
Effects of CO2 and O2 in Modified Atmosphere Packaging on Water Retention, Protein Stability, and Microbial Growth in Atlantic Salmon Fillets

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Preparation and Modified Atmosphere Packaging
2.2. Microbiological Analysis
2.3. Total Volatile Basic Nitrogen (TVB-N) Analysis
2.4. Cooking Loss Analysis
2.5. Protein Oxidation Determination
2.6. SDS–Polyacrylamide Gel Electrophoresis (SDS-PAGE)
2.7. Fourier Transform Infrared Spectroscopy (FTIR) Analysis
2.8. Low-Field Nuclear Magnetic Resonance (LF-NMR) Analysis
2.9. Statistical Analysis
3. Results
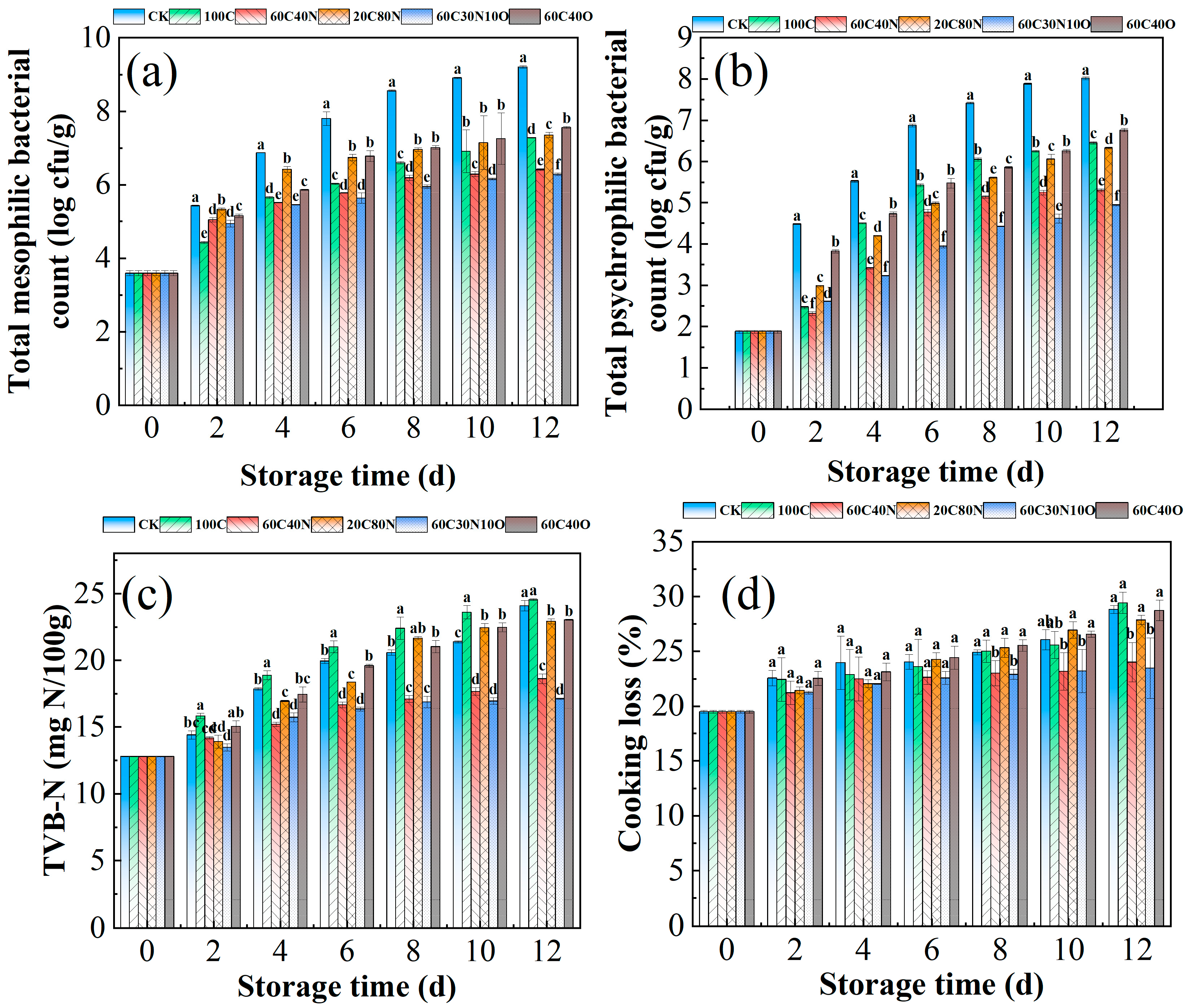
3.1. Microbial Growth
3.2. Changes in TVB-N Content
3.3. Changes in Cooking Loss
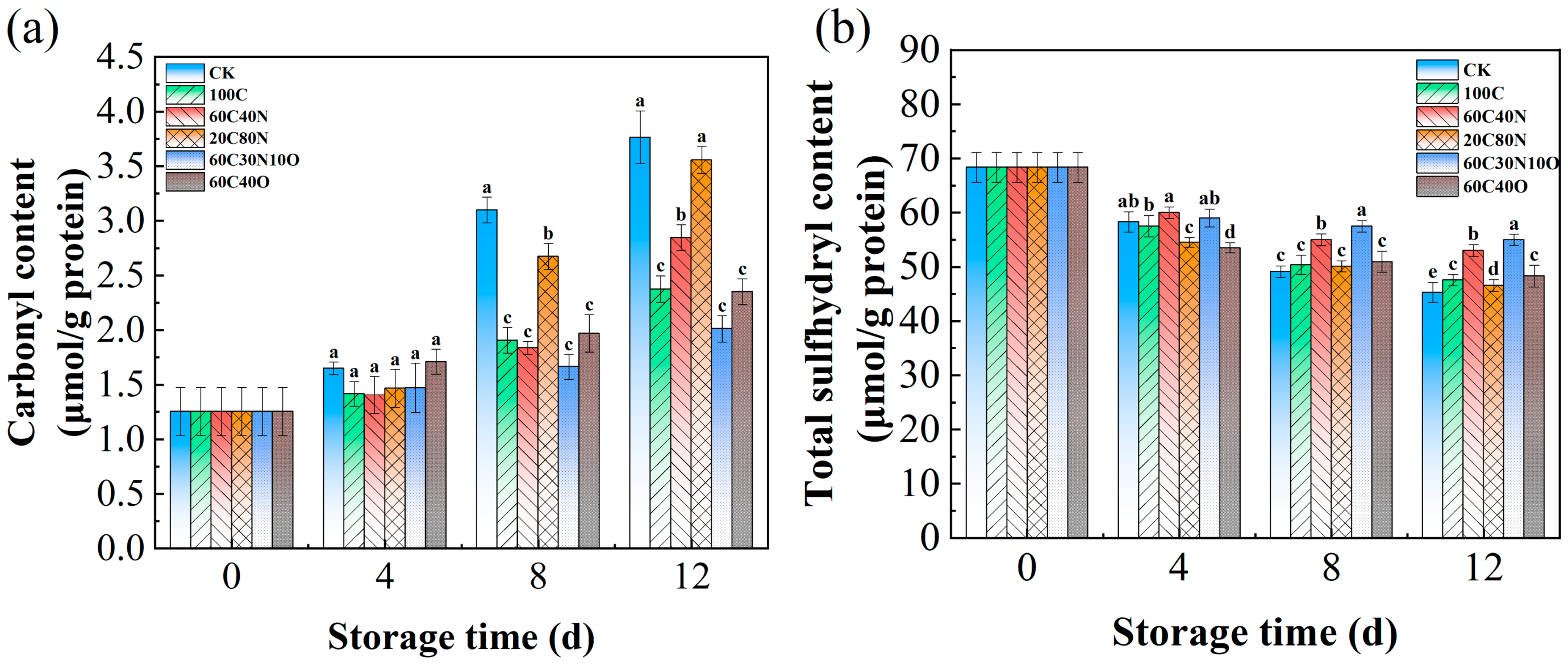
3.4. Changes of Carbonyl Content and Total Sulfhydryl Content
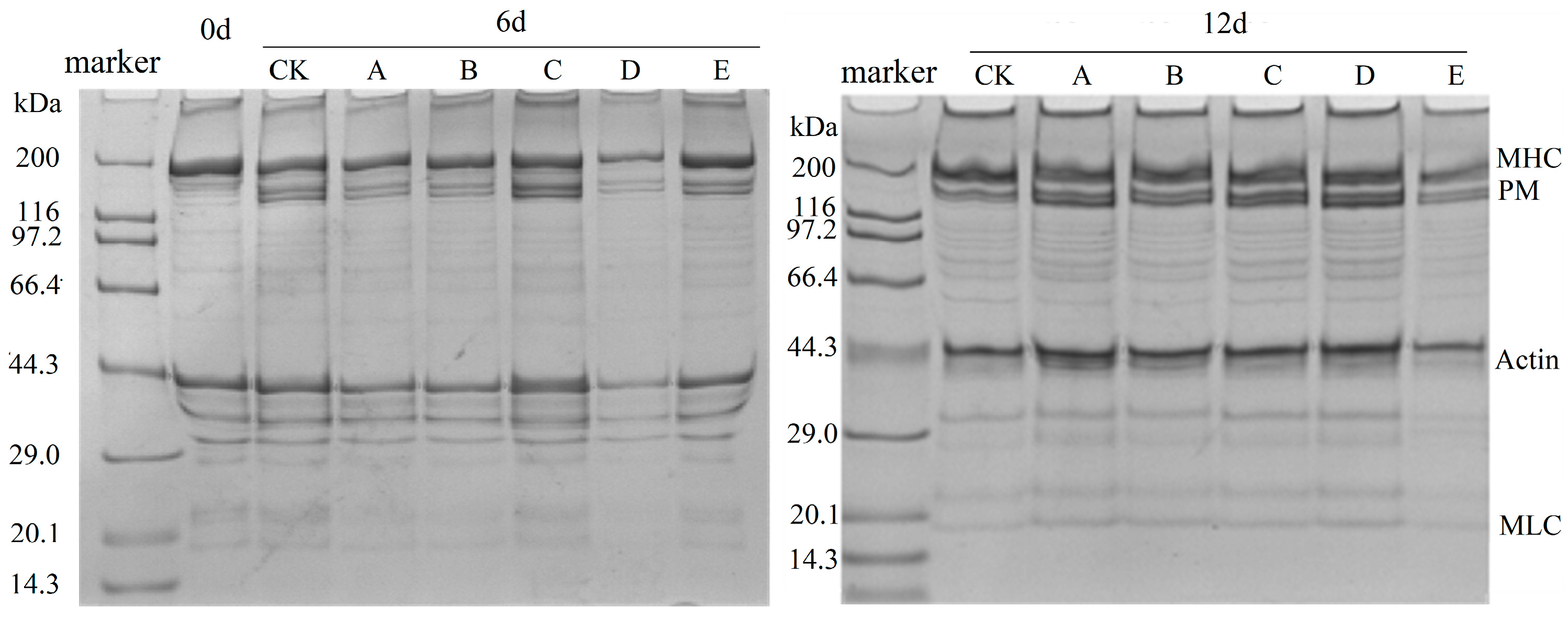
3.5. SDS-PAGE
3.6. Secondary Structure
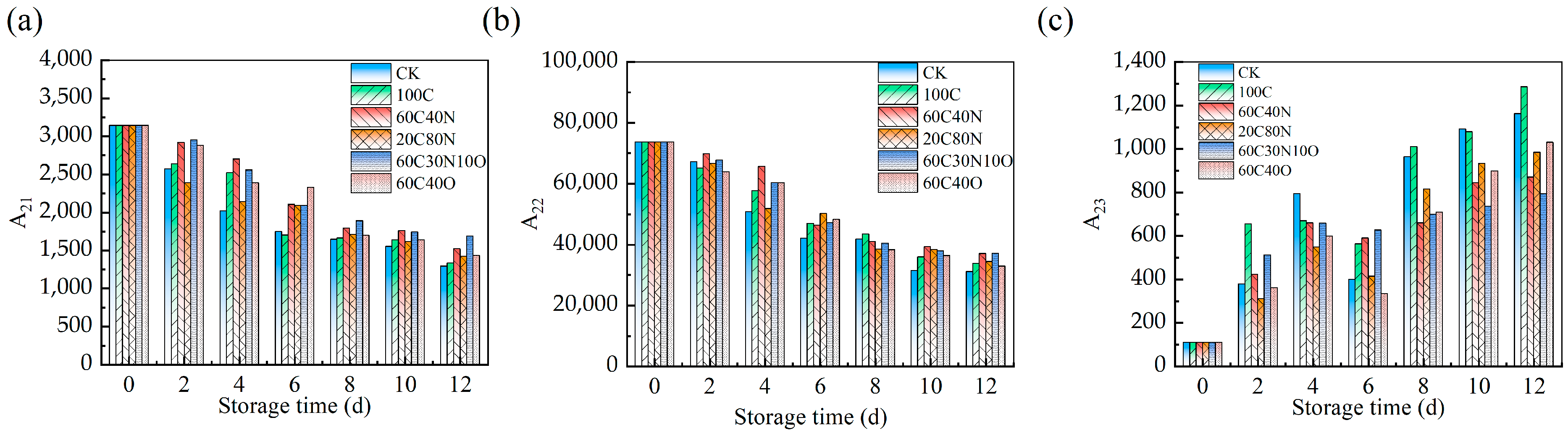
3.7. LF-NMR
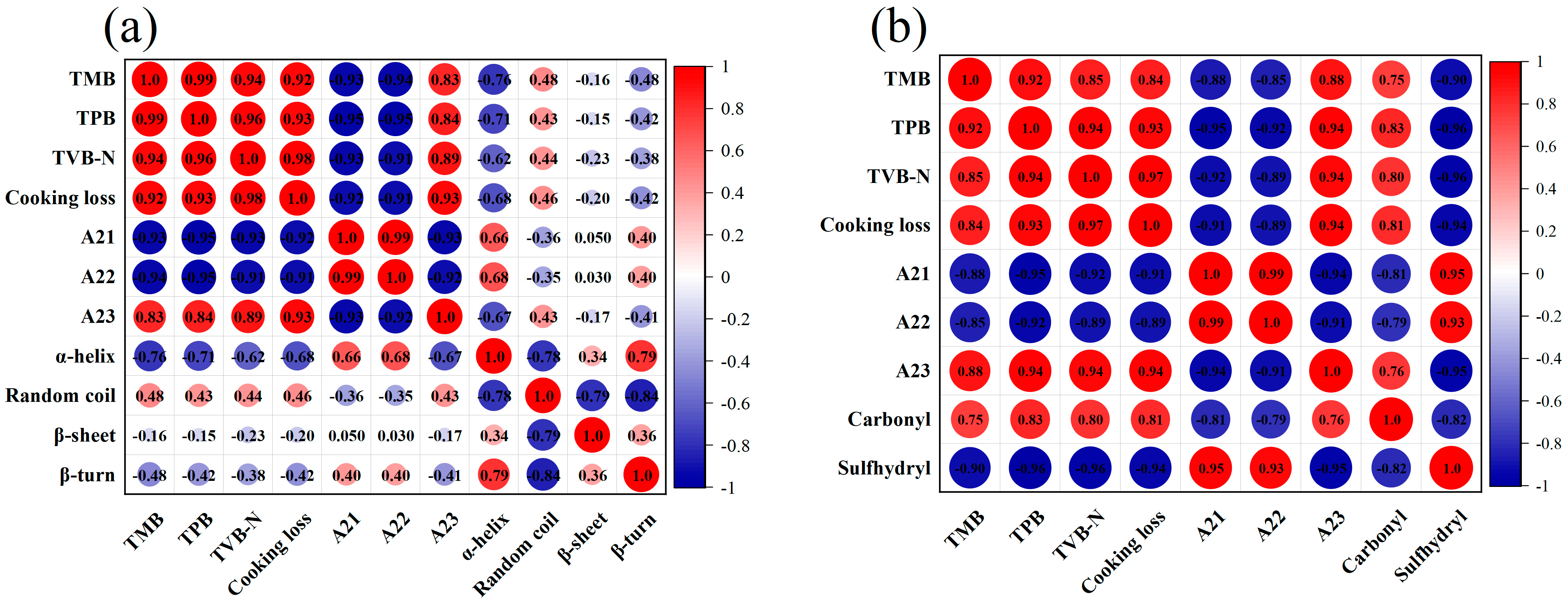
3.8. Correlation Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chan, S.S.; Roth, B.; Jessen, F.; Løvdal, T.; Jakobsen, A.N.; Lerfall, J. A comparative study of Atlantic salmon chilled in refrigerated seawater versus on ice: From whole fish to cold-smoked fillets. Sci. Rep. 2020, 10, 17160. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Majumdar, R.K.; Mehta, N.K.; Ngasotter, S.; Gaurav, K.; Tripathi, M. Evaluating the efficacy of citrus fruit peel extract in preserving the quality of silver carp (Hypophthalmichthys molitrix) surimi during frozen storage. J. Agric. Food Res. 2024, 18, 101440. [Google Scholar] [CrossRef]
