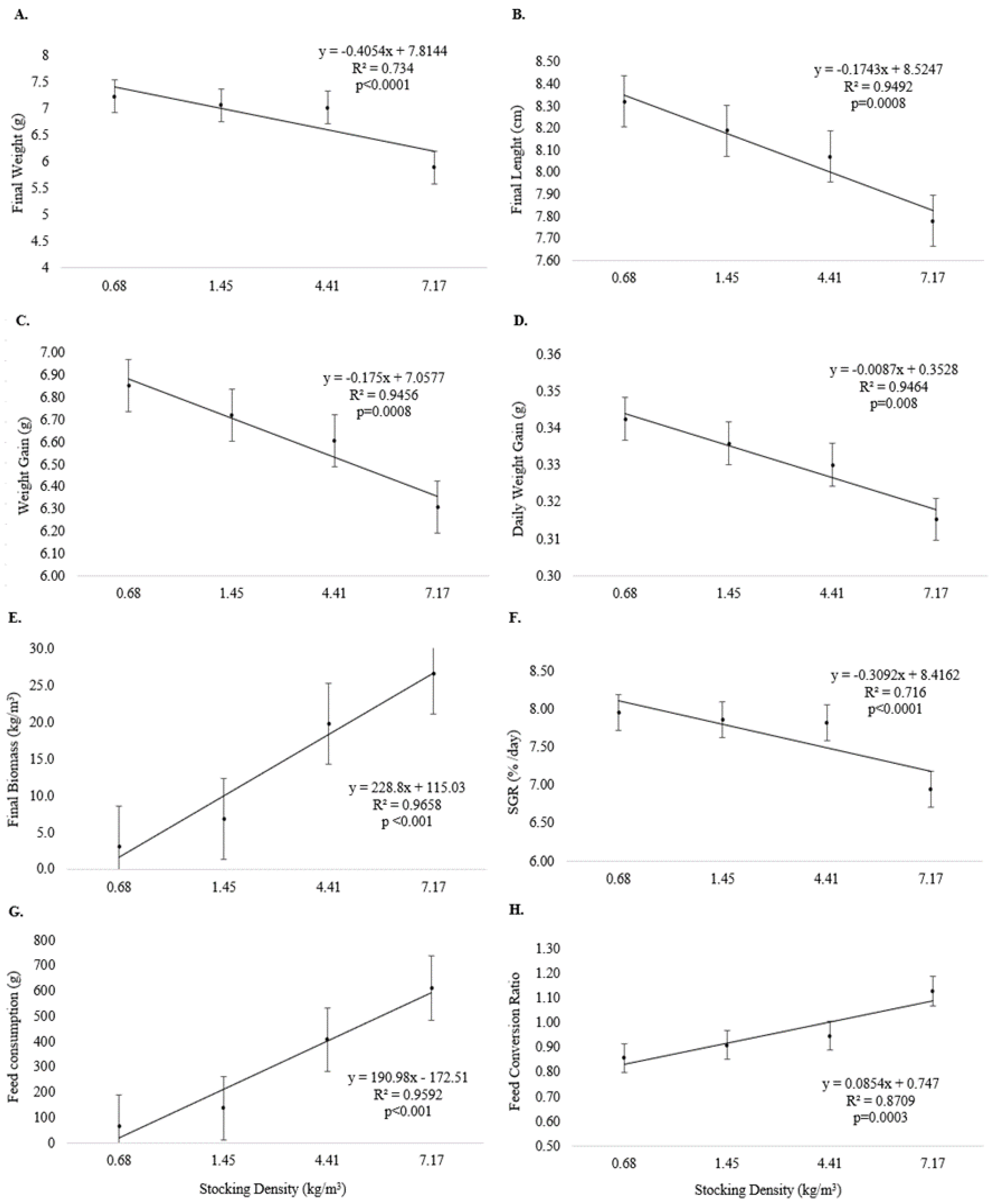
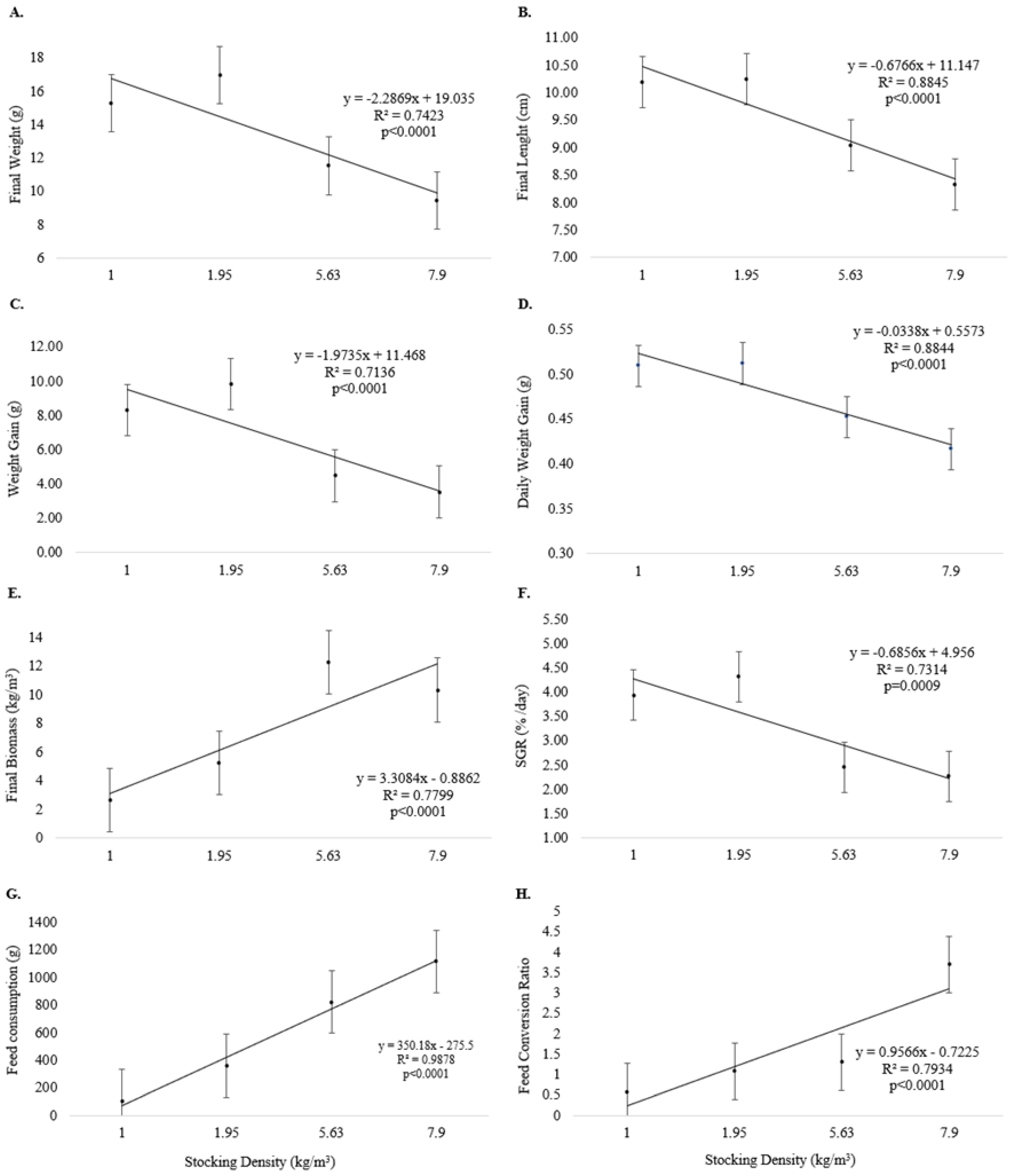
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ray, A. Biofloc technology for super-intensive shrimp culture. In Biofloc Technology—A Practical Guide Book, 2nd ed.; The World Aquaculture Society: Baton Rouge, LO, USA, 2012; Volume 167, p. 188. [Google Scholar]
- Chun, S.-J.; Cui, Y.; Ahn, C.-Y.; Oh, H.-M. Improving water quality using settleable microalga Ettlia sp. and the bacterial community in freshwater recirculating aquaculture system of Danio rerio. Water Res. 2018, 135, 112–121. [Google Scholar] [PubMed]
- Verdegem, M.C.J.; Bosma, R.H.; Verreth, J.A.J. Reducing water use for animal production through aquaculture. Int. J. Water Resour. Dev. 2006, 22, 101–113. [Google Scholar]
- Martins, C.I.M.; Eding, E.H.; Verdegem, M.C.J.; Heinsbroek, L.T.N.; Schneider, O.; Blancheton, J.P.; d’Orbcastel, E.R.; Verreth, J.A.J. New developments in recirculating aquaculture systems in Europe: A perspective on environmental sustainability. Aquac. Eng. 2010, 43, 83–93. [Google Scholar] [CrossRef]
- Sousa, R.M.; Silva, R.R.D.S.; Santos, A.S.D.; Silva, C.V.D.; Magalhães, J.A.; Fogaça, F.H.D.S.; Lopes, J.M. Tambatinga juveniles performance in a recirculation aquaculture system with different stocking densities. RSD 2020, 9, e178953317. [Google Scholar]
- Karnatak, G.; Das, B.K.; Mishal, P.; Tayung, T.; Kumari, S.; Sarkar, U.K.; Das, A.K.; Ali, Y. Impact of stocking density on growth, feed utilization and survival of cage reared minor carp, Labeo bata (Hamilton, 1822) in Maithon reservoir, India. Aquaculture 2021, 532, 736078. [Google Scholar]
- Clols-Fuentes, J.; Nguinkal, J.A.; Unger, P.; Kreikemeyer, B.; Palm, H.W. Bacterial community in African catfish (Clarias gariepinus) recirculating aquaculture systems under different stocking densities. Front. Mar. Sci. 2023, 10, 1073250. [Google Scholar]
- Yarahmadi, P.; Miandare, H.K.; Fayaz, S.; Caipang, C.M.A. Increased stocking density causes changes in expression of selected stress- and immune-related genes, humoral innate immune parameters and stress responses of rainbow trout (Oncorhynchus mykiss). Fish Shellfish Immunol. 2016, 48, 43–53. [Google Scholar]
- Wang, L.; Jia, S.; Guo, X.-R.; Lu, K.; Zhang, L.; Gong, J.; Guo, X.-P.; Hu, Y.; Cheng, T.; Shang, Q.; et al. Effect of stocking density on growth of largemouth bass (Micropterus salmoides) cultured in containers in a land-based recirculating aquaculture system (C-RAS). Aquac. Res. 2021, 53, 1518–1526. [Google Scholar]
- Bassmann, B.; Hahn, L.; Rebl, A.; Wenzel, L.C.; Hildebrand, M.-C.; Verleih, M.; Palm, H.W. Effects of stocking density, size, and external stress on growth and welfare of African catfish (Clarias gariepinus Burchell, 1822) in a commercial RAS. Fishes 2023, 8, 74. [Google Scholar] [CrossRef]
