
Locomotory Behavior of Water Striders with Amputated Legs




Abstract
:1. Introduction
2. Material and Methods
2.1. Animals
2.2. Protocol of Measurement of Load Changes
2.3. Experimental Setup
2.4. Analysis Procedure
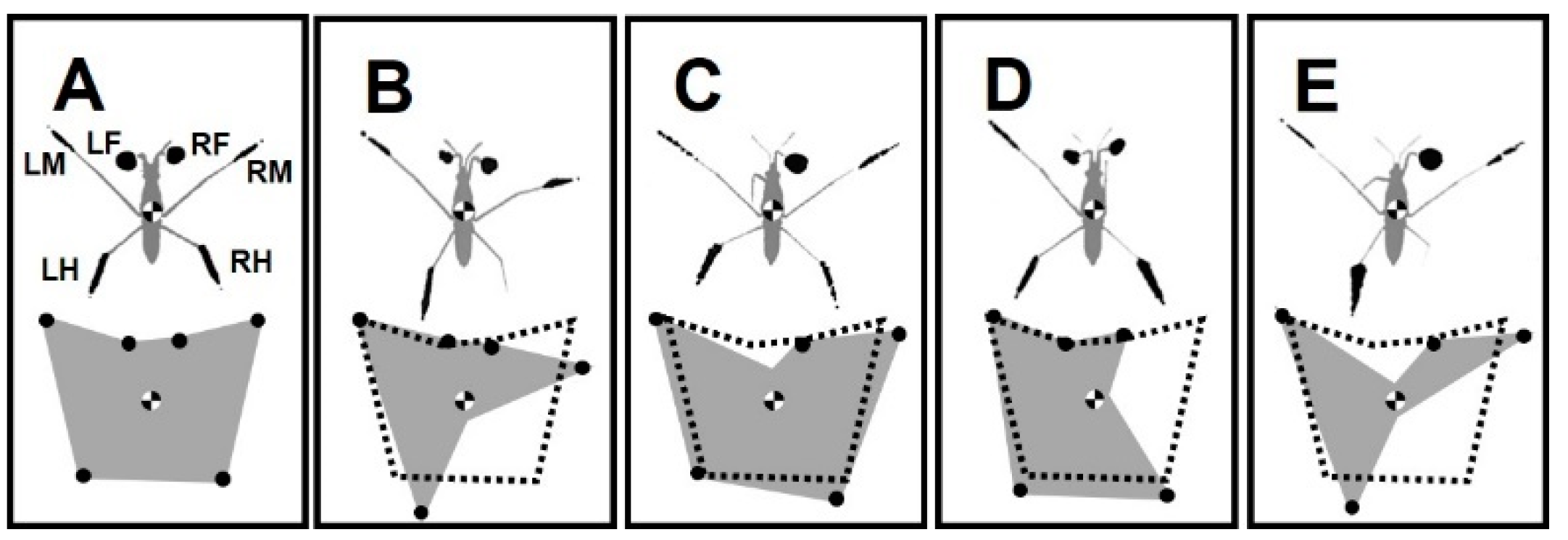
2.5. Labeling of Legs and Their Amputations
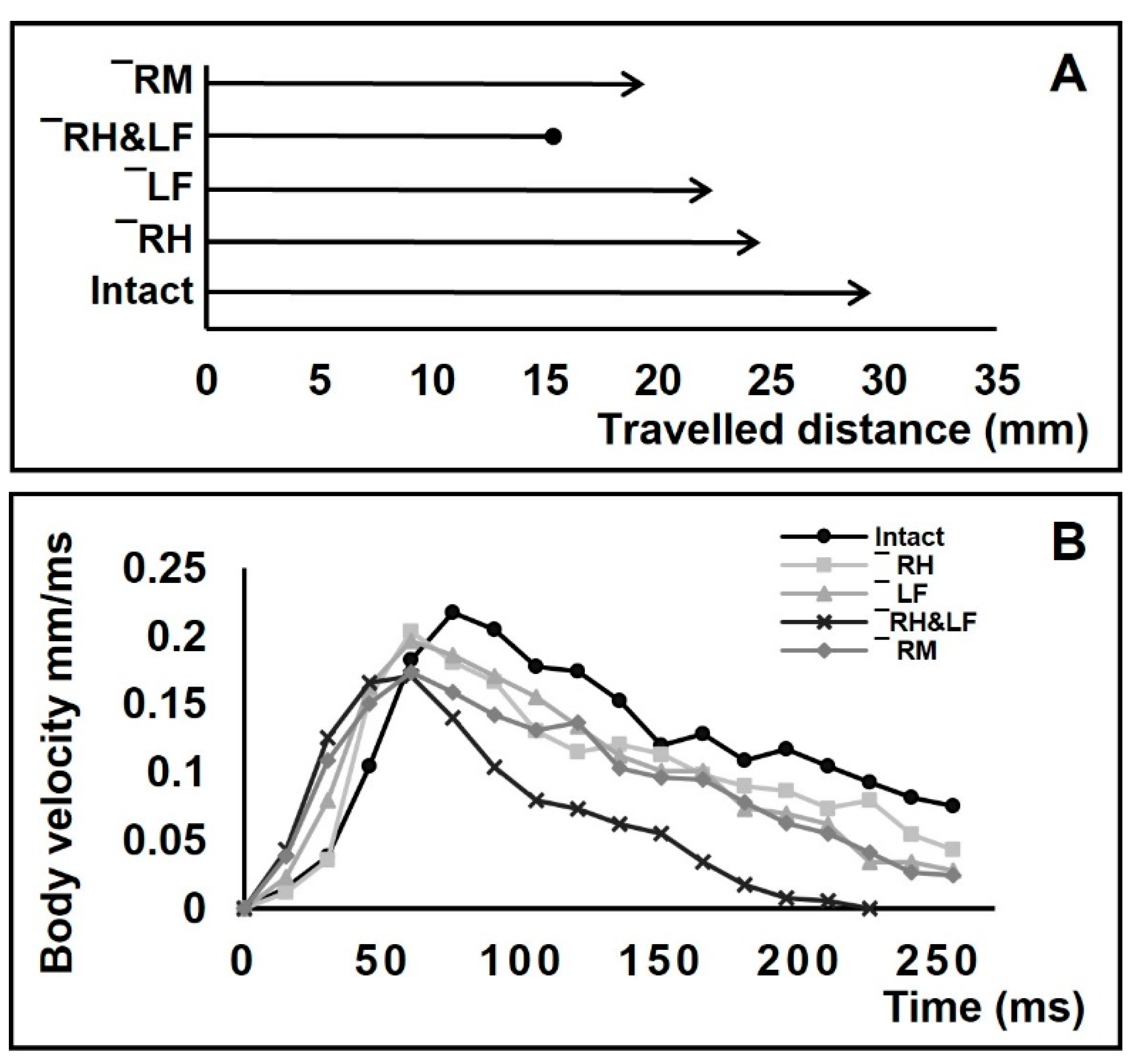
3. Results
