Application of microRNA Database Mining in Biomarker Discovery and Identification of Therapeutic Targets for Complex Disease



Abstract
1. Introduction
2. Materials and Methods
2.1. Human microRNA Disease Database Version 3.2
2.2. Prediction of hsa-miRNA-21 Targets & Functional Analyses
3. Results
4. Discussion and Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, P.; Wu, W.; Chen, Q.; Chen, M. Non-Coding RNAs and their Integrated Networks. J. Integr. Bioinform 2019, 16. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.C.; Feinbaum, R.L.; Ambros, V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell 1993, 75, 843–854. [Google Scholar] [CrossRef]
- Reinhart, B.J.; Slack, F.J.; Basson, M.; Pasquinelli, A.E.; Bettinger, J.C.; Rougvie, A.E.; Horvitz, H.R.; Ruvkun, G. The 21-nucleotide let-7 RNA regulates developmental timing in Caenorhabditis elegans. Nature 2000, 403, 901–906. [Google Scholar] [CrossRef] [PubMed]
- Ardekani, A.M.; Naeini, M.M. The Role of MicroRNAs in Human Diseases. Avicenna J. Med. Biotechnol. 2010, 2, 161–179. [Google Scholar] [PubMed]
- Rupaimoole, R.; Slack, F.J. MicroRNA therapeutics: Towards a new era for the management of cancer and other diseases. Nat. Rev. Drug Discov. 2017, 16, 203–222. [Google Scholar] [CrossRef]
- Ha, M.; Kim, V.N. Regulation of microRNA biogenesis. Nat. Rev. Mol. Cell Biol. 2014, 15, 509–524. [Google Scholar] [CrossRef]
- Liu, H.; Lei, C.; He, Q.; Pan, Z.; Xiao, D.; Tao, Y. Nuclear functions of mammalian MicroRNAs in gene regulation, immunity and cancer. Mol. Cancer 2018, 17, 64. [Google Scholar] [CrossRef]
- Laterza, O.F.; Lim, L.; Garrett-Engele, P.W.; Vlasakova, K.; Muniappa, N.; Tanaka, W.K.; Johnson, J.M.; Sina, J.F.; Fare, T.L.; Sistare, F.D.; et al. Plasma MicroRNAs as sensitive and specific biomarkers of tissue injury. Clin. Chem. 2009, 55, 1977–1983. [Google Scholar] [CrossRef]
- WHO. Cardiovascular Diseases (CVDs); World Health Organization (WHO): Geneva, Switzerland, 2017. [Google Scholar]
- Kalogeris, T.; Baines, C.P.; Krenz, M.; Korthuis, R.J. Cell biology of ischemia/reperfusion injury. Int. Rev. Cell Mol. Biol. 2012, 298, 229–317. [Google Scholar] [CrossRef]
- Hearse, D.J.; Humphrey, S.M.; Chain, E.B. Abrupt reoxygenation of the anoxic potassium-arrested perfused rat heart: A study of myocardial enzyme release. J. Mol. Cell Cardiol. 1973, 5, 395–407. [Google Scholar] [CrossRef]
