

Beyond Broca’s and Wernicke’s: Functional Mapping of Ancillary Language Centers Prior to Brain Tumor Surgery

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
3. Discussion
3.1. Language Laterality
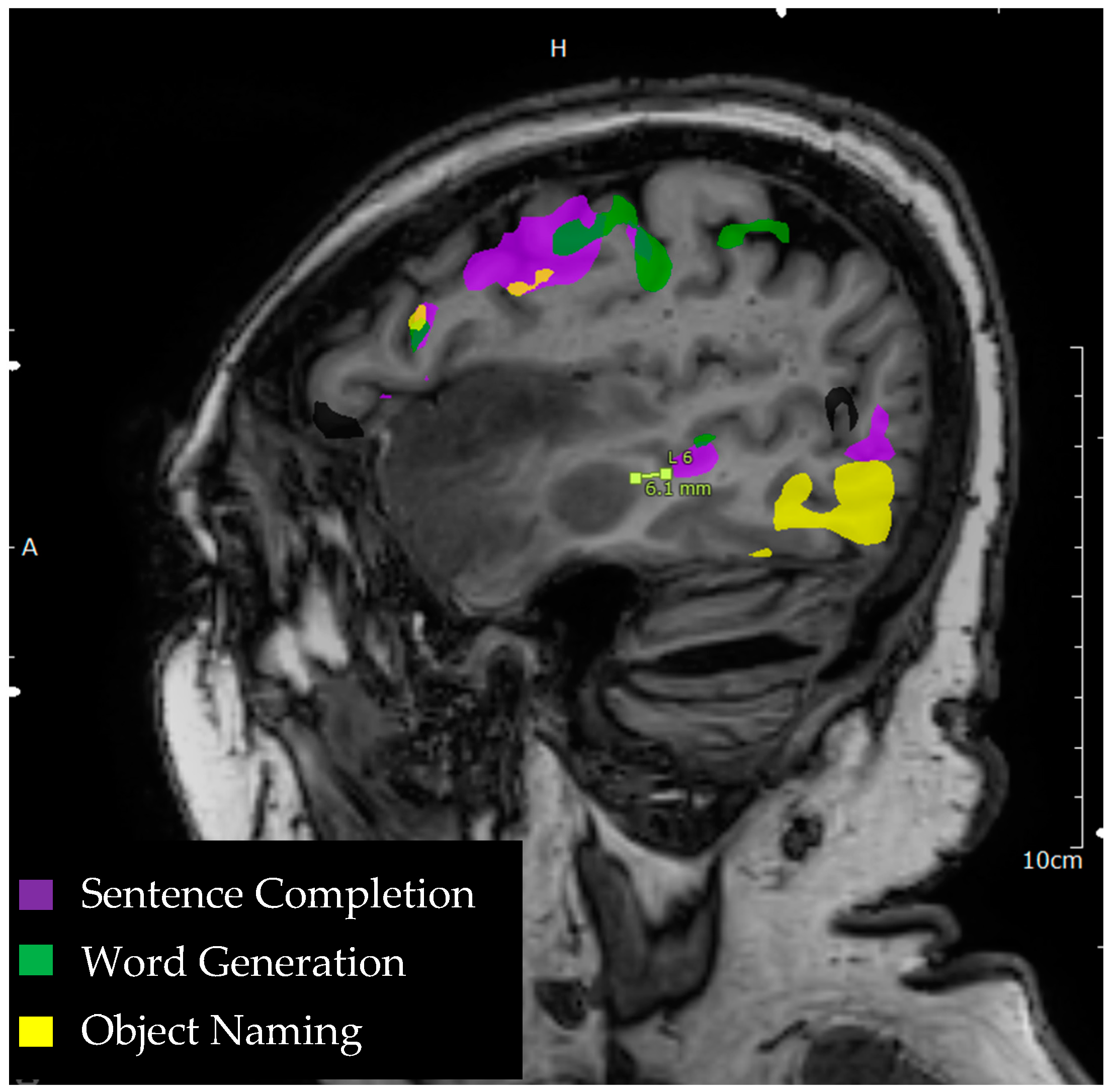
3.2. Lesion to Activation Distance
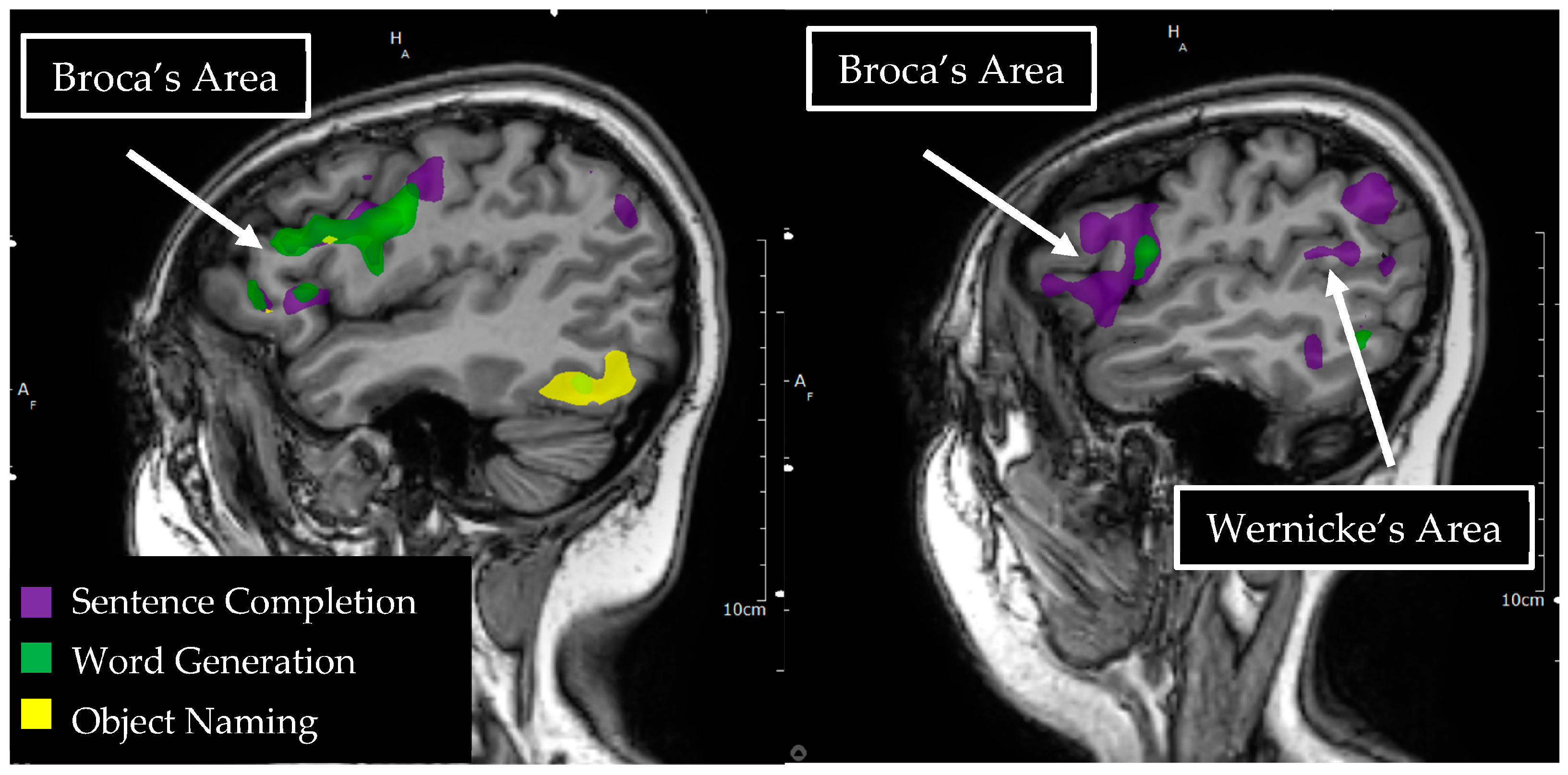
3.3. Broca’s Region
3.4. Wernicke’s Region
3.5. Ancillary Language Centers
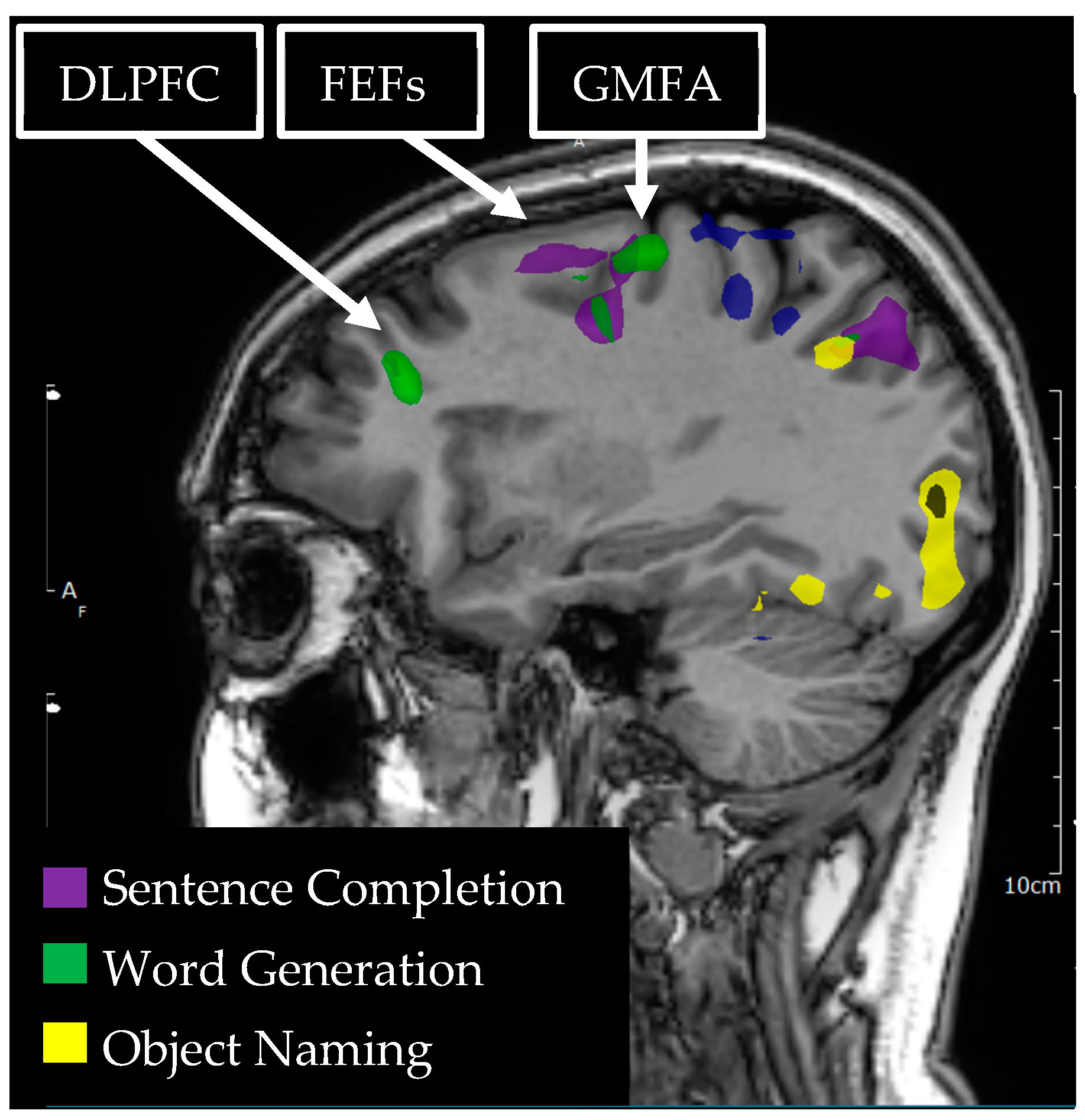
3.6. Dorsolateral Prefrontal Cortex
3.7. Graphemic Motor Frontal Area (Exner’s)
3.8. Frontal Eye Fields
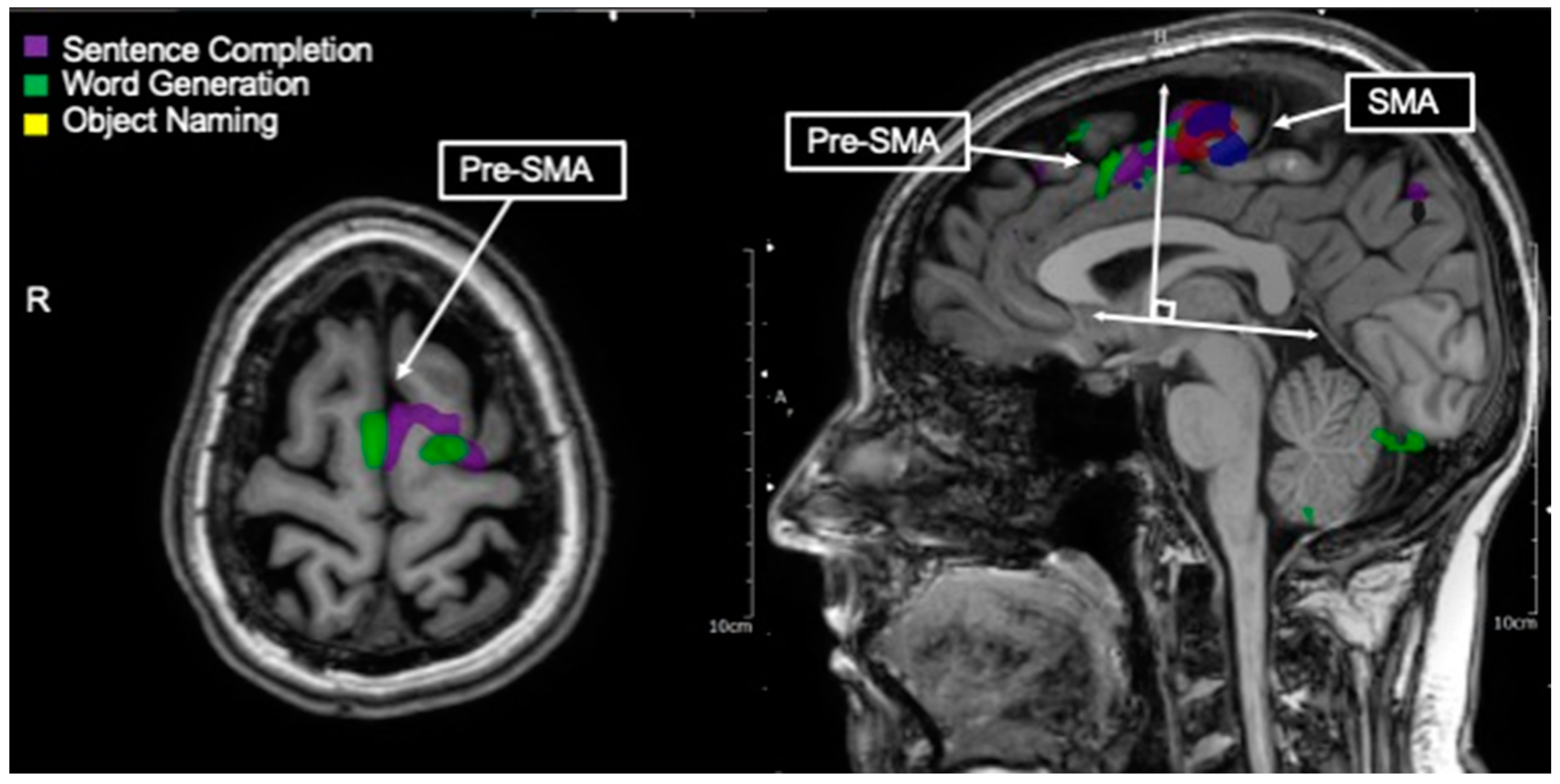
3.9. Pre-SMA/Language SMA
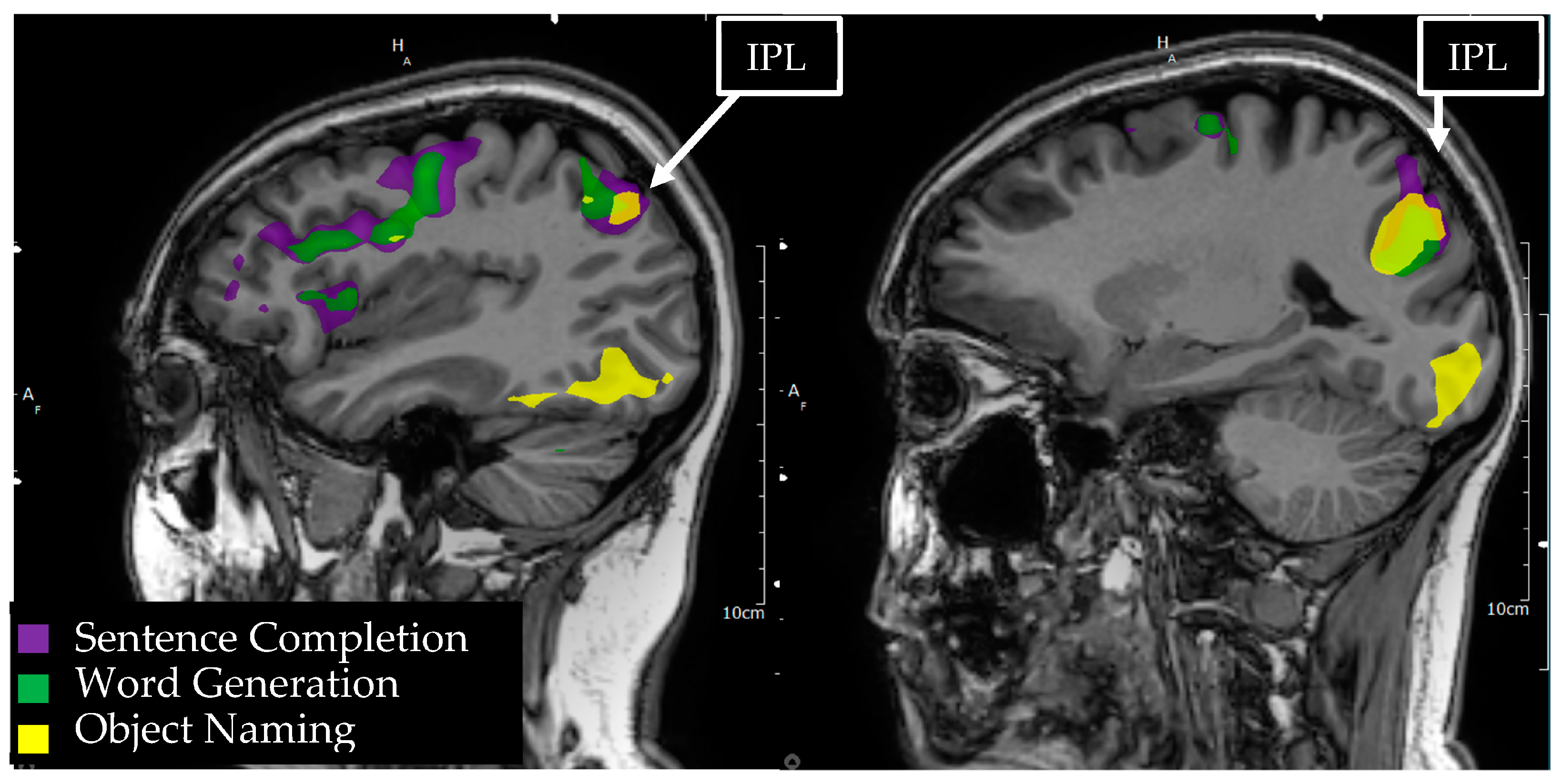
3.10. Inferior Parietal Lobule/Geschwind’s Area
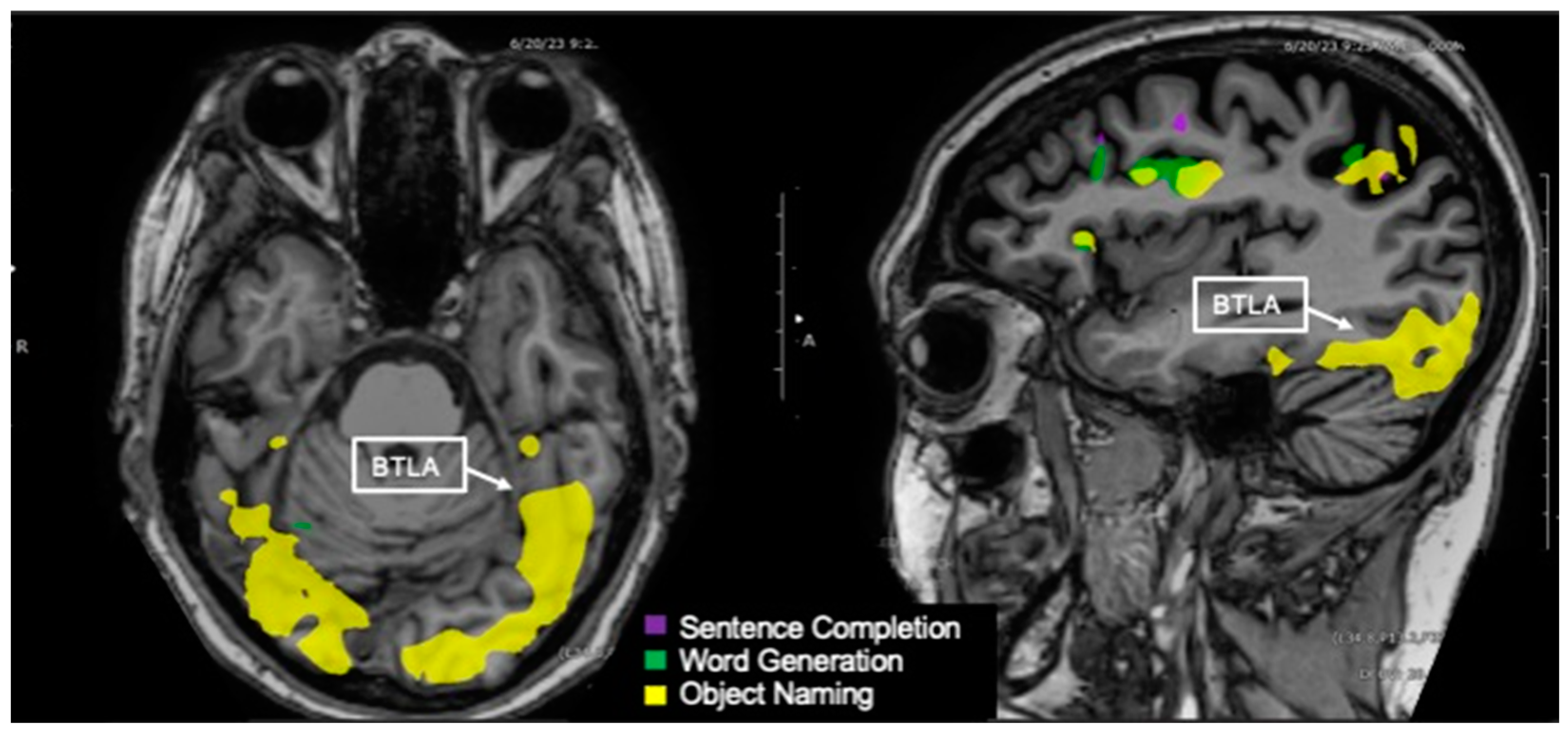
3.11. Basal Temporal Language Area
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Brown, T.; Shah, A.H.; Bregy, A.; Shah, N.H.; Thambuswamy, M.; Barbarite, E.; Fuhrman, T.; Komotar, R.J. Awake Craniotomy for Brain Tumor Resection: The Rule Rather Than the Exception? J. Neurosurg. Anesthesiol. 2013, 25, 240. [Google Scholar] [CrossRef] [PubMed]
- Bu, L.-H.; Zhang, J.; Lu, J.-F.; Wu, J.-S. Glioma surgery with awake language mapping versus generalized anesthesia: A systematic review. Neurosurg. Rev. 2021, 44, 1997–2011. [Google Scholar] [CrossRef] [PubMed]
- Gerritsen, J.K.W.; Viëtor, C.L.; Rizopoulos, D.; Schouten, J.W.; Klimek, M.; Dirven, C.M.F.; Vincent, A.J.-P.E. Awake craniotomy versus craniotomy under general anesthesia without surgery adjuncts for supratentorial glioblastoma in eloquent areas: A retrospective matched case-control study. Acta Neurochir. 2019, 161, 307–315. [Google Scholar] [CrossRef] [PubMed]
- De Witt Hamer, P.; Gil-Robles, S.; Zwinderman, A.; Duffau, H.; Berger, M. Impact of Intraoperative Stimulation Brain Mapping on Glioma Surgery Outcome: A Meta-Analysis. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2012, 30, 2559–2565. [Google Scholar] [CrossRef]
- De Benedictis, A.; Moritz-Gasser, S.; Duffau, H. Awake Mapping Optimizes the Extent of Resection for Low-Grade Gliomas in Eloquent Areas. Neurosurgery 2010, 66, 1074. [Google Scholar] [CrossRef]
- Reithmeier, T.; Krammer, M.; Gumprecht, H.; Gerstner, W.; Lumenta, C.B. Neuronavigation combined with electrophysiological monitoring for surgery of lesions in eloquent brain areas in 42 cases: A retrospective comparison of the neurological outcome and the quality of resection with a control group with similar lesions. Min-Minim. Invasive Neurosurg. 2003, 46, 65–71. [Google Scholar] [CrossRef]
- Duffau, H.; Lopes, M.; Arthuis, F.; Bitar, A.; Sichez, J.-P.; Van Effenterre, R.; Capelle, L. Contribution of intraoperative electrical stimulations in surgery of low grade gliomas: A comparative study between two series without (1985–1996) and with (1996–2003) functional mapping in the same institution. J. Neurol. Neurosurg. Psychiatry 2005, 76, 845–851. [Google Scholar] [CrossRef]
- Voets, N.L.; Bartsch, A.J.; Plaha, P. Functional MRI applications for intra-axial brain tumours: Uses and nuances in surgical practise. Br. J. Neurosurg. 2022, 1–16. [Google Scholar] [CrossRef]
- Jalilianhasanpour, R.; Beheshtian, E.; Ryan, D.; Luna, L.P.; Agarwal, S.; Pillai, J.J.; Sair, H.I.; Gujar, S.K. Role of Functional Magnetic Resonance Imaging in the Presurgical Mapping of Brain Tumors. Radiol. Clin. N. Am. 2021, 59, 377–393. [Google Scholar] [CrossRef]
- Arora, J.; Pugh, K.; Westerveld, M.; Spencer, S.; Spencer, D.D.; Todd Constable, R. Language lateralization in epilepsy patients: fMRI validated with the Wada procedure. Epilepsia 2009, 50, 2225–2241. [Google Scholar] [CrossRef]
- Rutten, G.J.M.; Ramsey, N.F.; van Rijen, P.C.; van Veelen, C.W.M. Reproducibility of fMRI-Determined Language Lateralization in Individual Subjects. Brain Lang. 2002, 80, 421–437. [Google Scholar] [CrossRef]
- Adcock, J.E.; Wise, R.G.; Oxbury, J.M.; Oxbury, S.M.; Matthews, P.M. Quantitative fMRI assessment of the differences in lateralization of language-related brain activation in patients with temporal lobe epilepsy. Neuroimage 2003, 18, 423–438. [Google Scholar] [CrossRef]
