
Carbon Nanotube-Based Drug Delivery System Increases Drug Content and Promotes Immune Response in Mandarin Fish


Abstract
1. Introduction
2. Materials and Methods
2.1. Compounds, Virus, and Experimental Fish
2.2. Preparation and Characterization of Drug Delivery Systems
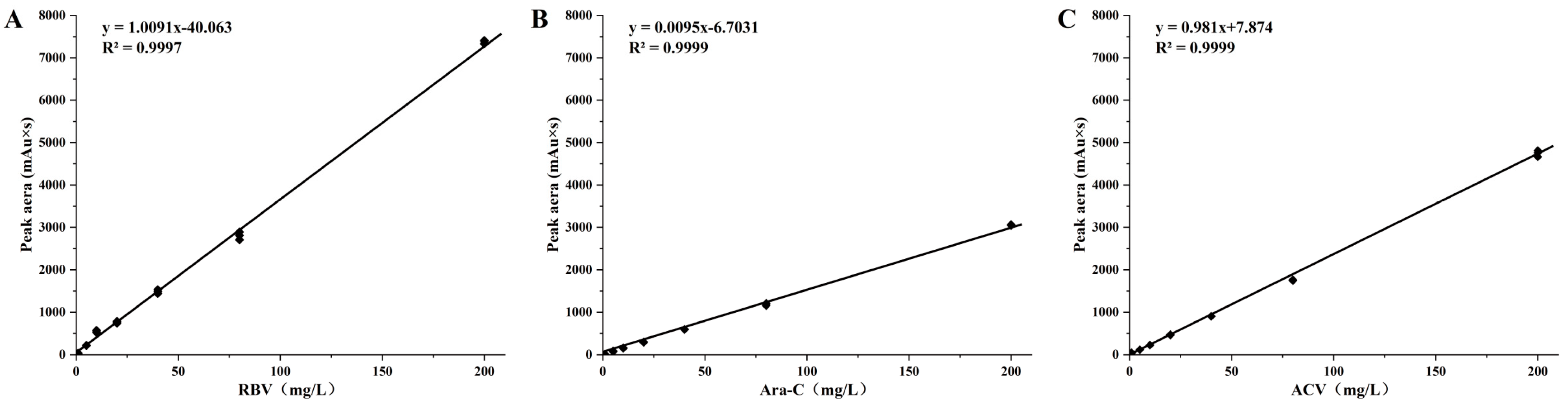
2.3. Drugs Loadings in Delivery System
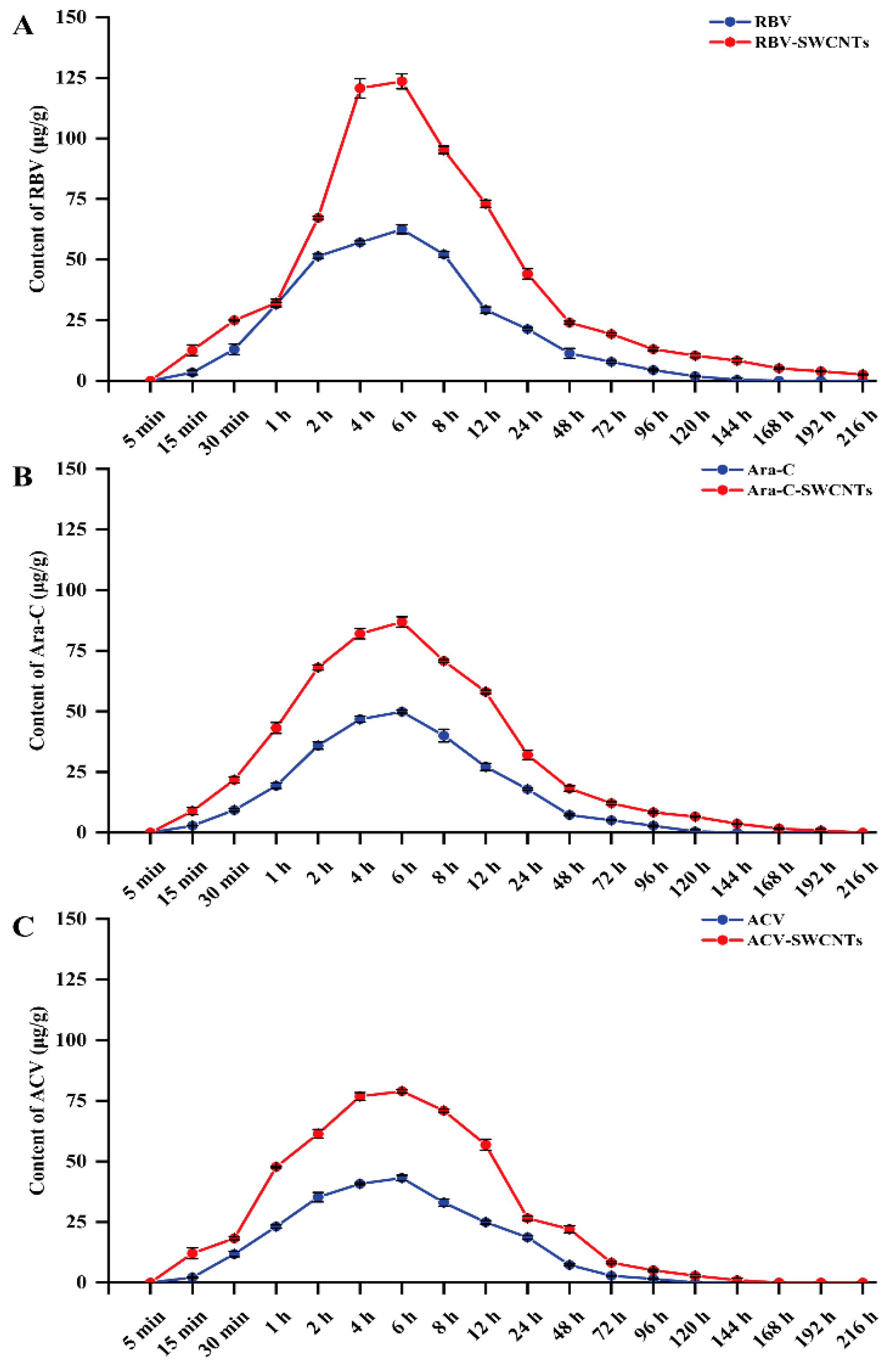
2.4. Drugs Metabolism in Kidney Tissue
2.5. Infection of Mandarin Fish with ISKNV and Drug Treatment
2.6. qPCR Analysis of Infection Rates of ISKNV and Expression of Immune-Related Genes
2.7. Measurement of Enzyme Activities
2.8. Statistical Analysis
3. Results
3.1. Synthesis and Characterization of Drug Delivery Systems
3.2. Drug Loading in Delivery Systems and Drugs Metabolism in Kidney Tissues
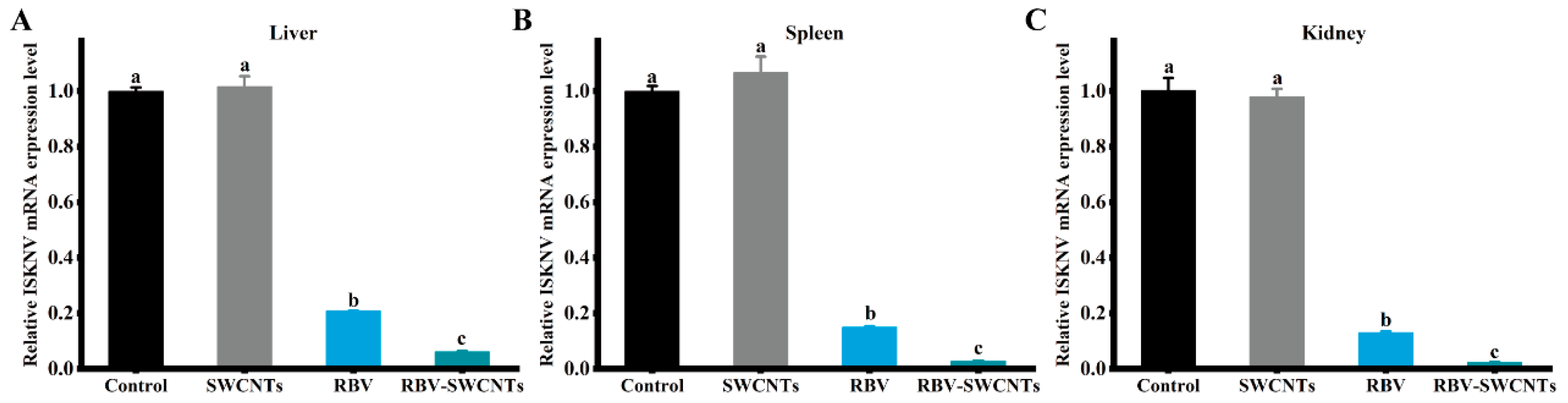
3.3. Infection Rate of Virus
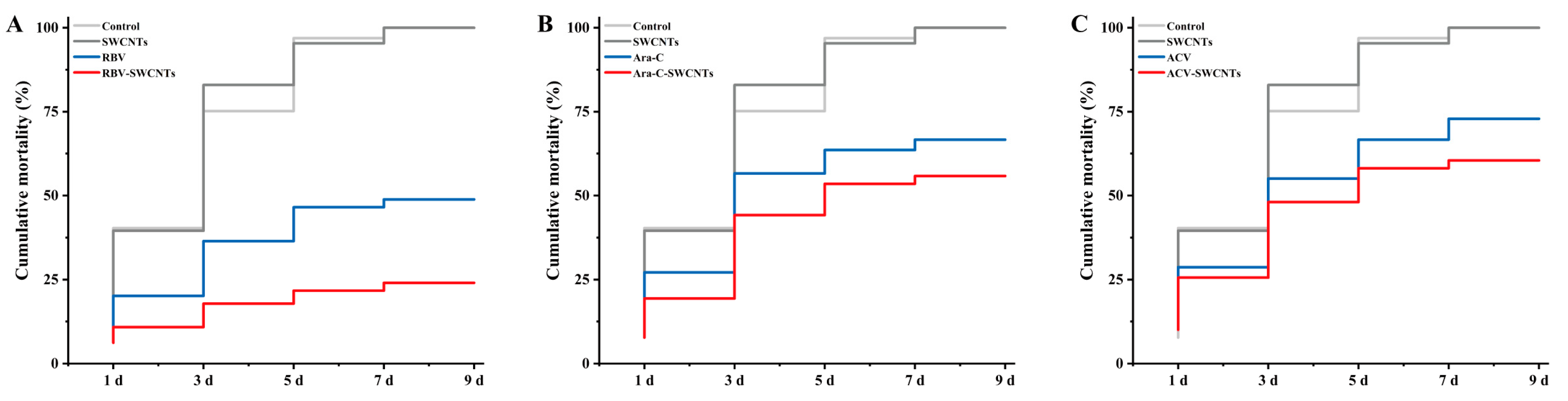
3.4. Cumulative Mortality
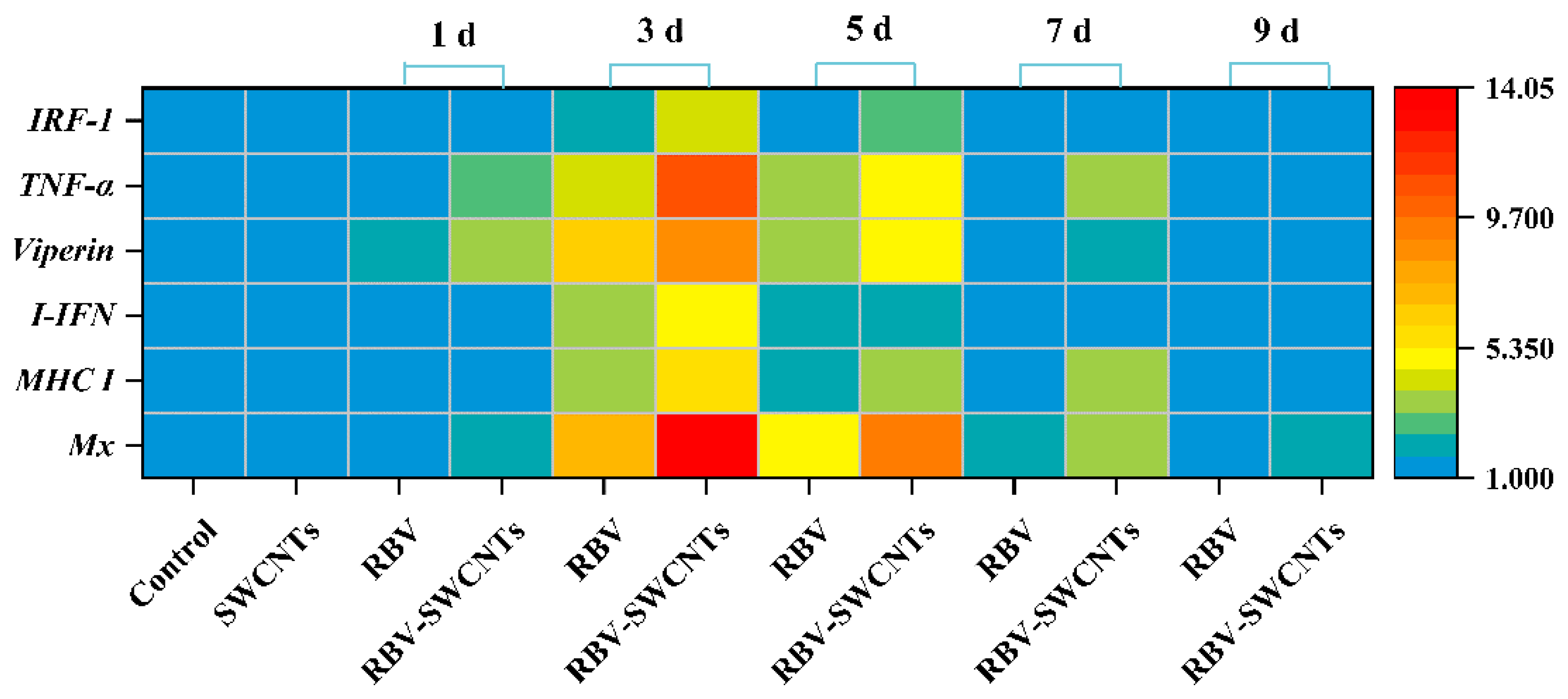
3.5. Expression of Immune-Related Genes
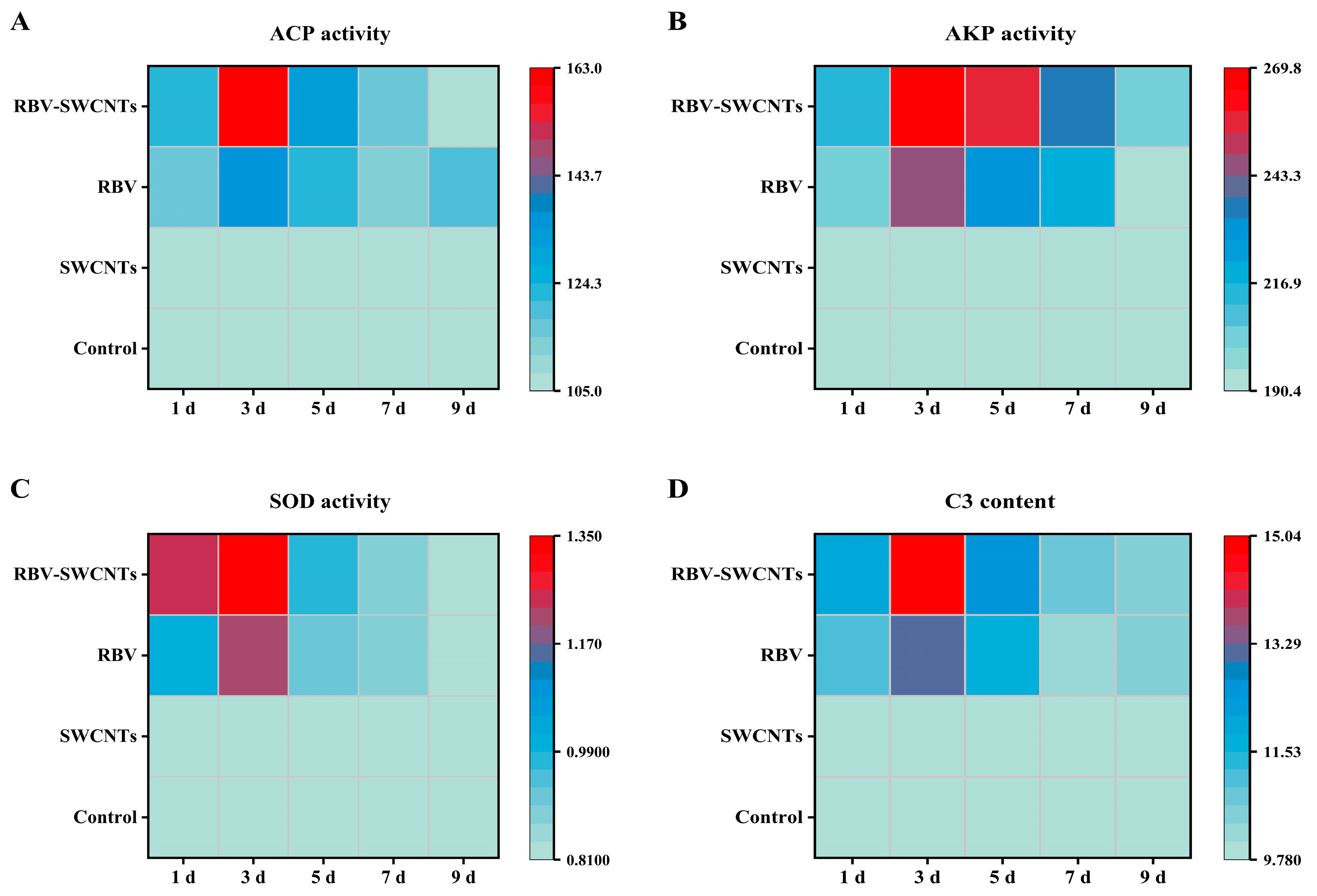
3.6. Evaluation of Non-Specific Immune Responses
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- De, R.; Mahata, M.K.; Kim, K.T. Structure-Based Varieties of Polymeric Nanocarriers and Influences of Their Physicochemical Properties on Drug Delivery Profiles. Adv. Sci. 2022, 9, e2105373. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Yang, J.; Asakura, Y.; Shuai, Q.; Yamauchi, Y. Nanoarchitectonics of Hollow Covalent Organic Frameworks: Synthesis and Applications. ACS Nano 2023, 17, 8918–8934. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.M.; Filipczak, N.; Torchilin, V.P. Cell penetrating peptides: A versatile vector for co-delivery of drug and genes in cancer. J. Control. Release 2021, 330, 1220–1228. [Google Scholar] [CrossRef]
- Zhu, Z.; Wang, J.; Pei, X.; Chen, J.; Wei, X.; Liu, Y.; Xia, P.; Wan, Q.; Gu, Z.; He, Y. Blue-ringed octopus-inspired microneedle patch for robust tissue surface adhesion and active injection drug delivery. Sci. Adv. 2023, 9, eadh2213. [Google Scholar] [CrossRef]
- Liu, J.; Cabral, H.; Mi, P. Nanocarriers address intracellular barriers for efficient drug delivery, overcoming drug resistance, subcellular targeting and controlled release. Adv. Drug Deliv. Rev. 2024, 207, 115239. [Google Scholar] [CrossRef] [PubMed]