- Granger, D.N.; Kvietys, P.R. Reperfusion injury and reactive oxygen species: The evolution of a concept. Redox Biol. 2015, 6, 524–551. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Shi, J.; Gao, Y.; Cui, C.; Zhang, S.; Li, J.; Zhou, Y.; Cui, Q. HMDD v3.0: A database for experimentally supported human microRNA-disease associations. Nucleic Acids Res. 2019, 47, D1013–D1017. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Qiu, C.; Tu, J.; Geng, B.; Yang, J.; Jiang, T.; Cui, Q. HMDD v2.0: A database for experimentally supported human microRNA and disease associations. Nucleic Acids Res. 2014, 42, D1070–D1074. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.; Zhang, Q.; Deng, M.; Miao, J.; Guo, Y.; Gao, W.; Cui, Q. An analysis of human microRNA and disease associations. PLoS ONE 2008, 3, e3420. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, X. miRDB: An online database for prediction of functional microRNA targets. Nucleic Acids Res. 2020, 48, D127–D131. [Google Scholar] [CrossRef]
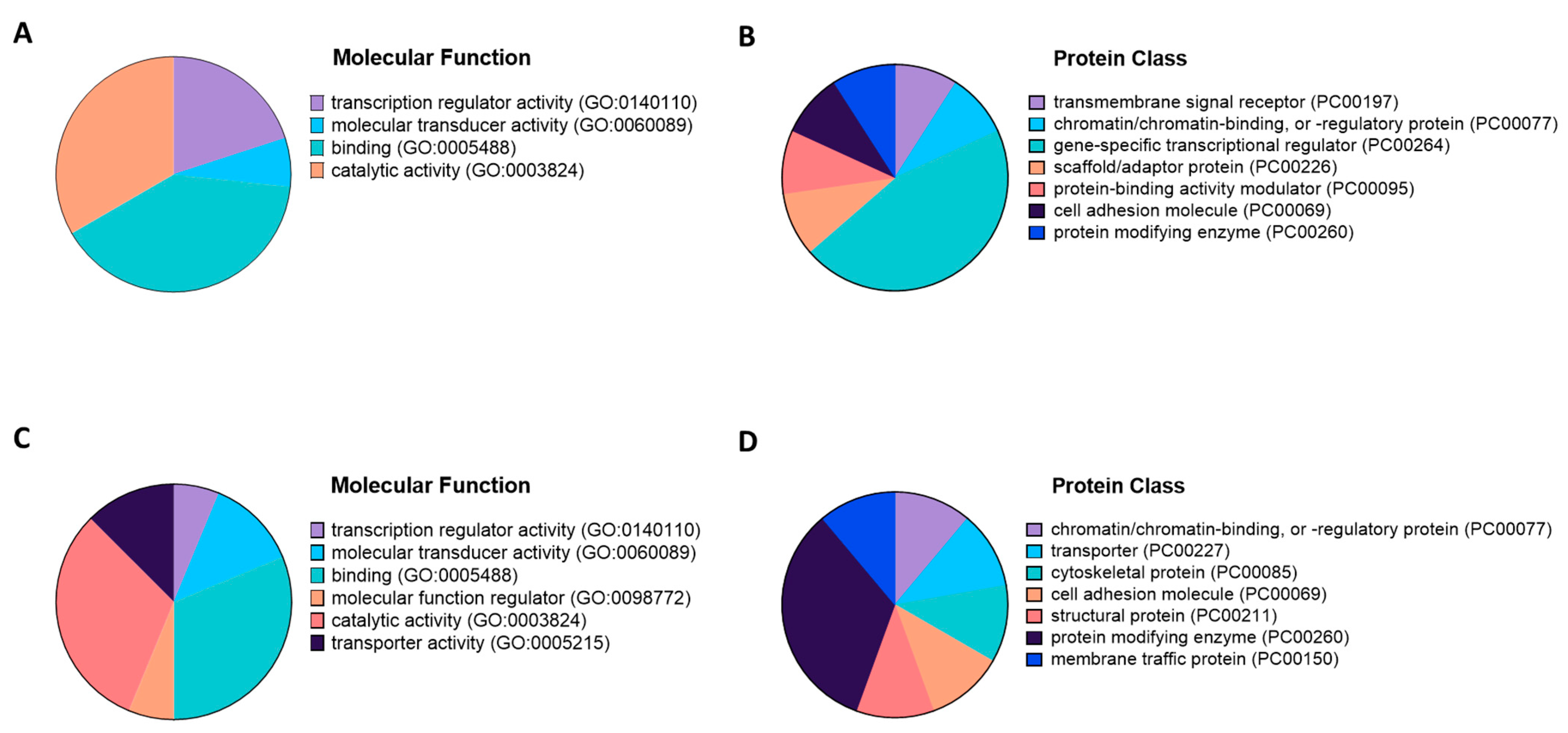
- The-Gene-Ontology-Consortium. Expansion of the Gene Ontology knowledgebase and resources. Nucleic Acids Res. 2017, 45, D331–D338. [Google Scholar] [CrossRef]
- Thomas, P.D.; Campbell, M.J.; Kejariwal, A.; Mi, H.; Karlak, B.; Daverman, R.; Diemer, K.; Muruganujan, A.; Narechania, A. PANTHER: A library of protein families and subfamilies indexed by function. Genome Res. 2003, 13, 2129–2141. [Google Scholar] [CrossRef]
- Mi, H.; Muruganujan, A.; Huang, X.; Ebert, D.; Mills, C.; Guo, X.; Thomas, P.D. Protocol Update for large-scale genome and gene function analysis with the PANTHER classification system (v.14.0). Nat. Protoc. 2019, 14, 703–721. [Google Scholar] [CrossRef]
- Krämer, A.; Green, J.; Pollard, J., Jr.; Tugendreich, S. Causal analysis approaches in Ingenuity Pathway Analysis. Bioinformatics 2014, 30, 523–530. [Google Scholar] [CrossRef]
- Cheng, Y.; Zhang, C. MicroRNA-21 in cardiovascular disease. J. Cardiovasc Transl. Res. 2010, 3, 251–255. [Google Scholar] [CrossRef]
- Coelho, M.; Oliveira, T.; Fernandes, R. Biochemistry of adipose tissue: An endocrine organ. Arch. Med. Sci. 2013, 9, 191–200. [Google Scholar] [CrossRef]
- Zhang, J.; Xing, Q.; Zhou, X.; Li, J.; Li, Y.; Zhang, L.; Zhou, Q.; Tang, B. Circulating miRNA-21 is a promising biomarker for heart failure. Mol. Med. Rep. 2017, 16, 7766–7774. [Google Scholar] [CrossRef]