- Chan, S.S.; Roth, B.; Skare, M.; Hernar, M.; Jessen, F.; Løvdal, T.; Jakobsen, A.N.; Lerfall, J. Effect of chilling technologies on water holding properties and other quality parameters throughout the whole value chain: From whole fish to cold-smoked fillets of Atlantic salmon (Salmo salar). Aquaculture 2020, 526, 735381. [Google Scholar] [CrossRef]
- Nawaz, A.; Irshad, S.; Ali Khan, I.; Khalifa, I.; Walayat, N.; Muhammad Aadil, R.; Kumar, M.; Wang, M.; Chen, F.; Cheng, K.-W.; et al. Protein oxidation in muscle-based products: Effects on physicochemical properties, quality concerns, and challenges to food industry. Food Res. Int. 2022, 157, 111322. [Google Scholar] [CrossRef]
- Bao, Y.; Boeren, S.; Ertbjerg, P. Myofibrillar protein oxidation affects filament charges, aggregation and water-holding. Meat Sci. 2018, 135, 102–108. [Google Scholar] [CrossRef]
- Nikoo, M.; Benjakul, S.; Ahmadi Gavlighi, H.; Xu, X.; Regenstein, J.M. Hydrolysates from rainbow trout (Oncorhynchus mykiss) processing by-products: Properties when added to fish mince with different freeze-thaw cycles. Food Biosci. 2019, 30, 100418. [Google Scholar] [CrossRef]
- Bai, X.; Yin, F.; Ru, A.; Li, M.; Tian, W.; Zhang, G.; Chen, Q.; Chai, R.; Liu, Y.; Cui, W.; et al. Myosin heavy chain isoform expression and meat quality characteristics of different muscles in yak (Bos grunniens). Meat Sci. 2024, 209, 109414. [Google Scholar] [CrossRef]
- Cao, L.; Rasco, B.A.; Tang, J.; Niu, L.; Lai, K.; Fan, Y.; Huang, Y. Effects of freshness on the cook loss and shrinkage of grass carp (Ctenopharyngodon idellus) fillets following pasteurization. Int. J. Food Prop. 2016, 19, 2297–2306. [Google Scholar] [CrossRef]