- Seghier, M.L.; Lazeyras, F.; Pegna, A.J.; Annoni, J.-M.; Zimine, I.; Mayer, E.; Michel, C.M.; Khateb, A. Variability of fMRI activation during a phonological and semantic language task in healthy subjects. Human Brain Mapp. 2004, 23, 140–155. [Google Scholar] [CrossRef]
- Nadkarni, T.N.; Andreoli, M.J.; Nair, V.A.; Yin, P.; Young, B.M.; Kundu, B.; Pankratz, J.; Radtke, A.; Holdsworth, R.; Kuo, J.S.; et al. Usage of fMRI for pre-surgical planning in brain tumor and vascular lesion patients: Task and statistical threshold effects on language lateralization. Neuroimage Clin. 2015, 7, 415–423. [Google Scholar] [CrossRef]
- Unadkat, P.; Fumagalli, L.; Rigolo, L.; Vangel, M.G.; Young, G.S.; Huang, R.; Mukundan, S., Jr.; Golby, A.; Tie, Y. Functional MRI Task Comparison for Language Mapping in Neurosurgical Patients. J. Neuroimaging 2019, 29, 348–356. [Google Scholar] [CrossRef]
- Bradshaw, A.R.; Bishop, D.V.M.; Woodhead, Z.V.J. Methodological considerations in assessment of language lateralisation with fMRI: A systematic review. PeerJ 2017, 5, e3557. [Google Scholar] [CrossRef]
- Knecht, S.; Jansen, A.; Frank, A.; van Randenborgh, J.; Sommer, J.; Kanowski, M.; Heinze, H.J. How atypical is atypical language dominance? Neuroimage 2003, 18, 917–927. [Google Scholar] [CrossRef]
- Abbott, D.F.; Waites, A.B.; Lillywhite, L.M.; Jackson, G.D. fMRI assessment of language lateralization: An objective approach. Neuroimage 2010, 50, 1446–1455. [Google Scholar] [CrossRef]
- Matsuo, K.; Chen, S.-H.A.; Tseng, W.-Y.I. AveLI: A robust lateralization index in functional magnetic resonance imaging using unbiased threshold-free computation. J. Neurosci. Methods 2012, 205, 119–129. [Google Scholar] [CrossRef]
- Branco, D.M.; Suarez, R.O.; Whalen, S.; O’Shea, J.P.; Nelson, A.P.; da Costa, J.C.; Golby, A.J. Functional MRI of memory in the hippocampus: Laterality indices may be more meaningful if calculated from whole voxel distributions. Neuroimage 2006, 32, 592–602. [Google Scholar] [CrossRef]
- Brumer, I.; Vita, E.D.; Ashmore, J.; Jarosz, J.; Borri, M. Implementation of clinically relevant and robust fMRI-based language lateralization: Choosing the laterality index calculation method. PLoS ONE 2020, 15, e0230129. [Google Scholar] [CrossRef] [PubMed]
- Ulmer, J.L.; Krouwer, H.G.; Mueller, W.M.; Ugurel, M.S.; Kocak, M.; Mark, L.P. Pseudo-Reorganization of Language Cortical Function at fMR Imaging: A Consequence of Tumor-Induced Neurovascular Uncoupling. Am. J. Neuroradiol. 2003, 24, 213–217. [Google Scholar] [PubMed]
- Pak, R.W.; Hadjiabadi, D.H.; Senarathna, J.; Agarwal, S.; Thakor, N.V.; Pillai, J.J.; Pathak, A.P. Implications of neurovascular uncoupling in functional magnetic resonance imaging (fMRI) of brain tumors. J. Cereb. Blood Flow Metab. 2017, 37, 3475–3487. [Google Scholar] [CrossRef] [PubMed]
- Pillai, J.J.; Zacá, D. Clinical utility of cerebrovascular reactivity mapping in patients with low grade gliomas. World J. Clin. Oncol. 2011, 2, 397–403. [Google Scholar] [CrossRef] [PubMed]
- Połczyńska, M.M.; Beck, L.; Kuhn, T.; Benjamin, C.F.; Ly, T.K.; Japardi, K.; Cavanagh, L.; Bookheimer, S.Y. Tumor location and reduction in functional MRI estimates of language laterality. J. Neurosurg. 2021, 135, 1674–1684. [Google Scholar] [CrossRef]
- Pillai, J.J.; Zacà, D. Comparison of BOLD cerebrovascular reactivity mapping and DSC MR perfusion imaging for prediction of neurovascular uncoupling potential in brain tumors. Technol. Cancer Res. Treat. 2012, 11, 361–374. [Google Scholar] [CrossRef]
- Zacà, D.; Jovicich, J.; Nadar, S.R.; Voyvodic, J.T.; Pillai, J.J. Cerebrovascular reactivity mapping in patients with low grade gliomas undergoing presurgical sensorimotor mapping with BOLD fMRI. J. Magn. Reson. Imaging 2014, 40, 383–390. [Google Scholar] [CrossRef]
- Gore, A.; Hu, R.; Patel, D.; Braileanu, M.; Hampton, D.; Joshi, H.; Kinger, N.; Louden, P.C.; O’Keefe, J.; Poliashenko, S.; et al. Combined task activation display as an effective method to teach introductory fMRI users. Clin. Imaging 2019, 55, 181–187. [Google Scholar] [CrossRef]
- Pillai, J.J.; Zaca, D. Relative utility for hemispheric lateralization of different clinical fMRI activation tasks within a comprehensive language paradigm battery in brain tumor patients as assessed by both threshold-dependent and threshold-independent analysis methods. Neuroimage 2011, 54 (Suppl. S1), S136–S145. [Google Scholar] [CrossRef]