- Imran, M.; Gowd, V.; Saha, P.; Rashid, S.; Ahmad Chaudhary, A.; Mohamed, M.Y.A.; Alawam, A.S.; Khan, R. Biologically inspired stealth—Camouflaged strategies in nanotechnology for the improved therapies in various diseases. Int. J. Pharm. 2023, 631, 122407. [Google Scholar] [CrossRef]
- Waheed, S.; Li, Z.; Zhang, F.; Chiarini, A.; Armato, U.; Wu, J. Engineering nano-drug biointerface to overcome biological barriers toward precision drug delivery. J. Nanobiotechnol. 2022, 20, 395. [Google Scholar] [CrossRef] [PubMed]
- Xia, K.; Yatabe, T.; Yonesato, K.; Kikkawa, S.; Yamazoe, S.; Nakata, A.; Ishikawa, R.; Shibata, N.; Ikuhara, Y.; Yamaguchi, K.; et al. Ultra-stable and highly reactive colloidal gold nanoparticle catalysts protected using multi-dentate metal oxide nanoclusters. Nat. Commun. 2024, 15, 851. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Song, X.; Liu, Y.; Fu, Y.; Ye, L.; Wang, N.; Wang, F.; Li, L.; Mohammadniaei, M.; Zhang, M.; et al. Synthesis of graphene quantum dots and their applications in drug delivery. J. Nanobiotechnol. 2020, 18, 142. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Xiong, Y.; Zhang, C.; Jia, Y.J.; Qiu, D.K.; Wang, G.X.; Zhu, B. Optimization of the efficacy of a SWCNTs-based subunit vaccine against infectious spleen and kidney necrosis virus in mandarin fish. Fish Shellfish Immunol. 2020, 106, 190–196. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.; Liu, G.L.; Ling, F.; Song, L.S.; Wang, G.X. Development toxicity of functionalized carbon nanotubes on rare minnow embryos and larvae. Nanotoxicology 2015, 9, 579–590. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Zhu, B.; Huang, A.; Hu, Y.; Wang, G.; Ling, F. Toxicological effects of multi-walled carbon nanotubes on Saccharomyces cerevisiae: The uptake kinetics and mechanisms and the toxic responses. J. Hazard. Mater. 2016, 318, 650–662. [Google Scholar] [CrossRef]
- Yu, X.B.; Chen, X.H.; Ling, F.; Hao, K.; Wang, G.X.; Zhu, B. Moroxydine hydrochloride inhibits grass carp reovirus replication and suppresses apoptosis in Ctenopharyngodon idella kidney cells. Antivir. Res. 2016, 131, 156–165. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Luo, F.; Li, J.; Zhu, B.; Wang, G.X. Biocompatibility assessment of single-walled carbon nanotubes using Saccharomyces cerevisiae as a model organism. J. Nanobiotechnol. 2018, 16, 44. [Google Scholar] [CrossRef]