- Yin, C.; Salloum, F.N.; Kukreja, R.C. A novel role of microRNA in late preconditioning: Upregulation of endothelial nitric oxide synthase and heat shock protein 70. Circ. Res. 2009, 104, 572–575. [Google Scholar] [CrossRef] [PubMed]
- Yin, C.; Wang, X.; Kukreja, R.C. Endogenous microRNAs induced by heat-shock reduce myocardial infarction following ischemia-reperfusion in mice. FEBS Lett. 2008, 582, 4137–4142. [Google Scholar] [CrossRef] [PubMed]
- Ji, R.; Cheng, Y.; Yue, J.; Yang, J.; Liu, X.; Chen, H.; Dean, D.B.; Zhang, C. MicroRNA expression signature and antisense-mediated depletion reveal an essential role of MicroRNA in vascular neointimal lesion formation. Circ. Res. 2007, 100, 1579–1588. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Ji, R.; Yue, J.; Yang, J.; Liu, X.; Chen, H.; Dean, D.B.; Zhang, C. MicroRNAs are aberrantly expressed in hypertrophic heart: Do they play a role in cardiac hypertrophy? Am. J. Pathol. 2007, 170, 1831–1840. [Google Scholar] [CrossRef] [PubMed]
- Thum, T.; Gross, C.; Fiedler, J.; Fischer, T.; Kissler, S.; Bussen, M.; Galuppo, P.; Just, S.; Rottbauer, W.; Frantz, S.; et al. MicroRNA-21 contributes to myocardial disease by stimulating MAP kinase signalling in fibroblasts. Nature 2008, 456, 980–984. [Google Scholar] [CrossRef]
- Yong, K.; Dogra, G.; Boudville, N.; Chan, D.; Adams, L.; Ching, H.; Lim, E.M.; Lim, W.H. Interleukin-12 is associated with arterial stiffness in healthy individuals. Am. J. Hypertens 2013, 26, 159–162. [Google Scholar] [CrossRef]
- Gryshkova, V.; Fleming, A.; McGhan, P.; De Ron, P.; Fleurance, R.; Valentin, J.P.; Nogueira da Costa, A. miR-21-5p as a potential biomarker of inflammatory infiltration in the heart upon acute drug-induced cardiac injury in rats. Toxicol. Lett. 2018, 286, 31–38. [Google Scholar] [CrossRef]
- Liu, W.; Zi, M.; Jin, J.; Prehar, S.; Oceandy, D.; Kimura, T.E.; Lei, M.; Neyses, L.; Weston, A.H.; Cartwright, E.J.; et al. Cardiac-specific deletion of mkk4 reveals its role in pathological hypertrophic remodeling but not in physiological cardiac growth. Circ. Res. 2009, 104, 905–914. [Google Scholar] [CrossRef]