- Rebl, A.; Zebunke, M.; Borchel, A.; Bochert, R.; Verleih, M.; Goldammer, T. Microarray-predicted marker genes and molecular pathways indicating crowding stress in rainbow trout (Oncorhynchus mykiss). Aquaculture 2017, 473, 355–365. [Google Scholar]
- Costa, O.T.F.D.; Dias, L.C.; Malmann, C.S.Y.; Lima Ferreira, C.A.D.; Carmo, I.B.D.; Wischneski, A.G.; Sousa, R.L.D.; Cavero, B.A.S.; Lameiras, J.L.V.; Santos, M.C. The effects of stocking density on the hematology, plasma protein profile and immunoglobulin production of juvenile tambaqui (Colossoma macropomum) farmed in Brazil. Aquaculture 2019, 499, 260–268. [Google Scholar]
- Santos, F.A.C.; Boaventura, T.P.; Julio, G.S.C.; Cortezzi, P.P.; Figueiredo, L.G.; Favero, G.C.; Palheta, G.D.A.; Melo, N.F.A.C.; Luz, R.K. Growth performance and physiological parameters of Colossoma macropomum in a recirculating aquaculture system (RAS): Importance of stocking density and classification. Aquaculture 2021, 534, 736274. [Google Scholar]
- Ananias, I.D.M.C.; Silva, S.D.S.; Santos, F.A.C.D.; Souza, A.D.S.; Magalhães, T.B.; Reis, P.A.R.; Favero, G.C.; Luz, R.K. Tambaqui production at different stocking densities in RAS: Growth and physiology. Fishes 2024, 9, 19. [Google Scholar]
- Sánchez, P.; Ambrosio, P.P.; Flos, R. Stocking density affects Senegalese sole (Solea senegalensis, Kaup) growth independently of size dispersion, evaluated using an individual photo-identification technique. Aquac. Res. 2013, 44, 231–241. [Google Scholar]
- Ahmed, I.; Reshi, Q.M.; Fazio, F. The influence of the endogenous and exogenous factors on hematological parameters in different fish species: A review. Aquacult Int 2020, 28, 869–899. [Google Scholar]
- Ruane, N.M.; Komen, H. Measuring cortisol in the water as an indicator of stress caused by increased loading density in common carp (Cyprinus carpio). Aquaculture 2003, 218, 685–693. [Google Scholar]
- Khansari, A.R.; Balasch, J.C.; Vallejos-Vidal, E.; Teles, M.; Fierro-Castro, C.; Tort, L.; Reyes-López, F.E. Comparative study of stress and immune-related transcript outcomes triggered by Vibrio anguillarum bacterin and air exposure stress in liver and spleen of gilthead seabream (Sparus aurata), zebrafish (Danio rerio) and rainbow trout (Oncorhynchus mykiss). Fish Shellfish Immunol. 2019, 86, 436–448. [Google Scholar]
- Barton, B.A. Stress in fishes: A diversity of responses with particular reference to changes in circulating corticosteroids. Integr. Comp. Biol. 2002, 42, 517–525. [Google Scholar]