- Oldfield, R.C. The assessment and analysis of handedness: The Edinburgh inventory. Neuropsychologia 1971, 9, 97–113. [Google Scholar] [CrossRef]
- Packheiser, J.; Schmitz, J.; Arning, L.; Beste, C.; Güntürkün, O.; Ocklenburg, S. A large-scale estimate on the relationship between language and motor lateralization. Sci. Rep. 2020, 10, 13027. [Google Scholar] [CrossRef]
- Badzakova-Trajkov, G.; Häberling, I.S.; Corballis, M.C. Cerebral asymmetries in monozygotic twins: An fMRI study. Neuropsychologia 2010, 48, 3086–3093. [Google Scholar] [CrossRef]
- Badzakova-Trajkov, G.; Häberling, I.S.; Roberts, R.P.; Corballis, M.C. Cerebral Asymmetries: Complementary and Independent Processes. PLoS ONE 2010, 5, e9682. [Google Scholar] [CrossRef]
- Szaflarski, J.P.; Holland, S.K.; Schmithorst, V.J.; Byars, A.W. fMRI study of language lateralization in children and adults. Human Brain Mapp. 2006, 27, 202–212. [Google Scholar] [CrossRef]
- Nenert, R.; Allendorfer, J.B.; Martin, A.M.; Banks, C.; Vannest, J.; Holland, S.K.; Szaflarski, J.P. Age-related language lateralization assessed by fMRI: The effects of sex and handedness. Brain Res. 2017, 1674, 20–35. [Google Scholar] [CrossRef]
- Stewart, C.C.; Swanson, S.J.; Sabsevitz, D.S.; Rozman, M.E.; Janecek, J.K.; Binder, J.R. Predictors of language lateralization in temporal lobe epilepsy. Neuropsychologia 2014, 60, 93–102. [Google Scholar] [CrossRef]
- Ocklenburg, S.; Güntürkün, O.; Hugdahl, K.; Hirnstein, M. Laterality and mental disorders in the postgenomic age—A closer look at schizophrenia and language lateralization. Neurosci. Biobehav. Rev. 2015, 59, 100–110. [Google Scholar] [CrossRef]
- Norton, E.S.; Beach, S.D.; Gabrieli, J.D. Neurobiology of dyslexia. Curr. Opin. Neurobiol. 2015, 30, 73–78. [Google Scholar] [CrossRef]
- Lindell, A.K.; Hudry, K. Atypicalities in Cortical Structure, Handedness, and Functional Lateralization for Language in Autism Spectrum Disorders. Neuropsychol. Rev. 2013, 23, 257–270. [Google Scholar] [CrossRef]
- Klimkeit, E.I.; Bradshaw, J.L. Anomalous Lateralisation in Neurodevelopmental Disorders. Cortex 2006, 42, 113–116. [Google Scholar] [CrossRef]
- Wood, J.M.; Kundu, B.; Utter, A.; Gallagher, T.A.; Voss, J.; Nair, V.A.; Kuo, J.S.; Field, A.S.; Moritz, C.H.; Meyerand, M.E.; et al. Impact of Brain Tumor Location on Morbidity and Mortality: A Retrospective Functional MR Imaging Study. Am. J. Neuroradiol. 2011, 32, 1420–1425. [Google Scholar] [CrossRef] [PubMed]
- Håberg, A.; Kvistad, K.A.; Unsgård, G.; Haraldseth, O.; Weiss, N.; Brem, H.; Takakura, K.; Sawaya, R.; Langmoen, I.A. Preoperative Blood Oxygen Level-dependent Functional Magnetic Resonance Imaging in Patients with Primary Brain Tumors: Clinical Application and Outcome. Neurosurgery 2004, 54, 902–915. [Google Scholar] [CrossRef] [PubMed]
- Haglund, M.M.; Berger, M.S.; Shamseldin, M.; Lettich, E.; Ojemann, G.A. Cortical Localization of Temporal Lobe Language Sites in Patients with Gliomas. Neurosurgery 1994, 34, 567. [Google Scholar] [PubMed]
- Kundu, B.; Penwarden, A.; Wood, J.M.; Gallagher, T.A.; Andreoli, M.J.; Voss, J.; Meier, T.; Nair, V.A.; Kuo, J.S.; Field, A.S.; et al. Association of functional magnetic resonance imaging indices with postoperative language outcomes in patients with primary brain tumors. Neurosurg. Focus 2013, 34, E6. [Google Scholar] [CrossRef] [PubMed]
- Riley, S.P.; Chu, D.Y.; Nair, V.A.; Baskaya, M.K.; Kuo, J.S.; Meyerand, M.E.; Prabhakaran, V. Characterizing the relationship between lesion-activation distance using fMRI and verbal measures in brain tumor patients. Interdiscip. Neurosurg. 2022, 27, 101391. [Google Scholar] [CrossRef]
- Mandonnet, E.; Duffau, H. Broca’s area: Why was neurosurgery neglected for so long when seeking to re-establish the scientific truth? Brain 2021, 144, e60. [Google Scholar] [CrossRef]
- Gajardo-Vidal, A.; Lorca-Puls, D.L.; team, P.; Warner, H.; Pshdary, B.; Crinion, J.T.; Leff, A.P.; Hope, T.M.H.; Geva, S.; Seghier, M.L.; et al. Damage to Broca’s area does not contribute to long-term speech production outcome after stroke. Brain 2021, 144, 817–832. [Google Scholar] [CrossRef]