- Minamino, T.; Yujiri, T.; Terada, N.; Taffet, G.E.; Michael, L.H.; Johnson, G.L.; Schneider, M.D. MEKK1 is essential for cardiac hypertrophy and dysfunction induced by Gq. Proc. Natl. Acad. Sci. USA 2002, 99, 3866–3871. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Jin, P.; Liu, J.; Xie, X. Exosomal microRNA-122 mediates obesity-related cardiomyopathy through suppressing mitochondrial ADP-ribosylation factor-like 2. Clin. Sci. (Lond.) 2019, 133, 1871–1881. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Ha, T.; Liu, L.; Zou, J.; Zhang, X.; Kalbfleisch, J.; Gao, X.; Williams, D.; Li, C. Increased expression of microRNA-146a decreases myocardial ischaemia/reperfusion injury. Cardiovasc. Res. 2013, 97, 432–442. [Google Scholar] [CrossRef] [PubMed]
- Shen, L.; Li, C.; Zhang, H.; Qiu, S.; Fu, T.; Xu, Y. Downregulation of miR-146a Contributes to Cardiac Dysfunction Induced by the Tyrosine Kinase Inhibitor Sunitinib. Front. Pharmacol. 2019, 10, 914. [Google Scholar] [CrossRef]
- Barupal, J.K.; Saini, A.K.; Chand, T.; Meena, A.; Beniwal, S.; Suthar, J.R.; Meena, N.; Kachhwaha, S.; Kothari, S.L. ExcellmiRDB for translational genomics: A curated online resource for extracellular microRNAs. Omics 2015, 19, 24–30. [Google Scholar] [CrossRef]
- Girijadevi, R.; Sreedevi, V.C.; Sreedharan, J.V.; Pillai, M.R. IntmiR: A complete catalogue of intronic miRNAs of human and mouse. Bioinformation 2011, 5, 458–459. [Google Scholar] [CrossRef][Green Version]
- Jiang, Q.; Wang, Y.; Hao, Y.; Juan, L.; Teng, M.; Zhang, X.; Li, M.; Wang, G.; Liu, Y. miR2Disease: A manually curated database for microRNA deregulation in human disease. Nucleic Acids Res. 2009, 37, D98–D104. [Google Scholar] [CrossRef]
- Ruepp, A.; Kowarsch, A.; Theis, F. PhenomiR: microRNAs in human diseases and biological processes. Methods Mol. Biol. 2012, 822, 249–260. [Google Scholar] [CrossRef]
- Nalluri, J.J.; Barh, D.; Azevedo, V.; Ghosh, P. miRsig: A consensus-based network inference methodology to identify pan-cancer miRNA-miRNA interaction signatures. Sci. Rep. 2017, 7, 39684. [Google Scholar] [CrossRef]
- Hanna, J.; Hossain, G.S.; Kocerha, J. The Potential for microRNA Therapeutics and Clinical Research. Front. Genet. 2019, 10, 478. [Google Scholar] [CrossRef]
- Bonneau, E.; Neveu, B.; Kostantin, E.; Tsongalis, G.J.; De Guire, V. How close are miRNAs from clinical practice? A perspective on the diagnostic and therapeutic market. Ejifcc 2019, 30, 114–127. [Google Scholar] [PubMed]




{kind=link}
{kind=link}
{kind=link}
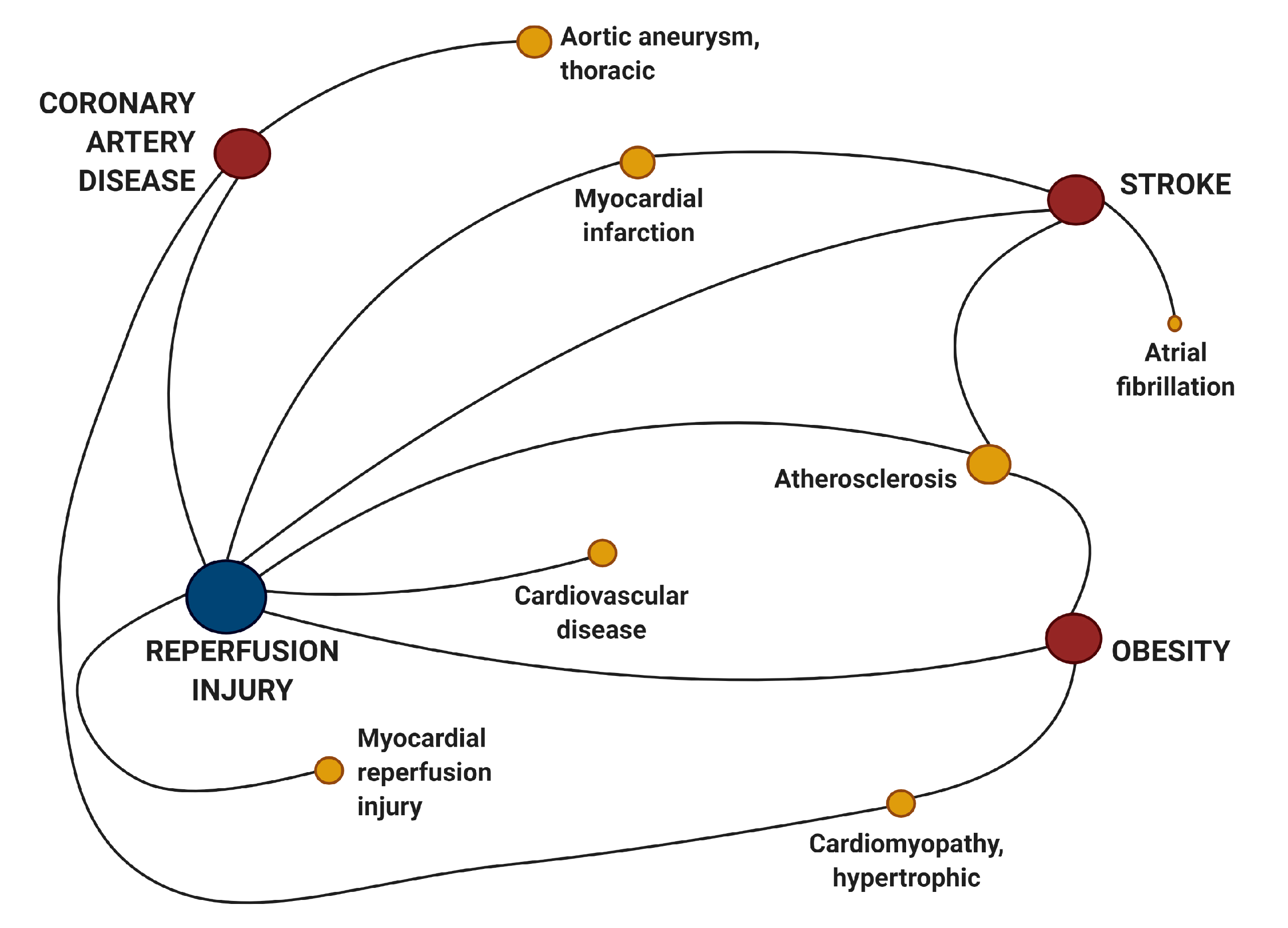
| Genetics I/R Injury | Targets CAD | hsa-miR-21 |
| hsa-miR-221 | ||
| Targets I/R injury | hsa-miR-146a | |