- Upadhyay, A.; Swain, H.S.; Das, B.K.; Ramtke, M.H.; Kumar, V.; Krishna, G.; Mohanty, B.P.; Chadha, N.K.; Das, A.K. Stocking density matters in open water cage culture: Influence on growth, digestive enzymes, haemato-immuno and stress responses of Puntius sarana (Ham, 1822). Aquaculture 2022, 30, 73745. [Google Scholar]
- Nakano, T.; Kameda, M.; Shoji, Y.; Hayashi, S.; Yamaguchi, T.; Sato, M. Effect of severe environmental thermal stress on redox state in salmon. Redox. Biol. 2014, 5, 772–776. [Google Scholar]
- Refaey, M.M.; Li, D.; Tian, X.; Zhang, Z.; Zhang, X.; Li, L.; Tang, R. High stocking density alters growth performance, blood biochemistry, intestinal histology, and muscle quality of Channel catfish Ictalurus punctatus. Aquaculture 2018, 492, 73–81. [Google Scholar] [CrossRef]
- Wang, Y.; Zhu, J.; Ge, X.; Sun, S.; Su, Y.; Li, B.; Hou, Y.; Ren, M. Effects of stocking density on the growth performance, digestive enzyme activities, antioxidant resistance, and intestinal microflora of Blunt Snout Bream (Megalobrama Amblycephala) juveniles. Aquac. Res. 2019, 50, 236–246. [Google Scholar] [CrossRef]
- Fatima, S.; Izhar, S.; Usman, Z.; Rashid, F.; Kanwal, Z.; Jabeen, G.; Latif, A.A. Effects of high stocking density on condition factor and profile of free thyroxine and cortisol in Catla catla (Hamilton, 1822) and Labeo rohita (Hamilton, 1822). Turk. J. Fish. Aquat. Sci. 2018, 18, 217–221. [Google Scholar]
- Escobar, L.M.D.; Ota, R.P.; Machado-Allison, A.; Andrade-López, J.; Farias, I.P.; Hrbek, T. A new species of Piaractus (Characiformes: Serrasalmidae) from the Orinoco Basin with a redescription of Piaractus brachypomus. J. Fish Biol. 2019, 95, 411–427. [Google Scholar] [CrossRef]
- Kolmann, M.A.; Kalacska, M.; Lucanus, O.; Sousa, L.; Wainwright, D.; Arroyo-Mora, J.P.; Andrade, M.C. Hyperspectral data as a biodiversity screening tool can differentiate among diverse. Sci. Rep. 2021, 11, 16157. [Google Scholar] [CrossRef]
- Kumar, A.; Pradhan, P.K.; Das, P.C.; Srivastava, S.M.; Lal, K.K.; Jena, J.K. Growth performance and compatibility of pacu, Piaractus brachypomus with Indian major carps in polyculture system. Aquaculture 2018, 490, 236–239. [Google Scholar] [CrossRef]
- Bharane, P.R.; Bethi, C.M.S.; Kudre, T.G. Effect of Catla catla roe protein isolate on textural and sensorial properties of surimi gel from Piaractus brachypomus. Food Meas. 2020, 14, 1391–1401. [Google Scholar] [CrossRef]
- Theerawoot, L. Diversity and distribution of external parasites from potentially cultured freshwater fishes in Nakhonsithammarat, Southern Thailand. Dis. Asian Aquac. 2008, 6, 235–244. [Google Scholar]
- Xiong, W.; Sui, X.; Liang, S.-H.; Chen, Y. Non-native freshwater fish species in China. Rev. Fish Biol. Fish. 2015, 25, 651–687. [Google Scholar] [CrossRef]