- Zhu, S.; Huang, A.G.; Luo, F.; Li, J.; Li, J.; Zhu, L.; Zhao, L.; Zhu, B.; Ling, F.; Wang, G.X. Application of virus targeting nanocarrier drug delivery system in virus-induced central nervous system disease treatment. ACS Appl. Mater. Interfaces 2019, 11, 19006–19016. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Wang, G.X.; Zhu, B. Application of antigen presenting cell-targeted nanovaccine delivery system in rhabdovirus disease prophylactics using fish as a model organism. J. Nanobiotechnol. 2020, 18, 24. [Google Scholar] [CrossRef] [PubMed]
- Fisher, C.E.; Knudsen, J.L.; Lease, E.D.; Jerome, K.R.; Rakita, R.M.; Boeckh, M.; Limaye, A.P. Risk factors and outcomes of ganciclovir-resistant cytomegalovirus infection in solid organ transplant recipients. Clin. Infect. Dis. 2017, 65, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Meister, T.L.; Brüggemann, Y.; Nocke, M.K.; Ulrich, R.G.; Schuhenn, J.; Sutter, K.; Gömer, A.; Bader, V.; Winklhofer, K.F.; Broering, R.; et al. A ribavirin-induced ORF2 single-nucleotide variant produces defective hepatitis E virus particles with immune decoy function. Proc. Natl. Acad. Sci. USA 2022, 119, e2202653119. [Google Scholar] [CrossRef]
- Pfeiffer, J.K.; Kirkegaard, K. A single mutation in poliovirus RNA-dependent RNA polymerase confers resistance to mutagenic nucleotide analogs via increased fidelity. Proc. Natl. Acad. Sci. USA 2003, 100, 7289–7294. [Google Scholar] [CrossRef] [PubMed]
- Leyssen, P.; Balzarini, J.; De Clercq, E.; Neyts, J. The predominant mechanism by which ribavirin exerts its antiviral activity invitro against flaviviruses and paramyxoviruses is mediated by inhibition of IMP dehydrogenase. J. Virol. 2005, 79, 1943–1947. [Google Scholar] [CrossRef] [PubMed]
- Wray, S.K.; Gilbert, B.E.; Noall, M.W.; Knight, V. Mode of action of ribavirin: Effect of nucleotide pool alterations on influenza virus ribonucleoprotein synthesis. Antivir. Res. 1985, 5, 29–37. [Google Scholar] [CrossRef] [PubMed]
- McCormick, J.B.; King, I.J.; Webb, P.A.; Scribner, C.L.; Craven, R.B.; Johnson, K.M.; Elliott, L.H.; Belmont-Williams, R. Lassa fever. Eff. Ther. Ribavirin. N. Engl. J. Med. 1986, 314, 20–26. [Google Scholar] [CrossRef]
- Stein, D.S.; Creticos, C.M.; Jackson, G.G.; Bernstein, J.M.; Hayden, F.G.; Schiff, G.M.; Bernstein, D.I. Oral ribavirin treatment of influenza A and B. Antimicrob. Agents Chemother. 1987, 31, 1285–1287. [Google Scholar] [CrossRef]