| hsa-miR-21 | ||
| hsa-miR-214 | ||
| hsa-miR-497 | ||
| Targets Stroke | hsa-miR-146a | |
| Genetics Obesity | Targets CAD | hsa-miR-21 |
| hsa-miR-222 | ||
| Targets Stroke | hsa-miR-155 | |
| Targets I/R injury | hsa-miR-21 |
| Target Rank | Target Score | miRNA Name | Gene Symbol | Gene Description |
|---|---|---|---|---|
| 1 | 99 | hsa-miR-21-5p | YOD1 | YOD1 deubiquitinase |
| 2 | 99 | hsa-miR-21-5p | FASLG | Fas ligand |
| 3 | 99 | hsa-miR-21-5p | PRDM11 | PR/SET domain 11 |
| 4 | 99 | hsa-miR-21-5p | VCL | vinculin |
| 5 | 99 | hsa-miR-21-5p | ZNF367 | zinc finger protein 367 |
| 6 | 98 | hsa-miR-21-5p | SKP2 | S-phase kinase associated protein 2 |
| 7 | 98 | hsa-miR-21-5p | TGFBI | transforming growth factor beta induced |
| 8 | 97 | hsa-miR-21-5p | IL12A | interleukin 12A |
| 9 | 97 | hsa-miR-21-5p | RAB6D | RAB6D, member RAS oncogene family |
| 10 | 97 | hsa-miR-21-5p | ADGRG2 | adhesion G protein-coupled receptor G2 |
| 11 | 97 | hsa-miR-21-5p | RALGPS2 | Ral GEF with PH domain and SH3 binding motif 2 |
| 12 | 97 | hsa-miR-21-5p | PLAG1 | PLAG1 zinc finger |
| 13 | 97 | hsa-miR-21-5p | RBPJ | recombination signal binding protein for IgK J region |
| 14 | 97 | hsa-miR-21-5p | PELI1 | pellino E3 ubiquitin protein ligase 1 |
| 15 | 97 | hsa-miR-21-5p | CREBRF | CREB3 regulatory factor |
| 16 | 97 | hsa-miR-21-5p | KRIT1 | KRIT1, ankyrin repeat containing |
| 17 | 96 | hsa-miR-21-5p | SCML2 | Scm polycomb group protein like 2 |
| 18 | 96 | hsa-miR-21-5p | RSAD2 | radical S-adenosyl methionine domain containing 2 |
| 19 | 96 | hsa-miR-21-5p | PBRM1 | polybromo 1 |
| 20 | 96 | hsa-miR-21-5p | GATAD2B | GATA zinc finger domain containing 2B |
| Target Rank | Target Score | miRNA Name | Gene Symbol | Gene Description |
|---|---|---|---|---|
| 1 | 99 | hsa-miR-21-3p | STK38L | serine/threonine kinase 38 like |
| 2 | 98 | hsa-miR-21-3p | PCDH19 | protocadherin 19 |
| 3 | 96 | hsa-miR-21-3p | LAMP1 | lysosomal associated membrane protein 1 |
| 4 | 96 | hsa-miR-21-3p | GRIA2 | glutamate ionotropic receptor AMPA type subunit 2 |
| 5 | 96 | hsa-miR-21-3p | TOGARAM1 | TOG array regulator of axonemal microtubules 1 |
| 6 | 96 | hsa-miR-21-3p | ATP1B1 | ATPase Na+/K+ transporting subunit beta 1 |
| 7 | 96 | hsa-miR-21-3p | TSC22D2 | TSC22 domain family member 2 |
| 8 | 96 | hsa-miR-21-3p | NAP1L5 | nucleosome assembly protein 1 like 5 |
| 9 | 95 | hsa-miR-21-3p | UBE4B | ubiquitination factor E4B |