- Garcia, L.D.O.; Gutiérrez-Espinosa, M.; Wásquez-Torres, W.; Baldisserotto, B. Dietary protein levels in Piaractus brachypomus submitted to extremely acidic or alkaline pH. Cienc. Rural 2014, 44, 301–306. [Google Scholar] [CrossRef]
- Favero, G.C.; Santos, F.A.C.; Júlio, G.S.C.; Pedras, P.P.C.; Ferreira, A.L.; Silva, W.S.; Ferreira, N.S.; Neves, L.C.; Luz, R.K. Effects of short feed restriction cycles in Piaractus brachypomus juveniles. Aquaculture 2021, 536, 736465. [Google Scholar] [CrossRef]
- Ferreira, A.L.; Bonifácio, C.T.; Silva, W.S.; Takata, R.; Favero, G.C.; Luz, R.K. Anesthesia with eugenol and menthol for Piaractus brachypomus (Cuvier, 1818): Induction and recovery times, ventilation frequency and hematological and biochemical responses. Aquaculture 2021, 544, 737076. [Google Scholar] [CrossRef]
- Favero, G.C.; Santos, F.A.C.; Júlio, G.S.C.; Batista, F.S.; Bonifácio, C.T.; Torres, I.F.A.; Paranhos, C.O.; Luz, R.K. Effects of water temperature and feeding time on growth performance and physiological parameters of Piaractus brachypomus juveniles. Aquaculture 2022, 548, 737716. [Google Scholar] [CrossRef]
- Corrêa, R.O. Manejo alimentar. In Criação de Tambaqui; Correa, R.O., Sousa, A.R.B., Martins-Junior, H., Eds.; Embrapa: Brasília, Brazil, 2018; pp. 13–19. [Google Scholar]
- Santos, F.A.C.; Costa Julio, G.S.; Luz, R.K. Stocking density in Colossoma macropomum larviculture, a freshwater fish, in recirculating aquaculture system. Aquac. Res. 2021, 52, 1185–1191. [Google Scholar] [CrossRef]
- Goldenfarb, P.B.; Bowyer, F.P.; Hall, E.; Brosious, E. Reproducibility in the hematology laboratory: The microhematocrit determination. Am. J. Clin. Pathol. 1971, 56, 35–39. [Google Scholar] [CrossRef]
- Wintrobe, M.M. Variations in the size and hemoglobin content of erythrocytes in the blood of various vertebrates. Folia Haematol. 1934, 51, 32–49. [Google Scholar]
- Mattioli, C.C.; Takata, R.; Leme, F.O.P.; Costa, D.C.; Melillo Filho, R.; Silva, W.S.; Luz, R.K. The effects of acute and chronic exposure to water salinity on juveniles of the carnivorous freshwater catfish Lophiosilurus alexandri. Aquaculture 2017, 481, 255–266. [Google Scholar] [CrossRef]
- Silva, E.R.M.D.; Costa, L.G.S.; Silva, A.D.S.; Souza, E.C.D.; Barbosa, I.C.D.C. Physical-chemical, chemistry and chemometric characterization of underground waters from pirabas and Barreiras Aquifers in municipalities of the state of Pará. Rev. Bras. Geog. Fis. 2018, 11, 1026–1041. [Google Scholar]
- Gomes, D.F.; Sanches, N.A.D.O.; Andrade, D.D.P.; Bastos, W.R. Occurrence of aquatic macroinvertebrates in an extrativist reserve of brazilian Amazon. Rev. Biol. Neotrop. 2019, 16, 50–60. [Google Scholar] [CrossRef]
- Silva, E.C.D.; Gutjahr, A.L.N.; Braga, C.E.D.S. Caracterização físico-química da água de um rio urbano amazônico, Capanema, Pará, Brasil. RSD 2021, 10, e51101622866. [Google Scholar] [CrossRef]