- Graci, J.D.; Cameron, C.E. Mechanisms of action of ribavirin against distinct viruses. Rev. Med. Virol. 2006, 16, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-Olivencia, G.; Estébanez, M.; Membrillo, F.J.; Ybarra, M.D.C. Use of ribavirin in viruses other than hepatitis, C. A review of the evidence. Enfermedades Infecc. Microbiol. Clin. 2019, 37, 602–608. [Google Scholar] [CrossRef] [PubMed]
- Takayama-Ito, M.; Saijo, M. Antiviral drugs against severe fever with thrombocytopenia syndrome virus infection. Front. Microbiol. 2020, 11, 150. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Shen, Y.; Li, B.; Wang, G.X.; Zhu, B. Evaluation on the antiviral activity of ribavirin against infectious hematopoietic necrosis virus in epithelioma papulosum cyprini cells. Virus Res. 2019, 263, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Song, K.; Zhang, Z.; Chen, C.; Wang, G.; Yao, J.; Ling, F. Evaluation on the antiviral activity of ribavirin against Micropterus salmoides rhabdovirus (MSRV) in vitro and in vivo. Aquaculture 2021, 543, 736975. [Google Scholar] [CrossRef]
- Baek, E.J.; Kim, M.J.; Kim, K.I. In vitro and in vivo evaluation of the antiviral activity of arctigenin, ribavirin, and ivermectin against viral hemorrhagic septicemia virus infection. Fish Shellfish Immunol. 2023, 132, 108456. [Google Scholar] [CrossRef]
- Liu, J.; Zhao, D.; He, W.; Zhang, H.; Li, Z.; Luan, Y. Nanoassemblies from amphiphilic cytarabine prodrug for leukemia targeted therapy. J. Colloid Interface Sci. 2017, 487, 239–249. [Google Scholar] [CrossRef] [PubMed]
- Geethakumari, D.; Bhaskaran Sathyabhama, A.; Raji Sathyan, K.; Mohandas, D.; Somasekharan, J.V.; Thavarool Puthiyedathu, S. Folate functionalized chitosan nanoparticles as targeted delivery systems for improved anticancer efficiency of cytarabine in MCF-7 human breast cancer cell lines. Int. J. Biol. Macromol. 2022, 199, 150–161. [Google Scholar] [CrossRef] [PubMed]
- Kripp, M.; Hofheinz, R.D. Treatment of lymphomatous and leukemic meningitis with liposomal encapsulated cytarabine. Int. J. Nanomed. 2008, 3, 397–401. [Google Scholar]
- Ping, L.; Ruxian, J.; Mengping, Z.; Pei, J.; Zhuoya, L.; Guosheng, L.; Zhenyu, W.; Hailei, W. Whole-cell biosynthesis of cytarabine by an unnecessary protein-reduced Escherichia coli that coexpresses purine and uracil phosphorylase. Biotechnol. Bioeng. 2022, 119, 1768–1780. [Google Scholar] [CrossRef]
- Chen, F.; Xu, H.; Liu, J.; Cui, Y.; Luo, X.; Zhou, Y.; Chen, Q.; Jiang, L. Efficacy and safety of nucleoside antiviral drugs for treatment of recurrent herpes labialis: A systematic review and meta-analysis. J. Oral Pathol. Med. 2017, 46, 561–568. [Google Scholar] [CrossRef] [PubMed]