- Van Haute, S.; Raes, K.; Devlieghere, F.; Sampers, I. Combined use of cinnamon essential oil and MAP/vacuum packaging to increase the microbial and sensorial shelf life of lean pork and salmon. Food Packag. Shelf Life 2017, 12, 51–58. [Google Scholar] [CrossRef]
- Wang, G.; Ma, F.; Zeng, L.; Bai, Y.; Wang, H.; Xu, X.; Zhou, G. Modified atmosphere packaging decreased Pseudomonas fragi cell metabolism and extracellular proteolytic activities on meat. Food Microbiol. 2018, 76, 443–449. [Google Scholar] [CrossRef]
- Liang, Z.; Veronica, V.; Huang, J.; Zhang, P.; Fang, Z. Combined effects of plant food processing by-products and high oxygen modified atmosphere packaging on the storage stability of beef patties. Food Control 2022, 133, 108586. [Google Scholar] [CrossRef]
- Tan, N.; Shao, Y.; Xu, Y.; Li, Z.; Huang, Z.; Zhang, W.; Deng, S.; Zhang, B.; Zhang, L.; Yuan, P. Tandem mass tag-based quantitative proteomics analysis of modified atmosphere packaging in large yellow croaker (Pseudosciaena crocea) fillets during refrigerated storage. Food Chem. 2025, 463, 141744. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Hu, S.; Gao, Y.; Ye, C.; Wang, H. Effect of collagen-lysozyme coating on fresh-salmon fillets preservation. LWT-Food Sci. Technol. 2017, 75, 59–64. [Google Scholar] [CrossRef]
- Zhang, L.; Li, Q.; Hong, H.; Luo, Y. Prevention of protein oxidation and enhancement of gel properties of silver carp (Hypophthalmichthys molitrix) surimi by addition of protein hydrolysates derived from surimi processing by-products. Food Chem. 2020, 316, 126343. [Google Scholar] [CrossRef] [PubMed]