| 10 | 95 | hsa-miR-21-3p | ZNF326 | zinc finger protein 326 |
| 11 | 95 | hsa-miR-21-3p | CDK8 | cyclin dependent kinase 8 |
| 12 | 94 | hsa-miR-21-3p | MAP2K4 | mitogen-activated protein kinase kinase 4 |
| 13 | 94 | hsa-miR-21-3p | AKAP11 | A-kinase anchoring protein 11 |
| 14 | 94 | hsa-miR-21-3p | GPM6A | glycoprotein M6A |
| 15 | 94 | hsa-miR-21-3p | MAP3K1 | mitogen-activated protein kinase kinase kinase 1 |
| 16 | 94 | hsa-miR-21-3p | PARD3B | par-3 family cell polarity regulator beta |
| 17 | 94 | hsa-miR-21-3p | ALCAM | activated leukocyte cell adhesion molecule |
| 18 | 94 | hsa-miR-21-3p | FOXO3 | forkhead box O3 |
| 19 | 94 | hsa-miR-21-3p | FYTTD1 | forty-two-three domain containing 1 |
| 20 | 94 | hsa-miR-21-3p | PHYHIPL | phytanoyl-CoA 2-hydroxylase interacting protein like |
| Canonical Pathways | Molecules |
|---|---|
| Crosstalk between Dendritic Cells and Natural Killer Cells | FASLG, IL12A |
| Altered T Cell and B Cell Signaling in Rheumatoid Arthritis | FASLG, IL12A |
| PD-1, PD-L1 cancer immunotherapy pathway | IL12A, SKP2 |
| Type I Diabetes Mellitus Signaling | FASLG, IL12A |
| Airway Pathology in Chronic Obstructive Pulmonary Disease | FASLG, IL12A |
| Role of Pattern Recognition Receptors in Recognition of Bacteria and Viruses | FASLG, IL12A |
| Necroptosis Signaling Pathway | FASLG, PELI1 |
| HMGB1 Signaling | FASLG, IL12A |
| Hepatic Cholestasis | FASLG, IL12A |
| Natural Killer Cell Signaling | FASLG, IL12A |
| Differential Regulation of Cytokine Production in Macrophages and T Helper Cells by IL-17A and IL-17F | IL12A |
| Role of Lipids/Lipid Rafts in the Pathogenesis of Influenza | RSAD2 |
| Differential Regulation of Cytokine Production in Intestinal Epithelial Cells by IL-17A and IL-17F | IL12A |
| Tumoricidal Function of Hepatic Natural Killer Cells | FASLG |
| Estrogen-mediated S-phase Entry | SKP2 |
| Systemic Lupus Erythematosus In B Cell Signaling Pathway | FASLG, IL12A |
| Neuroinflammation Signaling Pathway | FASLG, IL12A |
| Cytotoxic T Lymphocyte-mediated Apoptosis of Target Cells | FASLG |
| Notch Signaling | RBPJ |
| Antiproliferative Role of TOB in T Cell Signaling | SKP2 |
| Canonical Pathways | Molecules |
|---|---|
| EGF Signaling | MAP2K4, MAP3K1 |
| Toll-like Receptor Signaling | MAP2K4, MAP3K1 |
| NRF2-mediated Oxidative Stress Response | MAP2K4, MAP3K1 |
| Glutamate Receptor Signaling | GRIA2 |