- Andrews, J.P.; Cahn, N.; Speidel, B.A.; Chung, J.E.; Levy, D.F.; Wilson, S.M.; Berger, M.S.; Chang, E.F. Dissociation of Broca’s area from Broca’s aphasia in patients undergoing neurosurgical resections. J. Neurosurg. 2022, 1, 1–11. [Google Scholar] [CrossRef]
- Fedorenko, E.; Blank, I.A. Broca’s Area Is Not a Natural Kind. Trends Cogn. Sci. 2020, 24, 270–284. [Google Scholar] [CrossRef]
- Amunts, K.; Schleicher, A.; Bürgel, U.; Mohlberg, H.; Uylings, H.B.M.; Zilles, K. Broca’s region revisited: Cytoarchitecture and intersubject variability. J. Comp. Neurol. 1999, 412, 319–341. [Google Scholar] [CrossRef]
- Tomaiuolo, F.; MacDonald, J.D.; Caramanos, Z.; Posner, G.; Chiavaras, M.; Evans, A.C.; Petrides, M. Morphology, morphometry and probability mapping of the pars opercularis of the inferior frontal gyrus: An in vivo MRI analysis. Eur. J. Neurosci. 1999, 11, 3033–3046. [Google Scholar] [CrossRef]
- Fedorenko, E.; Hsieh, P.-J.; Nieto-Castañón, A.; Whitfield-Gabrieli, S.; Kanwisher, N. New Method for fMRI Investigations of Language: Defining ROIs Functionally in Individual Subjects. J. Neurophysiol. 2010, 104, 1177–1194. [Google Scholar] [CrossRef]
- Duncan, J.; Owen, A.M. Common regions of the human frontal lobe recruited by diverse cognitive demands. Trends Neurosci. 2000, 23, 475–483. [Google Scholar] [CrossRef]
- Hugdahl, K.; Raichle, M.E.; Mitra, A.; Specht, K. On the existence of a generalized non-specific task-dependent network. Front. Human Neurosci. 2015, 9, 430. Available online: https://www.frontiersin.org/articles/10.3389/fnhum.2015.00430 (accessed on 27 January 2023). [CrossRef]
- Tavor, I.; Jones, O.P.; Mars, R.B.; Smith, S.M.; Behrens, T.E.; Jbabdi, S. Task-free MRI predicts individual differences in brain activity during task performance. Science 2016, 352, 216–220. [Google Scholar] [CrossRef]
- Manan, H.A.; Franz, E.A.; Yahya, N. Utilization of functional MRI language paradigms for pre-operative mapping: A systematic review. Neuroradiology 2020, 62, 353–367. [Google Scholar] [CrossRef]
- Duffau, H. A personal consecutive series of surgically treated 51 cases of insular WHO Grade II glioma: Advances and limitations: Clinical article. J. Neurosurg. 2009, 110, 696–708. [Google Scholar] [CrossRef]
- Mandonnet, E. Transopercular Resection of IDH–Mutated Insular Glioma: A Critical Appraisal of an Initial Experience. World Neurosurg. 2019, 132, e563–e576. [Google Scholar] [CrossRef]
- Suarez-Meade, P.; Marenco-Hillembrand, L.; Sabsevitz, D.; Okromelidze, L.; Blake Perdikis, B.S.; Sherman, W.J.; Quinones-Hinojosa, A.; Middlebrooks, E.H.; Chaichana, K.L. Surgical resection of gliomas in the dominant inferior frontal gyrus: Consecutive case series and anatomy review of Broca’s area. Clin. Neurol. Neurosurg. 2022, 223, 107512. [Google Scholar] [CrossRef]
- Arora, S.; Vani, K. Translocation of Broca’s area in a case of low-grade glioma: Reinforcing the importance of preoperative functional magnetic resonance imaging. Egypt. J. Radiol. Nucl. Med. 2022, 53, 157. [Google Scholar] [CrossRef]
- Wernicke, C. Lehrbuch der Gehirnkrankheiten für Aerzte und Studirende. v.2-3, 1881–1883; Fischer: Kassel, Germany, 1881. [Google Scholar]
- Bogen, J.E.; Bogen, G.M. Wernicke’s region?where is it? Ann. N. Y. Acad. Sci. 1976, 280, 834–843. [Google Scholar] [CrossRef] [PubMed]
- Ardila, A.; Bernal, B.; Rosselli, M. The role of Wernicke’s area in language comprehension. Psychol. Neurosci. 2016, 9, 340–343. [Google Scholar] [CrossRef]
- Leff, A.P.; Schofield, T.M.; Crinion, J.T.; Seghier, M.L.; Grogan, A.; Green, D.W.; Price, C.J. The left superior temporal gyrus is a shared substrate for auditory short-term memory and speech comprehension: Evidence from 210 patients with stroke. Brain 2009, 132, 3401–3410. [Google Scholar] [CrossRef] [PubMed]
- Sarubbo, S.; Latini, F.; Sette, E.; Milani, P.; Granieri, E.; Fainardi, E.; Cavallo, M. Is the resection of gliomas in Wernicke’s area reliable? Wernicke’s area resection. Acta Neurochir. 2012, 154, 1653–1662. [Google Scholar] [CrossRef] [PubMed]