- Quenelle, D.C.; Birkmann, A.; Goldner, T.; Pfaff, T.; Zimmermann, H.; Bonsmann, S.; Collins, D.J.; Rice, T.L.; Prichard, M.N. Efficacy of pritelivir and acyclovir in the treatment of herpes simplex virus infections in a mouse model of herpes simplex encephalitis. Antivir. Res. 2018, 149, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Elion, G.B.; Furman, P.A.; Fyfe, J.A.; de Miranda, P.; Beauchamp, L.; Schaeffer, H.J. Selectivity of action of an antiherpetic agent, 9-(2-hydroxyethoxymethyl) guanine. Proc. Natl. Acad. Sci. USA 1977, 74, 5716–5720. [Google Scholar] [CrossRef] [PubMed]
- Miller, W.H.; Miller, R.L. Phosphorylation of acyclovir (acycloguanosine) monophosphate by GMP kinase. J. Biol. Chem. 1980, 255, 7204–7207. [Google Scholar] [CrossRef] [PubMed]
- Quijano Cardé, E.M.; Yazdi, Z.; Yun, S.; Hu, R.; Knych, H.; Imai, D.M.; Soto, E. Pharmacokinetic and efficacy study of acyclovir against cyprinid herpesvirus 3 in cyprinus carpio. Front. Vet. Sci. 2020, 7, 587952. [Google Scholar] [CrossRef]
- Hao, K.; Yuan, S.; Yu, F.; Chen, X.H.; Bian, W.J.; Feng, Y.H.; Zhao, Z. Acyclovir inhibits channel catfish virus replication and protects channel catfish ovary cells from apoptosis. Virus Res. 2021, 292, 198249. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.F.; He, J.; Zeng, R.Y.; Li, Z.M.; Luo, Z.Y.; Pan, W.Q.; He, J.G. Deletion of the infectious spleen and kidney necrosis virus ORF069L reduces virulence to mandarin fish Siniperca chuatsi. Fish Shellfish Immunol. 2019, 95, 328–335. [Google Scholar] [CrossRef]
- He, J.H.; Xia, Q.; Weng, S.; He, J.; Xu, X. Identification of infectious spleen and kidney necrosis virus (ISKNV)-encoded microRNAs. Virus Genes 2020, 56, 724–733. [Google Scholar] [CrossRef] [PubMed]
- Chinchar, V.G.; Hick, P.; Ince, I.A.; Jancovich, J.K.; Marschang, R.; Qin, Q.; ICTV Report Consortium. ICTV virus taxonomy profile: Iridoviridae. J. Gen. Virol. 2017, 98, 890–891. [Google Scholar] [CrossRef] [PubMed]
- Fusianto, C.; Hick, P.M.; Becker, J.A. Stability of infectious spleen and kidney necrosis virus and susceptibility to physical and chemical disinfectants. Aquaculture 2019, 506, 104–111. [Google Scholar] [CrossRef]