| FGF Signaling | MAP3K1 |
| Apoptosis Signaling | MAP2K4 |
| p38 MAPK Signaling | MAP2K4 |
| Inhibition of Angiogenesis by TSP1 | MAP2K4 |
| Integrin Signaling | MAP2K4 |
| IGF-1 Signaling | FOXO3 |
| PI3K/AKT Signaling | FOXO3 |
| Huntington’s Disease Signaling | MAP2K4 |
| TNFR2 Signaling | MAP2K4, MAP3K1 |
| April Mediated Signaling | MAP2K4, MAP3K1 |
| B Cell Activating Factor Signaling | MAP2K4, MAP3K1 |
| TNFR1 Signaling | MAP2K4, MAP3K1 |
| CD27 Signaling in Lymphocytes | MAP2K4, MAP3K1 |
| Pyridoxal 5′-phosphate Salvage Pathway | CDK8, MAP2K4 |
| Ceramide Signaling | MAP2K4, MAP3K1 |
| RANK Signaling in Osteoclasts | MAP2K4, MAP3K1 |
| Category | Molecules |
|---|---|
| Cardiac Dilation | VCL |
| Cardiac Enlargement | FASLG, VCL |
| Heart Failure | RBPJ, VCL |
| Cardiac Hypoplasia | PBRM1, RBPJ |
| Cardiac Dysfunction | FASLG, RBPJ, VCL |
| Cardiac Arrhythmia | VCL |
| Cardiac Inflammation | RBPJ |
| Tachycardia | VCL |
| Category | Molecules |
|---|---|
| Cardiac Necrosis/Cell Death | FOXO3, MAP2K4, MAP3K1, UBE4B |
| Heart Failure | MAP2K4, MAP3K1, UBE4B |
| Cardiac Congestive Cardiac Failure | MAP3K1, UBE4B |
| Cardiac Enlargement | FOXO3, MAP2K4, UBE4B |
| Cardiac Arrhythmia | ATP1B1, FOXO3 |
| Cardiac Inflammation | MAP3K1 |
| Cardiac Infarction | FOXO3 |
| Cardiac Dilation | FOXO3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Major, J.L.; Bagchi, R.A.; Pires da Silva, J. Application of microRNA Database Mining in Biomarker Discovery and Identification of Therapeutic Targets for Complex Disease. Methods Protoc. 2021, 4, 5. https://doi.org/10.3390/mps4010005
Major JL, Bagchi RA, Pires da Silva J. Application of microRNA Database Mining in Biomarker Discovery and Identification of Therapeutic Targets for Complex Disease. Methods and Protocols. 2021; 4(1):5. https://doi.org/10.3390/mps4010005
Chicago/Turabian StyleMajor, Jennifer L., Rushita A. Bagchi, and Julie Pires da Silva. 2021. "Application of microRNA Database Mining in Biomarker Discovery and Identification of Therapeutic Targets for Complex Disease" Methods and Protocols 4, no. 1: 5. https://doi.org/10.3390/mps4010005
APA StyleMajor, J. L., Bagchi, R. A., & Pires da Silva, J. (2021). Application of microRNA Database Mining in Biomarker Discovery and Identification of Therapeutic Targets for Complex Disease. Methods and Protocols, 4(1), 5. https://doi.org/10.3390/mps4010005