- Sarubbo, S.; Basso, G.; Chioffi, F.; Cesnik, E.; Paradiso, B.; Grandi, E.; Fainardi, E.; Tugnoli, V.; Farneti, M.; Granieri, E. Technical, Anatomical, and Functional Study after Removal of a Symptomatic Cavernous Angioma Located in Deep Wernicke’s Territories with Cortico-Subcortical Awake Mapping. Case Rep. Neurol. Med. 2013, 2013, e835029. [Google Scholar] [CrossRef]
- Salek, K.E.; Hassan, I.S.; Kotrotsou, A.; Abrol, S.; Faro, S.H.; Mohamed, F.B.; Zinn, P.O.; Wei, W.; Li, N.; Kumar, A.J.; et al. Silent Sentence Completion Shows Superiority Localizing Wernicke’s Area and Activation Patterns of Distinct Language Paradigms Correlate with Genomics: Prospective Study. Sci. Rep. 2017, 7, 12054. [Google Scholar] [CrossRef]
- Petrovich, N.M.; Holodny, A.I.; Brennan, C.W.; Gutin, P.H. Isolated Translocation of Wernicke’s Area to the Right Hemisphere in a 62-Year-Man with a Temporo-Parietal Glioma. Am. J. Neuroradiol. 2004, 25, 130–133. [Google Scholar]
- Sarubbo, S.; Bars, E.; Moritz-Gasser, S.; Duffau, H. Complete recovery after surgical resection of left Wernicke’s area in awake patient: A brain stimulation and functional MRI study. Neurosurg. Rev. 2011, 35, 287–292; discussion 292. [Google Scholar] [CrossRef]
- Duffau, H. Does post-lesional subcortical plasticity exist in the human brain? Neurosci. Res. 2009, 65, 131–135. [Google Scholar] [CrossRef]
- Herbet, G.; Duffau, H. Revisiting the Functional Anatomy of the Human Brain: Toward a Meta-Networking Theory of Cerebral Functions. Physiol. Rev. 2020, 100, 1181–1228. [Google Scholar] [CrossRef]
- Giampiccolo, D.; Duffau, H. Controversy over the temporal cortical terminations of the left arcuate fasciculus: A reappraisal. Brain 2022, 145, 1242–1256. [Google Scholar] [CrossRef]
- Pochon, J.-B.; Levy, R.; Poline, J.-B.; Crozier, S.; Lehéricy, S.; Pillon, B.; Deweer, B.; Le Bihan, D.; Dubois, B. The Role of Dorsolateral Prefrontal Cortex in the Preparation of Forthcoming Actions: An fMRI Study. Cereb. Cortex 2001, 11, 260–266. [Google Scholar] [CrossRef]
- Hertrich, I.; Dietrich, S.; Blum, C.; Ackermann, H. The Role of the Dorsolateral Prefrontal Cortex for Speech and Language Processing. Front. Human Neurosci. 2021, 15, 645209. Available online: https://www.frontiersin.org/articles/10.3389/fnhum.2021.645209 (accessed on 4 January 2023). [CrossRef]
- Klaus, J.; Schutter, D.J.L.G. The Role of Left Dorsolateral Prefrontal Cortex in Language Processing. Neuroscience 2018, 377, 197–205. [Google Scholar] [CrossRef]
- Motomura, K.; Chalise, L.; Ohka, F.; Aoki, K.; Tanahashi, K.; Hirano, M.; Nishikawa, T.; Wakabayashi, T.; Natsume, A. Supratotal Resection of Diffuse Frontal Lower Grade Gliomas with Awake Brain Mapping, Preserving Motor, Language, and Neurocognitive Functions. World Neurosurg. 2018, 119, 30–39. [Google Scholar] [CrossRef]
- Duffau, H.; Gatignol, P.; Mandonnet, E.; Peruzzi, P.; Tzourio-Mazoyer, N.; Capelle, L. New insights into the anatomo-functional connectivity of the semantic system: A study using cortico-subcortical electrostimulations. Brain 2005, 128, 797–810. [Google Scholar] [CrossRef]
- Duffau, H.; Leroy, M.; Gatignol, P. Cortico-subcortical organization of language networks in the right hemisphere: An electrostimulation study in left-handers. Neuropsychologia 2008, 46, 3197–3209. [Google Scholar] [CrossRef]
- Tate, M.C.; Herbet, G.; Moritz-Gasser, S.; Tate, J.E.; Duffau, H. Probabilistic map of critical functional regions of the human cerebral cortex: Broca’s area revisited. Brain 2014, 137, 2773–2782. [Google Scholar] [CrossRef]
- Herbet, G.; Moritz-Gasser, S.; Duffau, H. Electrical stimulation of the dorsolateral prefrontal cortex impairs semantic cognition. Neurology 2018, 90, e1077–e1084. [Google Scholar] [CrossRef]
- Campanella, F.; Crescentini, C.; Mussoni, A.; Skrap, M. Refractory semantic access dysphasia resulting from resection of a left frontal glioma. Neurocase 2013, 19, 27–35. [Google Scholar] [CrossRef]
- Jiao, L.; Meng, N.; Wang, Z.; Schwieter, J.W.; Liu, C. Partially shared neural mechanisms of language control and executive control in bilinguals: Meta-analytic comparisons of language and task switching studies. Neuropsychologia 2022, 172, 108273. [Google Scholar] [CrossRef] [PubMed]