- Zhu, Z.M.; Duan, C.; Li, Y.; Huang, C.L.; Weng, S.P.; He, J.G.; Dong, C.F. Pathogenicity and histopathology of infectious spleen and kidney necrosis virus genotype II (ISKNV-II) recovering from mass mortality of farmed Asian seabass, Lates calcarifer, in southern China. Aquaculture 2020, 534, 736326. [Google Scholar] [CrossRef]
- Lin, Q.; Fu, X.; Liu, L.; Liang, H.; Guo, H.; Yin, S.; Li, N. Application and development of a TaqMan real-time PCR for detecting infectious spleen and kidney necrosis virus in Siniperca chuatsi. Microb. Pathog. 2017, 107, 98–105. [Google Scholar] [CrossRef]
- Zhu, B.; Liu, G.L.; Ling, F.; Wang, G.X. Carbon nanotube-based nanocarrier loaded with ribavirin against grass carp reovirus. Antivir. Res. 2015, 118, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Pan, C.; Xu, S.; Hu, L.; Su, X.; Ou, J.; Zou, H.; Guo, Z.; Zhang, Y.; Guo, B. Using oxidized carbon nanotubes as matrix for analysis of small molecules by MALDI-TOF MS. J. Am. Soc. Mass Spectrom. 2005, 16, 883–892. [Google Scholar] [CrossRef]
- Zhang, F.; Lu, G.; Liu, J.; Yan, Z.; Zhang, Z. Bioaccumulation, distribution and metabolism of BDE-153 in the freshwater fish Carassius auratus after dietary exposure. Ecotoxicol. Environ. Saf. 2014, 108, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Rigo-Bonnin, R.; Padullés, A.; Corral-Comesaña, S.; Cerezo, G.; Grinyó, J.M.; Colom, H.; Alía-Ramos, P.; Lloberas, N. Measurement of ganciclovir concentration in human plasma by ultra-performance liquid chromatography-tandem mass spectrometry. Clin. Chim. Acta 2014, 427, 58–64. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−△△CT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- FAO. The State of World Fisheries and Aquaculture; Food and Agriculture Oraganization of the United Nations: Rome, Italy, 2014. [Google Scholar]
- Embregts, C.W.; Forlenza, M. Oral vaccination of fish: Lessons from humans and veterinary species. Dev. Comp. Immunol. 2016, 64, 118–137. [Google Scholar] [CrossRef]
- Danhier, F.; Ansorena, E.; Silva, J.M.; Coco, R.; Le Breton, A.; Préat, V. PLGA-based nanoparticles: An overview of biomedical applications. J. Control. Release 2012, 161, 505–522. [Google Scholar] [CrossRef] [PubMed]
- Sadeghi, I.; Byrne, J.; Shakur, R.; Langer, R. Engineered drug delivery devices to address Global Health challenges. J. Control. Release 2021, 331, 503–514. [Google Scholar] [CrossRef] [PubMed]
- Pereiro, P.; Figueras, A.; Novoa, B. Compilation of antiviral treatments and strategies to fight fish viruses. Rev. Aquac. 2021, 13, 1223–1254. [Google Scholar] [CrossRef]