- Burggren, W.W.; Arriaga-Bernal, J.C.; Méndez-Arzate, P.M.; Méndez-Sánchez, J.F. Metabolic physiology of the Mayan cichlid fish (Mayaheros Uropthalmus): Re-examination of classification as an oxyconformer. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2019, 237, 110538. [Google Scholar] [CrossRef]
- Braz-Mota, S.; Almeida-Val, V.M.F. Ecological adaptations of Amazonian fishes acquired during evolution under environmental variations in dissolved oxygen: A review of responses to hypoxia in fishes, featuring the hypoxia-tolerant Astronotus spp. J. Exp. Zool. A 2021, 335, 771–786. [Google Scholar] [CrossRef]
- Saint-Paul, U. Diurnal routine O2 consumption at different O2 concentrations by Colossoma macropomum and Colossoma brachypomus (Teleostei: Serrasalmidae). CBPA 1988, 89, 675–682. [Google Scholar]
- Neves, L.C.; Favero, G.C.; Beier, S.L.; Ferreira, N.S.; Palheta, G.D.A.; Melo, N.F.A.C.; Luz, R.K. Physiological and metabolic responses in juvenile Colossoma macropomum exposed to hypoxia. Fish Physiol. Biochem. 2020, 46, 2157–2167. [Google Scholar] [CrossRef]
- Neves, L.C.; Silva, W.S.; Ferreira, A.L.; Favero, G.C.; Beier, S.L.; Palheta, G.D.A.; Melo, N.F.A.C.; Luz, R.K. Physiological responses of juvenile Colossoma macropomum after different periods of air exposure. Aquaculture 2022, 548, 737583. [Google Scholar] [CrossRef]
- Lima, J.A.F.; Ferrari, V.A.; Colares de Melo, J.S.; Gaspar, L.A.; Chabalin, E.; Santos, E.P. Comportamento do pacu, Colossoma mitrei, em um cultivo experimental, no centro-oeste do Brasil. Bol. Técnico CEPTA Cent. Pesqui. Treinamento Aquicultura 1988, 1, 15–29. [Google Scholar]
- Pinheiro, M.S.L.; Selini Dorce, L.; Momo Ziemniczak, H.; Honorato Da Silva, C.A.; Hertes Neu, D. Toxicidade aguda da amônia em pacu (Piaractus mesopotamicus). Rev. Acad. Ciênc. Anim. 2021, 19, 1. [Google Scholar] [CrossRef]
- Quaresma, F.D.S.; Santos, F.L.B.D.; Ribeiro, P.F.; Leite, L.A.; Sampaio, A.H. Acute toxicity of non-ionized ammonia on tambacu (Colossoma macropomum x Piaractus mesopotamicus). Rev. Ciên. Agron. 2020, 51, e20186277. [Google Scholar] [CrossRef]
- Dairiki, J.K.; Da Silva, T.B.A. Revisão de Literatura: Exigências Nutricionais do Tambaqui—Compilação de Trabalhos, Formulação de Ração a e Desafios Futuros; Embrapa Amazônica Ocidental: Manaus, Brazil, 2011; 44p. [Google Scholar]
- Izel, A.C.U.; Crescencio, R.; O’Sullivan, F.L.d.A.; Chagas, E.C.; Boijink, C.d.L. Cultivo do Tambaqui no Amazonas; ABC da Agricultura Familiar; Embrapa Amazônica Ocidental: Manaus, Brazil, 2014; 51p. [Google Scholar]