- Newman, S.D.; Malaia, E.; Seo, R.; Cheng, H. The Effect of Individual Differences in Working Memory Capacity on Sentence Comprehension: An fMRI Study. Brain Topogr. 2013, 26, 458–467. [Google Scholar] [CrossRef] [PubMed]
- Buchweitz, A.; Mason, R.A.; Tomitch, L.M.B.; Just, M.A. Brain activation for reading and listening comprehension: An fMRI study of modality effects and individual differences in language comprehension. Psychol. Neurosci. 2009, 2, 111–123. [Google Scholar] [CrossRef] [PubMed]
- Ashtari, M.; Perrine, K.; Elbaz, R.; Syed, U.; Thaden, E.; McIlree, C.; Dolgoff-Kaspar, R.; Clarke, T.; Diamond, A.; Ettinger, A. Mapping the Functional Anatomy of Sentence Comprehension and Application to Presurgical Evaluation of Patients with Brain Tumor. Am. J. Neuroradiol. 2005, 26, 1461–1468. [Google Scholar] [PubMed]
- Black, D.F.; Vachha, B.; Mian, A.; Faro, S.H.; Maheshwari, M.; Sair, H.I.; Petrella, J.R.; Pillai, J.J.; Welker, K. American Society of Functional Neuroradiology–Recommended fMRI Paradigm Algorithms for Presurgical Language Assessment. Am. J. Neuroradiol. 2017, 38, E65–E73. [Google Scholar] [CrossRef]
- Sanjuán, A.; Bustamante, J.-C.; Forn, C.; Ventura-Campos, N.; Barrós-Loscertales, A.; Martínez, J.-C.; Villanueva, V.; Ávila, C. Comparison of two fMRI tasks for the evaluation of the expressive language function. Neuroradiology 2010, 52, 407–415. [Google Scholar] [CrossRef]
- Mark, I.T.; Black, D.F.; DeLone, D.R.; Passe, T.J.; Witte, R.J.; Little, J.T.; Ho, M.-L.; Fagan, A.J.; Parney, I.F.; Burns, T.C.; et al. Higher temporal resolution multiband fMRI provides improved presurgical language maps. Neuroradiology 2021, 63, 439–445. [Google Scholar] [CrossRef]
- Vinci-Booher, S.; Cheng, H.; James, K.H. An Analysis of the Brain Systems Involved with Producing Letters by Hand. J. Cogn. Neurosci. 2019, 31, 138. [Google Scholar] [CrossRef]
- Keller, C.; Meister, I.G. Agraphia caused by an infarction in Exner’s area. J. Clin. Neurosci. 2014, 21, 172–173. [Google Scholar] [CrossRef]
- Jiménez de la Peña, M.; Gil Robles, S.; Recio Rodríguez, M.; Ruiz Ocaña, C.; Martínez de Vega, V. Cortical and subcortical mapping of language areas: Correlation of functional MRI and tractography in a 3T scanner with intraoperative cortical and subcortical stimulation in patients with brain tumors located in eloquent areas. Radiología 2013, 55, 505–513. [Google Scholar] [CrossRef]
- Roux, F.-E.; Dufor, O.; Giussani, C.; Wamain, Y.; Draper, L.; Longcamp, M.; Démonet, J.-F. The graphemic/motor frontal area Exner’s area revisited. Ann. Neurol. 2009, 66, 537–545. [Google Scholar] [CrossRef]
- Scarone, P.; Gatignol, P.; Guillaume, S.; Denvil, D.; Capelle, L.; Duffau, H. Agraphia after awake surgery for brain tumor: New insights into the anatomo-functional network of writing. Surg. Neurol. 2009, 72, 223–241. [Google Scholar] [CrossRef]
- Ives-Deliperi, V.L.; Butler, J.T. Localizing the Language Network with fMRI and Functional Connectivity: Implications for Pre-Surgical Planning. Open J. Mod. Neurosurg. 2018, 8, 174. [Google Scholar] [CrossRef]
- Gkiatis, K.; Garganis, K.; Benjamin, C.F.; Karanasiou, I.; Kondylidis, N.; Harushukuri, J.; Matsopoulos, G.K. Standardization of presurgical language fMRI in Greek population: Mapping of six critical regions. Brain Behav. 2022, 12, e2609. [Google Scholar] [CrossRef]
- Benjamin, C.F.; Walshaw, P.D.; Hale, K.; Gaillard, W.D.; Baxter, L.C.; Berl, M.M.; Polczynska, M.; Noble, S.; Alkawadri, R.; Hirsch, L.J.; et al. Presurgical language fMRI: Mapping of six critical regions. Human Brain Mapp. 2017, 38, 4239–4255. [Google Scholar] [CrossRef]
- Pallud, J.; Zanello, M.; Kuchcinski, G.; Roux, A.; Muto, J.; Mellerio, C.; Dezamis, E.; Oppenheim, C. Individual Variability of the Human Cerebral Cortex Identified Using Intraoperative Mapping. World Neurosurg. 2018, 109, e313–e317. [Google Scholar] [CrossRef]
- Vernet, M.; Quentin, R.; Chanes, L.; Mitsumasu, A.; Valero-Cabré, A. Frontal eye field, where art thou? Anatomy, function, and non-invasive manipulation of frontal regions involved in eye movements and associated cognitive operations. Front. Integr. Neurosci. 2014, 8, 66. Available online: https://www.frontiersin.org/articles/10.3389/fnint.2014.00066 (accessed on 16 April 2023).