- Niamnuy, C.; Devahastin, S.; Soponronnarit, S. Changes in protein compositions and their effects on physical changes of shrimp during boiling in salt solution. Food Chem. 2008, 108, 165–175. [Google Scholar] [CrossRef]
- Eranda, D.H.U.; Chaijan, M.; Uysal-Unalan, I.; Panpipat, W.; Naik, A.S.; Dib, A.L.; Karnjanapratum, S.; Gagaoua, M. Biopreservation of pre-processed fresh fish by bio-based coatings: A single strategy with multiple benefits towards waste prevention. Food Biosci. 2024, 58, 103696. [Google Scholar] [CrossRef]
- Chen, B.; Mei, J.; Xie, J. Effects of packaging methods and temperature variations on the quality and microbial diversity of grouper (Epinephelus Lanceolatus) during cold storage. Food Biosci. 2024, 60, 104315. [Google Scholar] [CrossRef]
- Ozogul, F.; Durmuş, M.; Kosker, A.R.; Özkütük, A.S.; Kuley, E.; Yazgan, H.; Yazgan, R.; Simat, V.; Ozogul, Y. The impact of marine and terrestrial based extracts on the freshness quality of modified atmosphere packed sea bass fillets. Food Biosci. 2023, 53, 102545. [Google Scholar] [CrossRef]
- Zhang, J.; Li, Y.; Liu, X.; Lei, Y.; Regenstein, J.M.; Luo, Y. Characterization of the microbial composition and quality of lightly salted grass carp (Ctenopharyngodon idellus) fillets with vacuum or modified atmosphere packaging. Int. J. Food Microbiol. 2019, 293, 87–93. [Google Scholar] [CrossRef]
- Qian, Y.; Liu, C.; Zhang, J.; Ertbjerg, P.; Yang, S. Effects of modified atmosphere packaging with varied CO2 and O2 concentrations on the texture, protein, and odor characteristics of salmon during cold storage. Foods 2022, 11, 3560. [Google Scholar] [CrossRef]