- Maeda, H.; Silva, P.C.; Aguiar, M.; Padua, D.M.C.; Oliveira, R.P.C.; Machado, N.P.; Rodrigues, V.; Silva, R.H. Efeitos da densidade de estocagem na segunda alevinagem de tilápia nilótica (Oreochromis niloticus), em sistema raceway. R. Bras. Zootec. 2006, 7, 265–272. [Google Scholar]
- Li, H.W.; Brocksen, R.W. Approaches to the analysis of energetic costs of intraspecific competition for space by rainbow trout (Salmo gairdneri). J. Fish Biol. 1977, 11, 329–341. [Google Scholar] [CrossRef]
- Melo, D.C.; Oliveira, D.A.A.; Melo, M.M.; Júnior, D.V.; Teixeira, E.A.; Guimarães, S.R. Perfil proteico de tilápia nilótica chitralada (Oreochromis niloticus), submetida ao estresse crônico por hipóxia. Arq. Bras. Med. Vet. Zootec. 2009, 61, 1183–1190. [Google Scholar]
- Carvalho, M.A.M.D.; Costa, R.B.D.; Silva, L.D.A.; Oliveira, C.G.D.; Miranda, L.A.M.; Martins, L.P.; Sales, R.D.O.; Farias, J.O. Crescimento do curimatã comum, Prochilodus cearaensis (Steindachner,1911) em sistema de recirculação de água (SRA) em três diferentes densidades de estocagem. Rev. Bras. Hig. San. Anim. 2020, 14, 1–8. [Google Scholar]
- Ali, M.; Iqbal, F.; Salam, A.; Sial, F.; Athar, M. Comparative study of body composition of four fish species in relation to pond depth. Int. J. Environ. Sci. Technol. 2006, 2, 359–364. [Google Scholar]
- Angeles-Escobar, B.E.; Silva, S.M.B.C.; Severi, W. Growth, red blood cells, and gill alterations of red pacu (Piaractus brachypomus) fingerlings by chronic exposure to different total suspended solids in biofloc. J. World Aquac. Soc. 2022, 53, 652–668. [Google Scholar]
- Houlihan, D.; Boujard, T.; Jobling, M. (Eds.) Food Intake in Fish; John Wiley & Sons: Hoboken, NJ, USA, 2008. [Google Scholar]
- Liu, Z.-S.; Zhang, L.; Chen, W.-L.; He, C.-F.; Qian, X.-Y.; Liu, W.-B.; Li, X.-F. Insights into the interaction between stocking density and feeding rate in fish Megalobrama ambylcephala based on growth performance, innate immunity, antioxidant activity, and the GH-IGF1 axis. Aquaculture 2024, 580, 740355. [Google Scholar]
- Izel-Silva, J.; Ono, E.A.; Queiroz, M.N.; Dos-Santos, R.B.; Affonso, E.G. Aeration strategy in the intensive culture of tambaqui, Colossoma macropomum, in the tropics. Aquaculture 2020, 529, 735644. [Google Scholar]
- Torrans, L.; Ott, B.; Bosworth, B. Impact of minimum daily dissolved oxygen concentration on production performance of hybrid female channel catfish × male blue catfish. N. Am. J. Aquac. 2015, 77, 485–490. [Google Scholar]
- Gamperl, A.K.; Ajiboye, O.O.; Zanuzzo, F.S.; Sandrelli, R.M.; Peroni, E.D.F.C.; Beemelmanns, A. The impacts of increasing temperature and moderate hypoxia on the production characteristics, cardiac morphology and haematology of Atlantic salmon (Salmo salar). Aquaculture 2020, 519, 734874. [Google Scholar]