- Laatsch, L.; Krisky, C. Changes in fMRI activation following rehabilitation of reading and visual processing deficits in subjects with traumatic brain injury. Brain Inj. 2006, 20, 1367–1375. [Google Scholar] [CrossRef]
- Choi, W.; Desai, R.H.; Henderson, J.M. The neural substrates of natural reading: A comparison of normal and nonword text using eyetracking and fMRI. Front. Hum. Neurosci. 2014, 8, 1024. Available online: https://www.frontiersin.org/articles/10.3389/fnhum.2014.01024 (accessed on 16 April 2023). [CrossRef]
- Choi, W.; Henderson, J.M. Neural correlates of active vision: An fMRI comparison of natural reading and scene viewing. Neuropsychologia 2015, 75, 109–118. [Google Scholar] [CrossRef]
- Sun, L.; Li, C.; Wang, S.; Si, Q.; Lin, M.; Wang, N.; Sun, J.; Li, H.; Liang, Y.; Wei, J.; et al. Left frontal eye field encodes sound locations during passive listening. Cereb. Cortex 2023, 33, 3067–3079. [Google Scholar] [CrossRef] [PubMed]
- Milea, D.; Lobel, E.; Lehéricy, S.; Duffau, H.; Rivaud-Péchoux, S.; Berthoz, A.; Pierrot-Deseilligny, C. Intraoperative frontal eye field stimulation elicits ocular deviation and saccade suppression. NeuroReport 2002, 13, 1359. [Google Scholar] [CrossRef] [PubMed]
- Müller-Schunk, S.; Stephan, T.; Bense, S.; Ilmberger, J.; Gall, C.; Dieterich, M.; Reulen, H.J.; Yousry, T.A. Anatomic location of the frontal eye field: Comparison of fMRI and intraoperative mapping. Neuroimage 2000, 11, S669. [Google Scholar] [CrossRef]
- Usami, K.; Matsumoto, R.; Kunieda, T.; Shimotake, A.; Matsuhashi, M.; Miyamoto, S.; Fukuyama, H.; Takahashi, R.; Ikeda, A. Pre-SMA actively engages in conflict processing in human: A combined study of epicortical ERPs and direct cortical stimulation. Neuropsychologia 2013, 51, 1011–1017. [Google Scholar] [CrossRef] [PubMed]
- Nachev, P.; Kennard, C.; Husain, M. Functional role of the supplementary and pre-supplementary motor areas. Nat. Rev. Neurosci. 2008, 9, 856–869. [Google Scholar] [CrossRef]
- Picard, N.; Strick, P.L. Imaging the premotor areas. Curr. Opin. Neurobiol. 2001, 11, 663–672. [Google Scholar] [CrossRef]
- Nachev, P.; Wydell, H.; O’Neill, K.; Husain, M.; Kennard, C. The role of the pre-supplementary motor area in the control of action. Neuroimage 2007, 36, T155–T163. [Google Scholar] [CrossRef]
- Ziegler, W.; Kilian, B.; Deger, K. The role of the left mesial frontal cortex in fluent speech: Evidence from a case of left supplementary motor area hemorrhage. Neuropsychologia 1997, 35, 1197–1208. [Google Scholar] [CrossRef]
- Ackermann, H.; Hertrich, I.; Ziegler, W.; Bitzer, M.; Bien, S. Acquired dysfluencies following infarction of the left mesiofrontal cortex. Aphasiology 1996, 10, 409–417. [Google Scholar] [CrossRef]
- Van Borsel, J.; Van Lierde, K.; Van Cauwenberge, P.; Guldemont, I.; Van Orshoven, M. Severe acquired stuttering following injury of the left supplementary motor region: A case report. J. Fluen. Disord. 1998, 23, 49–58. [Google Scholar] [CrossRef]
- Chang, E.F.; Breshears, J.D.; Raygor, K.P.; Lau, D.; Molinaro, A.M.; Berger, M.S. Stereotactic probability and variability of speech arrest and anomia sites during stimulation mapping of the language dominant hemisphere. J. Neurosurg. 2017, 126, 114–121. [Google Scholar] [CrossRef]
- Quirarte, J.A.; Kumar, V.A.; Liu, H.-L.; Noll, K.R.; Wefel, J.S.; Lang, F.F. Language supplementary motor area syndrome correlated with dynamic changes in perioperative task-based functional MRI activations: Case report. J. Neurosurg. 2020, 134, 1738–1742. [Google Scholar] [CrossRef]
- Acioly, M.A.; Cunha, A.M.; Parise, M.; Rodrigues, E.; Tovar-Moll, F. Recruitment of Contralateral Supplementary Motor Area in Functional Recovery Following Medial Frontal Lobe Surgery: An fMRI Case Study. J. Neurol. Surg. Part A Cent. Eur. Neurosurg. 2015, 76, 508–512. [Google Scholar] [CrossRef]
- Endo, Y.; Saito, Y.; Otsuki, T.; Takahashi, A.; Nakata, Y.; Okada, K.; Hirozane, M.; Kaido, T.; Kaneko, Y.; Takada, E.; et al. Persistent verbal and behavioral deficits after resection of the left supplementary motor area in epilepsy surgery. Brain Dev. 2014, 36, 74–79. [Google Scholar] [CrossRef]
- Thiebaut de Schotten, M.; Dell’Acqua, F.; Valabregue, R.; Catani, M. Monkey to human comparative anatomy of the frontal lobe association tracts. Cortex 2012, 48, 82–96. [Google Scholar] [CrossRef]
- Dick, A.S.; Bernal, B.; Tremblay, P. The Language Connectome: New Pathways, New Concepts. Neuroscientist 2014, 20, 453–467. [Google Scholar] [CrossRef]
- Vassal, F.; Boutet, C.; Lemaire, J.-J.; Nuti, C. New insights into the functional significance of the frontal aslant tract: An anatomo–functional study using intraoperative electrical stimulations combined with diffusion tensor imaging-based fiber tracking. Br. J. Neurosurg. 2014, 28, 685–687. [Google Scholar] [CrossRef]
- Dragoy, O.; Zyryanov, A.; Bronov, O.; Gordeyeva, E.; Gronskaya, N.; Kryuchkova, O.; Klyuev, E.; Kopachev, D.; Medyanik, I.; Mishnyakova, L.; et al. Functional linguistic specificity of the left frontal aslant tract for spontaneous speech fluency: Evidence from intraoperative language mapping. Brain Lang. 2020, 208, 104836. [Google Scholar] [CrossRef]
- Agyemang, K.; Rose, A.; Sheikh, M.E.; Asha, M.; Molinari, E.; Fullerton, N.E.; Brennan, D.; Grivas, A. Two cases of SMA syndrome after neurosurgical injury to the frontal aslant tract. Acta Neurochir. 2023. [Google Scholar] [CrossRef]
- Binder, J.R. The Wernicke area: Modern evidence and a reinterpretation. Neurology 2015, 85, 2170–2175. [Google Scholar] [CrossRef]
- Abutalebi, J.; Della Rosa, P.A.; Ding, G.; Weekes, B.; Costa, A.; Green, D.W. Language proficiency modulates the engagement of cognitive control areas in multilinguals. Cortex 2013, 49, 905–911. [Google Scholar] [CrossRef] [PubMed]
- Hertrich, I.; Dietrich, S.; Ackermann, H. The role of the supplementary motor area for speech and language processing. Neurosci. Biobehav. Rev. 2016, 68, 602–610. [Google Scholar] [CrossRef] [PubMed]
- Lyo, J.K.; Arevalo-Perez, J.; Petrovich Brennan, N.; Peck, K.K.; Holodny, A.I. Pre-operative fMRI localization of the supplementary motor area and its relationship with postoperative speech deficits. Neuroradiol. J. 2015, 28, 281–288. [Google Scholar] [CrossRef] [PubMed]
- Krainik, A.; Lehéricy, S.; Duffau, H.; Capelle, L.; Chainay, H.; Cornu, P.; Cohen, L.; Boch, A.-L.; Mangin, J.-F.; Bihan, D.L.; et al. Postoperative speech disorder after medial frontal surgery: Role of the supplementary motor area. Neurology 2003, 60, 587–594. [Google Scholar] [CrossRef] [PubMed]
- Bailey, P.D.; Zacà, D.; Basha, M.M.; Agarwal, S.; Gujar, S.K.; Sair, H.I.; Eng, J.; Pillai, J.J. Presurgical fMRI and DTI for the Prediction of Perioperative Motor and Language Deficits in Primary or Metastatic Brain Lesions. J. Neuroimaging 2015, 25, 776–784. [Google Scholar] [CrossRef]