- Yang, Z.; Yan, J.; Xie, J. Effect of vacuum and modified atmosphere packaging on moisture state, quality, and microbial communities of grouper (Epinephelus coioides) fillets during cold storage. Food Res. Int. 2023, 173, 113340. [Google Scholar] [CrossRef]
- Lerfall, J.; Bjørge Thomassen, G.M.; Jakobsen, A.N. Quality of fresh saithe (Pollachius virens) in modified atmosphere packages as affected by the gas composition. Food Packag. Shelf Life 2018, 18, 147–156. [Google Scholar] [CrossRef]
- McSharry, S.; Koolman, L.; Whyte, P.; Bolton, D. An investigation of the survival and/or growth of Clostridioides (Clostridium) difficile in beef stored under aerobic, anaerobic and commercial vacuum packaging conditions at 2 °C and 20 °C. Food Control 2021, 119, 107475. [Google Scholar] [CrossRef]
- Bekhit, A.E.-D.A.; Holman, B.W.B.; Giteru, S.G.; Hopkins, D.L. Total volatile basic nitrogen (TVB-N) and its role in meat spoilage: A review. Trends Food Sci. Technol. 2021, 109, 280–302. [Google Scholar] [CrossRef]
- Vargas Baldi, S.C.; Parisi, G.; Bonelli, A.; Balieiro, J.C.C.; Lapa Guimarães, J.; Macedo Viegas, E.M. Effects of different stunning/slaughter methods on frozen fillets quality of cobia (Rachycentron canadum). Aquaculture 2018, 486, 107–113. [Google Scholar] [CrossRef]
- Wang, Y.-Y.; Wang, H.; Zhou, F.; Wu, Y.; Ma, H.; Zhao, R.; He, J.; Gu, Z. Effect of ultrasonic thawing temperature on the quality of quick-frozen small yellow croaker (Larimichthys polyactis) and its possible mechanisms. LWT-Food Sci. Technol. 2023, 179, 114620. [Google Scholar] [CrossRef]
- Sun, B.; Zhao, Y.; Yu, J.; Ling, J.; Shang, H.; Liu, Z. The combined efficacy of superchilling and high CO2 modified atmosphere packaging on shelf life and quality of swimming crab (Portunus trituberculatus). J. Aquat. Food Prod. Technol. 2017, 26, 655–664. [Google Scholar] [CrossRef]