- Luckenbach, J.A.; Murashige, R.; Daniels, H.V.; Godwin, J.; Borski, R.J. Temperature affects insulin-like growth factor I and growth of juvenile southern flounder, Paralichthys lethostigma. Comp. Biochem. Physiol. A 2007, 146, 95–104. [Google Scholar]
- Urbinati, E.C.; Zanuzzo, F.S.; Biller-Takahashi, J.D. Estresse e sistema imune em peixes. In Biologia e Fisiologia de Peixes Neotropicais de Água Doce; Baldisseroto, B., Cyrino, J.E.P., Urbinati, E.C., Eds.; FUNEP: Jaboticabal, Brazil, 2014; pp. 87–105. [Google Scholar]
- Barton, B.A.; Iwama, G.K. Physiological changes in fish from stress in aquaculture with emphasis on the response and effects of corticosteroids. Annu. Rev. Fish Dis. 1991, 1, 3–26. [Google Scholar]
- Qi, C.; Xie, C.; Tang, R.; Qin, X.; Wang, D.; Li, D. Effect of stocking density on growth, physiological responses, and body composition of juvenile blunt snout bream, Megalobrama amblycephala. J World Aquac. Soc. 2016, 47, 358–368. [Google Scholar] [CrossRef]
- Bacchetta, C.; Rossi, A.S.; Ale, A.; Cazenave, J. Physiological effects of stocking density on the fish Piaractus mesopotamicus fed with red seaweed (Pyropia columbina) and β-carotene-supplemented diets. Aquac. Res. 2020, 51, 1992–2003. [Google Scholar] [CrossRef]
- Baldisserotto, B. Fisiologia de Peixes Aplicada à Piscicultura; UFSM: Santa Maria, Brazil, 2002; 49p. [Google Scholar]
- Guo, H.-Y.; Dong, X.-Y.; Zhang, X.-M.; Zhang, P.-D.; Li, W.-T. Survival, growth and physiological responses of juvenile Japanese flounder (Paralichthys olivaceus, Temminck & Schlegel, 1846) exposed to different dissolved oxygen concentrations and stocking densities. J. Appl. Ichthyol. 2017, 33, 731–739. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stocking Densities | NH3 (mg/L) | Temperature (°C) | pH |
|---|---|---|---|
| Experiment I | |||
| D0.68 | 0.0153 ± 0.01 | 28.44 ± 0.45 | 6.53 ± 0.05 |
| D1.45 | 0.0153 ± 0.01 | 28.34 ± 0.38 | 6.50 ± 0.08 |
| D4.41 | 0.0153 ± 0.01 | 28.29 ± 0.27 | 6.48 ± 0.10 |
| D7.17 | 0.0153 ± 0.00 | 28.19 ± 0.57 | 6.53 ± 0.10 |
| p-value | 0.7935 | 0.9928 | 0.8015 |
| Experiment II | |||
| D1 | 0.0256 ± 0.00 | 26.60 ± 2.63 | 7.07 ± 0.05 |
| D1.95 | 0.0270 ± 0.00 | 25.75 ± 0.02 | 7.03 ± 0.05 |
| D5.63 | 0.0265 ± 0.00 | 25.80 ± 0.09 | 7.03 ± 0.05 |
| D7.9 | 0.0261 ± 0.00 | 25.77 ± 0.10; | 7.03 ± 0.05 |
| p-value | 0.0770 ± 0.00 | 0.2527 | 0.8592 |
| Parameter | Stocking Density | Overall p-Value | Linear p-Value | Quadratic p-Value | |||
|---|---|---|---|---|---|---|---|
| D1 | D1.95 | D5.63 | D7.90 | ||||
| GLU (mg/dL) | 95.07 ± 1.21 | 81.55 ± 8.49 | 81.35 ± 6.26 | 93.75 ± 3.17 | 0.1505 | 0.7175 | 0.0520 |
| TC (mg/dL) | 100.29 ± 3.62 | 114.42 ± 9.29 | 109.87 ± 9.74 | 84.45 ± 7.79 | 0.1121 | 0.0891 | 0.0858 |
| TG (mg/dL) | 207.21 ± 26.46 ab | 246.78 ± 53.22 a | 175.54 ± 19.73 ab | 142.88 ± 18.13 b | 0.0233 | 0.0067 * | 0.7374 |