- Voss, J.; Meier, T.B.; Freidel, R.; Kundu, B.; Nair, V.A.; Holdsworth, R.; Kuo, J.S.; Prabhakaran, V. The role of secondary motor and language cortices in morbidity and mortality: A retrospective functional MRI study of surgical planning for patients with intracranial tumors. Neurosurg. Focus 2013, 34, E7. [Google Scholar] [CrossRef]
- Binkofski, F.C.; Klann, J.; Caspers, S. Chapter 4—On the Neuroanatomy and Functional Role of the Inferior Parietal Lobule and Intraparietal Sulcus. In Neurobiology of Language; Hickok, G., Small, S.L., Eds.; Academic Press: San Diego, CA, USA, 2016; pp. 35–47. ISBN 978-0-12-407794-2. [Google Scholar]
- Mirman, D.; Landrigan, J.-F.; Britt, A.E. Taxonomic and thematic semantic systems. Psychol. Bull. 2017, 143, 499–520. [Google Scholar] [CrossRef]
- Fridriksson, J.; Kjartansson, O.; Morgan, P.S.; Hjaltason, H.; Magnusdottir, S.; Bonilha, L.; Rorden, C. Impaired Speech Repetition and Left Parietal Lobe Damage. J. Neurosci. 2010, 30, 11057–11061. [Google Scholar] [CrossRef]
- Raizada, R.D.S.; Poldrack, R.A. Selective Amplification of Stimulus Differences during Categorical Processing of Speech. Neuron 2007, 56, 726–740. [Google Scholar] [CrossRef]
- Sakurai, Y.; Asami, M.; Mannen, T. Alexia and agraphia with lesions of the angular and supramarginal gyri: Evidence for the disruption of sequential processing. J. Neurol. Sci. 2010, 288, 25–33. [Google Scholar] [CrossRef]
- Southwell, D.G.; Riva, M.; Jordan, K.; Caverzasi, E.; Li, J.; Perry, D.W.; Henry, R.G.; Berger, M.S. Language outcomes after resection of dominant inferior parietal lobule gliomas. J. Neurosurg. 2017, 127, 781–789. [Google Scholar] [CrossRef]
- Maldonado, I.L.; Moritz-Gasser, S.; de Champfleur, N.M.; Bertram, L.; Moulinié, G.; Duffau, H. Surgery for gliomas involving the left inferior parietal lobule: New insights into the functional anatomy provided by stimulation mapping in awake patients: Clinical article. J. Neurosurg. 2011, 115, 770–779. [Google Scholar] [CrossRef]
- Cousin, E.; Peyrin, C.; Pichat, C.; Lamalle, L.; Le Bas, J.-F.; Baciu, M. Functional MRI approach for assessing hemispheric predominance of regions activated by a phonological and a semantic task. Eur. J. Radiol. 2007, 63, 274–285. [Google Scholar] [CrossRef]
- Roux, F.-E.; Boulanouar, K.; Lotterie, J.-A.; Mejdoubi, M.; LeSage, J.P.; Berry, I. Language Functional Magnetic Resonance Imaging in Preoperative Assessment of Language Areas: Correlation with Direct Cortical Stimulation. Neurosurgery 2003, 52, 1335–1347. [Google Scholar] [CrossRef]
- Lüders, H.; Hahn, J.; Lesser, R.P.; Dinner, D.S.; Iii, H.H.M.; Wyllie, E.; Friedman, L.; Friedman, D.; Skipper, G. Basal Temporal Subdural Electrodes in the Evaluation of Patients with Intractable Epilepsy. Epilepsia 1989, 30, 131–142. [Google Scholar] [CrossRef]
- Lüders, H.; Lesser, R.P.; Dinner, D.S.; Morris, H.H.; Wyllie, E.; Godoy, J. Localization of Cortical Function: New Information from Extraoperative Monitoring of Patients with Epilepsy. Epilepsia 1988, 29, S56–S65. [Google Scholar] [CrossRef]
- Lüders, H.; Lesser, R.P.; Hahn, J.; Dinner, D.S.; Morris, H.H.; Wyllie, E.; Godoy, J. Basal temporal language area. Brain 1991, 114 Pt 2, 743–754. [Google Scholar] [CrossRef]
- Pouratian, N.; Bookheimer, S.Y.; Rubino, G.; Martin, N.A.; Toga, A.W. Category-specific naming deficit identified by intraoperative stimulation mapping and postoperative neuropsychological testing: Case report. J. Neurosurg. 2003, 99, 170–176. [Google Scholar] [CrossRef]
- Usui, K.; Ikeda, A.; Takayama, M.; Matsuhashi, M.; Yamamoto, J.; Satoh, T.; Begum, T.; Mikuni, N.; Takahashi, J.B.; Miyamoto, S.; et al. Conversion of semantic information into phonological representation: A function in left posterior basal temporal area. Brain 2003, 126, 632–641. [Google Scholar] [CrossRef]
- Rigolo, L.; Essayed, W.I.; Tie, Y.; Norton, I.; Mukundan, S., Jr.; Golby, A. Intraoperative Use of Functional MRI for Surgical Decision Making after Limited or Infeasible Electrocortical Stimulation Mapping. J. Neuroimaging 2020, 30, 184–191. [Google Scholar] [CrossRef]
- Prasse, G.; Meyer, H.-J.; Scherlach, C.; Maybaum, J.; Hoffmann, A.; Kasper, J.; Karl Fehrenbach, M.; Wilhelmy, F.; Meixensberger, J.; Hoffmann, K.-T.; et al. Preoperative language tract integrity is a limiting factor in recovery from aphasia after glioma surgery. Neuroimage Clin. 2023, 37, 103310. [Google Scholar] [CrossRef] [PubMed]
- Ghinda, D.C.; Wu, J.-S.; Duncan, N.W.; Northoff, G. How much is enough—Can resting state fMRI provide a demarcation for neurosurgical resection in glioma? Neurosci. Biobehav. Rev. 2018, 84, 245–261. [Google Scholar] [CrossRef] [PubMed]
- Yuan, B.; Zhang, N.; Yan, J.; Cheng, J.; Lu, J.; Wu, J. Resting-state functional connectivity predicts individual language impairment of patients with left hemispheric gliomas involving language network. Neuroimage Clin. 2019, 24, 102023. [Google Scholar] [CrossRef] [PubMed]
- Sparacia, G.; Parla, G.; Lo Re, V.; Cannella, R.; Mamone, G.; Carollo, V.; Midiri, M.; Grasso, G. Resting-State Functional Connectome in Patients with Brain Tumors Before and After Surgical Resection. World Neurosurg. 2020, 141, e182–e194. [Google Scholar] [CrossRef]
- Fox, M.E.; King, T.Z. Functional Connectivity in Adult Brain Tumor Patients: A Systematic Review. Brain Connect. 2018, 8, 381–397. [Google Scholar] [CrossRef]
- Vakamudi, K.; Posse, S.; Jung, R.; Cushnyr, B.; Chohan, M.O. Real-time presurgical resting-state fMRI in patients with brain tumors: Quality control and comparison with task-fMRI and intraoperative mapping. Human Brain Mapp. 2020, 41, 797–814. [Google Scholar] [CrossRef]
- Lemée, J.-M.; Berro, D.H.; Bernard, F.; Chinier, E.; Leiber, L.-M.; Menei, P.; Ter Minassian, A. Resting-state functional magnetic resonance imaging versus task-based activity for language mapping and correlation with perioperative cortical mapping. Brain Behav. 2019, 9, e01362. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lawrence, A.; Carvajal, M.; Ormsby, J. Beyond Broca’s and Wernicke’s: Functional Mapping of Ancillary Language Centers Prior to Brain Tumor Surgery. Tomography 2023, 9, 1254-1275. https://doi.org/10.3390/tomography9040100
Lawrence A, Carvajal M, Ormsby J. Beyond Broca’s and Wernicke’s: Functional Mapping of Ancillary Language Centers Prior to Brain Tumor Surgery. Tomography. 2023; 9(4):1254-1275. https://doi.org/10.3390/tomography9040100
Chicago/Turabian StyleLawrence, Ashley, Michael Carvajal, and Jacob Ormsby. 2023. "Beyond Broca’s and Wernicke’s: Functional Mapping of Ancillary Language Centers Prior to Brain Tumor Surgery" Tomography 9, no. 4: 1254-1275. https://doi.org/10.3390/tomography9040100
APA StyleLawrence, A., Carvajal, M., & Ormsby, J. (2023). Beyond Broca’s and Wernicke’s: Functional Mapping of Ancillary Language Centers Prior to Brain Tumor Surgery. Tomography, 9(4), 1254-1275. https://doi.org/10.3390/tomography9040100