- Ma, C.; Zhang, J.; Zhang, R.; Zhou, L.; Ni, L.; Zhang, W. Study on the effects of pre-slaughter transport stress on water holding capacity of pork: Insights from oxidation, structure, function, and degradation properties of protein. Food Chem. X 2024, 24, 101913. [Google Scholar] [CrossRef]
- He, Y.; Huang, H.; Li, L.; Yang, X.; Hao, S.; Chen, S.; Deng, J. The effects of modified atmosphere packaging and enzyme inhibitors on protein oxidation of tilapia muscle during iced storage. LWT-Food Sci. Technol. 2018, 87, 186–193. [Google Scholar] [CrossRef]
- Vate, N.K.; Benjakul, S. Combined effect of squid ink tyrosinase and tannic acid on heat induced aggregation of natural actomyosin from sardine. Food Hydrocol. 2016, 56, 62–70. [Google Scholar] [CrossRef]
- Li, P.; Zhou, Q.; Chu, Y.; Lan, W.; Mei, J.; Xie, J. Effects of chitosan and sodium alginate active coatings containing ε-polysine on qualities of cultured pufferfish (Takifugu obscurus) during cold storage. Int. J. Biol. Macromol. 2020, 160, 418–428. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Tan, Y.; Luo, Y.; Li, X.; Hong, H. Evidence of myofibrillar protein oxidation and degradation induced by exudates during the thawing process of bighead carp fillets. Food Chem. 2024, 434, 137396. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.; Hu, J.; Gao, B.; Zhang, B.; Li, P.; Shang, N. Effects of the interaction between Aeromonas sobria and Macrococcus caseolyticus on protein degradation of refrigerated sturgeon fillets: Novel perspective on fish spoilage. LWT-Food Sci. Technol. 2023, 183, 114908. [Google Scholar] [CrossRef]