| tHb (g/dL) | 8.20 ± 0.48 a | 6.45 ± 0.47 b | 7.11 ± 0.22 ab | 6.59 ± 0.32 b | 0.0472 | 0.1068 | 0.5191 |
| Ht (%) | 33.50 ± 1.23 | 23.83 ± 2.47 | 26.78 ± 2.69 | 28.92 ± 2.40 | 0.0829 | 0.7255 | 0.1201 |
| TPP (g/dL) | 4.88 ± 0.09 | 4.98 ± 0.16 | 5.07 ± 0.07 | 4.72 ± 0.15 | 0.2905 | 0.3333 | 0.1028 |
| RBC (×106/μL) | 1.12 ± 0.18 | 1.38 ± 0.10 | 1.24 ± 0.07 | 1.28 ± 0.12 | 0.5376 | 0.7820 | 0.7500 |
| AST (UI/L) | 39.42 ± 7.27 | 28.91 ± 2.91 | 31.01 ± 2.70 | 33.09 ± 4.28 | 0.4609 | 0.6670 | 0.3561 |
| ALT (UI/L) | 9.29 ± 0.16 | 7.23 ± 0.36 | 8.48 ± 0.83 | 9.43 ± 0.83 | 0.1199 | 0.2589 | 0.2082 |
| MCV (ftl) | 317.56 ± 44.49 a | 178.57 ± 5.41 b | 232.13 ± 32.25 ab | 198.76 ± 16.27 b | 0.0387 | 0.1300 | 0.4233 |
| MCH (pg) | 74.42 ± 11.70 | 48.92 ± 3.54 | 59.29 ± 2.57 | 55.44 ± 4.60 | 0.1210 | 0.3371 | 0.4499 |
| MCHC (g/dL) | 24.77 ± 1.32 | 27.56 ± 2.62 | 27.85 ± 3.33 | 21.06 ± 1.84 | 0.2392 | 0.2103 | 0.1124 |
| VSI (%) | 7.96 ± 0.46 | 7.28 ± 0.20 | 7.01 ± 0.31 | 7.42 ± 0.31 | 0.3029 | 0.3900 | 0.1329 |
| HSI (%) | 2.00 ± 0.07 | 1.85 ± 0.16 | 2.00 ± 0.14 | 1.90 ± 0.26 | 0.8952 | 0.9470 | 0.8391 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Morais Carvalho Ananias, I.; dos Santos Silva, S.; de Sena Souza, A.; dos Santos, F.A.C.; Magalhães, T.B.; de Freitas Martins, W.J.; Favero, G.C.; Luz, R.K. Effects of Stocking Densities on the Growth Performance and Physiology of Juvenile Piaractus brachypomus in Recirculating Aquaculture System. Fishes 2025, 10, 171. https://doi.org/10.3390/fishes10040171
de Morais Carvalho Ananias I, dos Santos Silva S, de Sena Souza A, dos Santos FAC, Magalhães TB, de Freitas Martins WJ, Favero GC, Luz RK. Effects of Stocking Densities on the Growth Performance and Physiology of Juvenile Piaractus brachypomus in Recirculating Aquaculture System. Fishes. 2025; 10(4):171. https://doi.org/10.3390/fishes10040171
Chicago/Turabian Stylede Morais Carvalho Ananias, Imaculada, Sidney dos Santos Silva, André de Sena Souza, Fábio Aremil Costa dos Santos, Thamara Bentivole Magalhães, Wagner Junio de Freitas Martins, Gisele Cristina Favero, and Ronald Kennedy Luz. 2025. "Effects of Stocking Densities on the Growth Performance and Physiology of Juvenile Piaractus brachypomus in Recirculating Aquaculture System" Fishes 10, no. 4: 171. https://doi.org/10.3390/fishes10040171
APA Stylede Morais Carvalho Ananias, I., dos Santos Silva, S., de Sena Souza, A., dos Santos, F. A. C., Magalhães, T. B., de Freitas Martins, W. J., Favero, G. C., & Luz, R. K. (2025). Effects of Stocking Densities on the Growth Performance and Physiology of Juvenile Piaractus brachypomus in Recirculating Aquaculture System. Fishes, 10(4), 171. https://doi.org/10.3390/fishes10040171