- Tate, P.M.; Mastrodomenico, V.; Mounce, B.C. Ribavirin induces polyamine depletion via nucleotide depletion to limit virus replication. Cell Rep. 2019, 28, 2620–2633.e4. [Google Scholar] [CrossRef] [PubMed]
- Aljabr, W.; Touzelet, O.; Pollakis, G.; Wu, W.; Munday, D.C.; Hughes, M.; Hertz-Fowler, C.; Kenny, J.; Fearns, R.; Barr, J.N.; et al. Investigating the influence of ribavirin on human respiratory syncytial virus rna synthesis by using a high-resolution transcriptome sequencing approach. J. Virol. 2016, 90, 4876–4888. [Google Scholar] [CrossRef] [PubMed]
- Ying, C.; De Clercq, E.; Neyts, J. Ribavirin and mycophenolic acid potentiate the activity of guanine- and diaminopurine-based nucleoside analogues against hepatitis B virus. Antivir. Res. 2000, 48, 117–124. [Google Scholar] [CrossRef]
- Lertwanakarn, T.; Trongwongsa, P.; Yingsakmongkol, S.; Khemthong, M.; Tattiyapong, P.; Surachetpong, W. Antiviral activity of ribavirin against tilapia tilapinevirus in fish cells. Pathogens 2021, 10, 1616. [Google Scholar] [CrossRef]
- Rivas-Aravena, A.; Vallejos-Vidal, E.; Cortez-San Martin, M.; Reyes-Lopez, F.; Tello, M.; Mora, P.; Sandino, A.M.; Spencer, E. Inhibitory effect of a nucleotide analog on infectious salmon anemia virus infection. J. Virol. 2011, 85, 8037–8045. [Google Scholar] [CrossRef]
- Troszok, A.; Kolek, L.; Szczygieł, J.; Wawrzeczko, J.; Borzym, E.; Reichert, M.; Kamińska, T.; Ostrowski, T.; Jurecka, P.; Adamek, M.; et al. Acyclovir inhibits Cyprinid herpesvirus 3 multiplication in vitro. J. Fish Dis. 2018, 41, 1709–1718. [Google Scholar] [CrossRef]
- Nasr-Eldahan, S.; Nabil-Adam, A.; Shreadah, M.A.; Maher, A.M.; El-Sayed Ali, T. A review article on nanotechnology in aquaculture sustainability as a novel tool in fish disease control. Aquac. Int. 2021, 29, 1459–1480. [Google Scholar] [CrossRef] [PubMed]
- Sahoo, N.G.; Bao, H.; Pan, Y.; Pal, M.; Kakran, M.; Cheng, H.K.; Li, L.; Tan, L.P. Functionalized carbon nanomaterials as nanocarriers for loading and delivery of a poorly water-soluble anticancer drug: A comparative study. Chem. Commun. 2011, 47, 5235–5237. [Google Scholar] [CrossRef]