- Martínez, M.A.; Velazquez, G.; Cando, D.; Núñez-Flores, R.; Borderías, A.J.; Moreno, H.M. Effects of high pressure processing on protein fractions of blue crab (Callinectes sapidus) meat. Innov. Food Sci. Emerg. Technol. 2017, 41, 323–329. [Google Scholar] [CrossRef]
- Chan, S.S.; Roth, B.; Jessen, F.; Jakobsen, A.N.; Lerfall, J. Water holding properties of Atlantic salmon. Compr. Rev. Food Sci. Food Saf. 2022, 21, 477–498. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qian, Y.-F.; Shi, C.-J.; Liu, C.-C.; Zhang, J.-J.; Yang, S.-P. Effects of CO2 and O2 in Modified Atmosphere Packaging on Water Retention, Protein Stability, and Microbial Growth in Atlantic Salmon Fillets. Fishes 2025, 10, 141. https://doi.org/10.3390/fishes10040141
Qian Y-F, Shi C-J, Liu C-C, Zhang J-J, Yang S-P. Effects of CO2 and O2 in Modified Atmosphere Packaging on Water Retention, Protein Stability, and Microbial Growth in Atlantic Salmon Fillets. Fishes. 2025; 10(4):141. https://doi.org/10.3390/fishes10040141
Chicago/Turabian StyleQian, Yun-Fang, Cheng-Jian Shi, Cheng-Cheng Liu, Jing-Jing Zhang, and Sheng-Ping Yang. 2025. "Effects of CO2 and O2 in Modified Atmosphere Packaging on Water Retention, Protein Stability, and Microbial Growth in Atlantic Salmon Fillets" Fishes 10, no. 4: 141. https://doi.org/10.3390/fishes10040141
APA StyleQian, Y.-F., Shi, C.-J., Liu, C.-C., Zhang, J.-J., & Yang, S.-P. (2025). Effects of CO2 and O2 in Modified Atmosphere Packaging on Water Retention, Protein Stability, and Microbial Growth in Atlantic Salmon Fillets. Fishes, 10(4), 141. https://doi.org/10.3390/fishes10040141