- Guven, A.; Villares, G.J.; Hilsenbeck, S.G.; Lewis, A.; Landua, J.D.; Dobrolecki, L.E.; Wilson, L.J.; Lewis, M.T. Carbon nanotube capsules enhance the in vivo efficacy of cisplatin. Acta Biomater. 2017, 58, 466–478. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Luo, F.; Zhu, B.; Ling, F.; Wang, E.L.; Liu, T.Q.; Wang, G.X. A nanobody-mediated virus-targeting drug delivery platform for the central nervous system viral disease therapy. Microbiol. Spectr. 2021, 9, e0148721. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Accession No. | Primer Sequences (from 5′ to 3′) | Product Size (bp) | |
|---|---|---|---|---|
| β-Actin | AY885683 | Forward | ATTTCCGACACGGAGAGG | 199 |
| Reverse | CATGGGTTTAGGATACGCTC | |||
| MHC I | AY391782 | Forward | CCTGGCAGAAAAATGGACAAG | 271 |
| Reverse | CCAACAACACCAATGACAATC | |||
| IRF1 | JN859505.1 | Forward | CAGAGCAGCAAGATTTCCCATT | 72 |
| Reverse | CCCCGATCTCAACTGACAAAG | |||
| Mx | AY392097 | Forward | TGTTCCAGTGTCTGCTGATTGA | 345 |
| Reverse | GACAGTCCAGGAAACGCATATA | |||
| Viperin | AY395718 | Forward | CTCCTGCTTTGCCAGAGATTT | 201 |
| Reverse | CAGTTGCCAGGAGTTGTATGTT | |||
| I-IFN | AB376666.1 | Forward | CAGAGTCAATGCTCCGCTT | 297 |
| Reverse | CTCAGATGACTGCCGTTGC | |||
| TNF-α | AJ311800.2 | Forward | TGTGCCGCCGCTGTCTGCTTCACGCT | 291 |
| Reverse | GATGAGGAAAGACACCTGGCTGTAGA | |||
| ISKNV-MCP | HQ317465.1 | Forward | TGCCCATTCCCCTCTTC | 192 |
| Reverse | CGCTCCTCGCTTGTCAGTA | |||
| Concentration (μg/mL) | Recovery (%) | Precision (%) (Coefficients of Variation) | |
|---|---|---|---|
| Intra-Day (n = 5) | Inter-Day (n = 5) | ||
| 60 | 92.46 ± 1.570 | 1.81 | 3.18 |
| 80 | 95.90 ± 2.287 | 1.78 | 2.57 |
| 100 | 93.05 ± 1.915 | 1.25 | 2.58 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jia, Y.; Zhao, Z.; Chen, L.; Liu, Y.; Zhu, B. Carbon Nanotube-Based Drug Delivery System Increases Drug Content and Promotes Immune Response in Mandarin Fish. Fishes 2025, 10, 92. https://doi.org/10.3390/fishes10030092
Jia Y, Zhao Z, Chen L, Liu Y, Zhu B. Carbon Nanotube-Based Drug Delivery System Increases Drug Content and Promotes Immune Response in Mandarin Fish. Fishes. 2025; 10(3):92. https://doi.org/10.3390/fishes10030092
Chicago/Turabian StyleJia, Yijun, Zhao Zhao, Leyang Chen, Yongqi Liu, and Bin Zhu. 2025. "Carbon Nanotube-Based Drug Delivery System Increases Drug Content and Promotes Immune Response in Mandarin Fish" Fishes 10, no. 3: 92. https://doi.org/10.3390/fishes10030092
APA StyleJia, Y., Zhao, Z., Chen, L., Liu, Y., & Zhu, B. (2025). Carbon Nanotube-Based Drug Delivery System Increases Drug Content and Promotes Immune Response in Mandarin Fish. Fishes, 10(3), 92. https://doi.org/10.3390/fishes10030092