3.1. Severity Grades of Amputations
3.2. Changes in Standing Posture
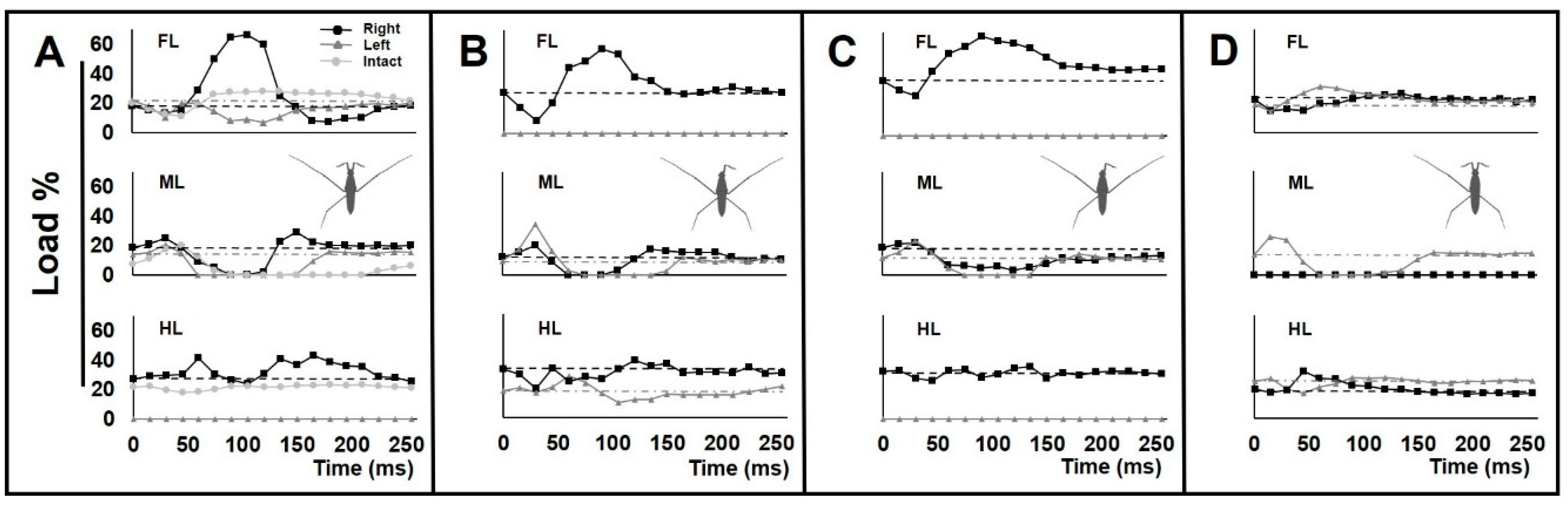
3.3. Load Change on the Legs during Locomotion
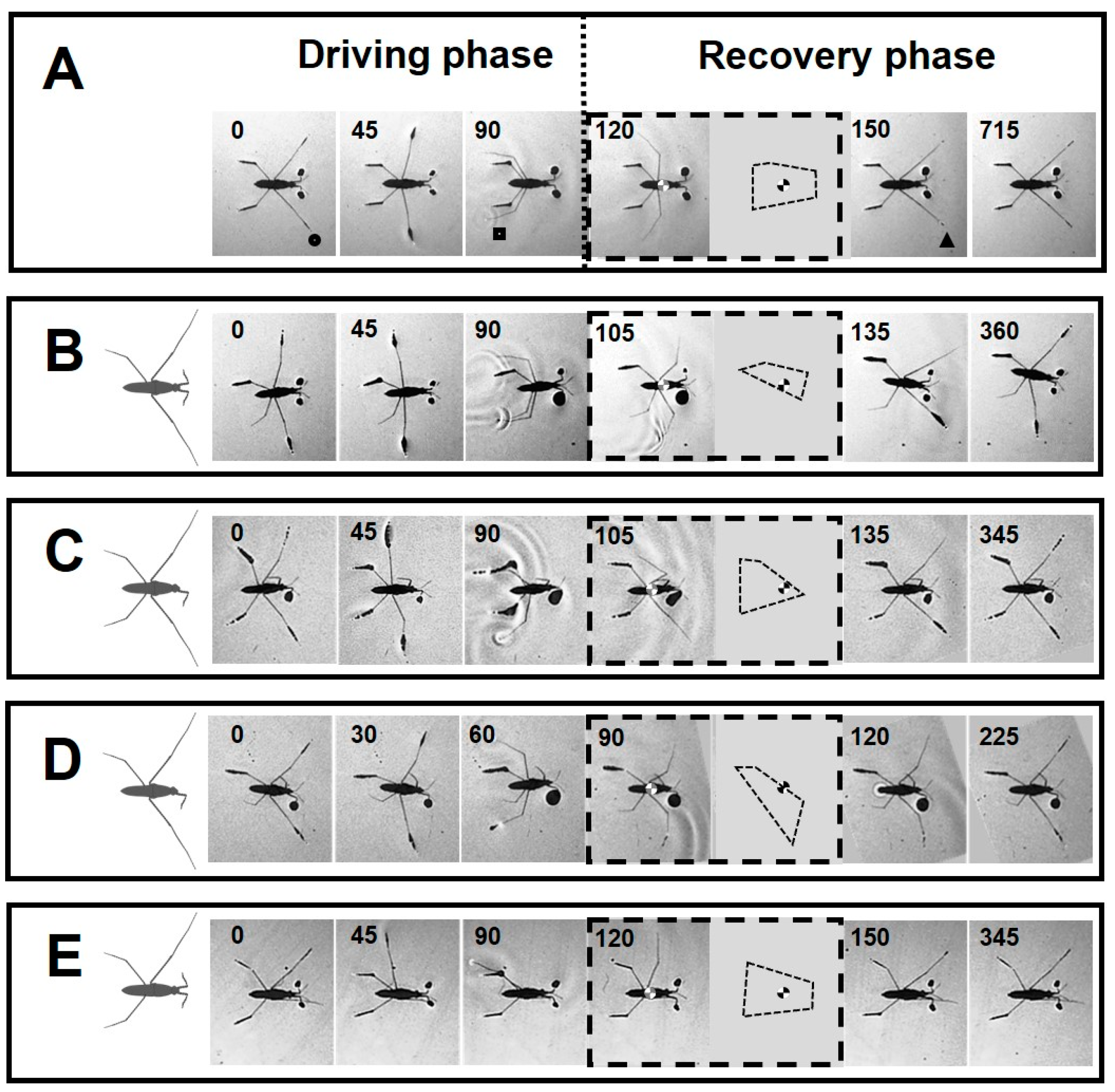
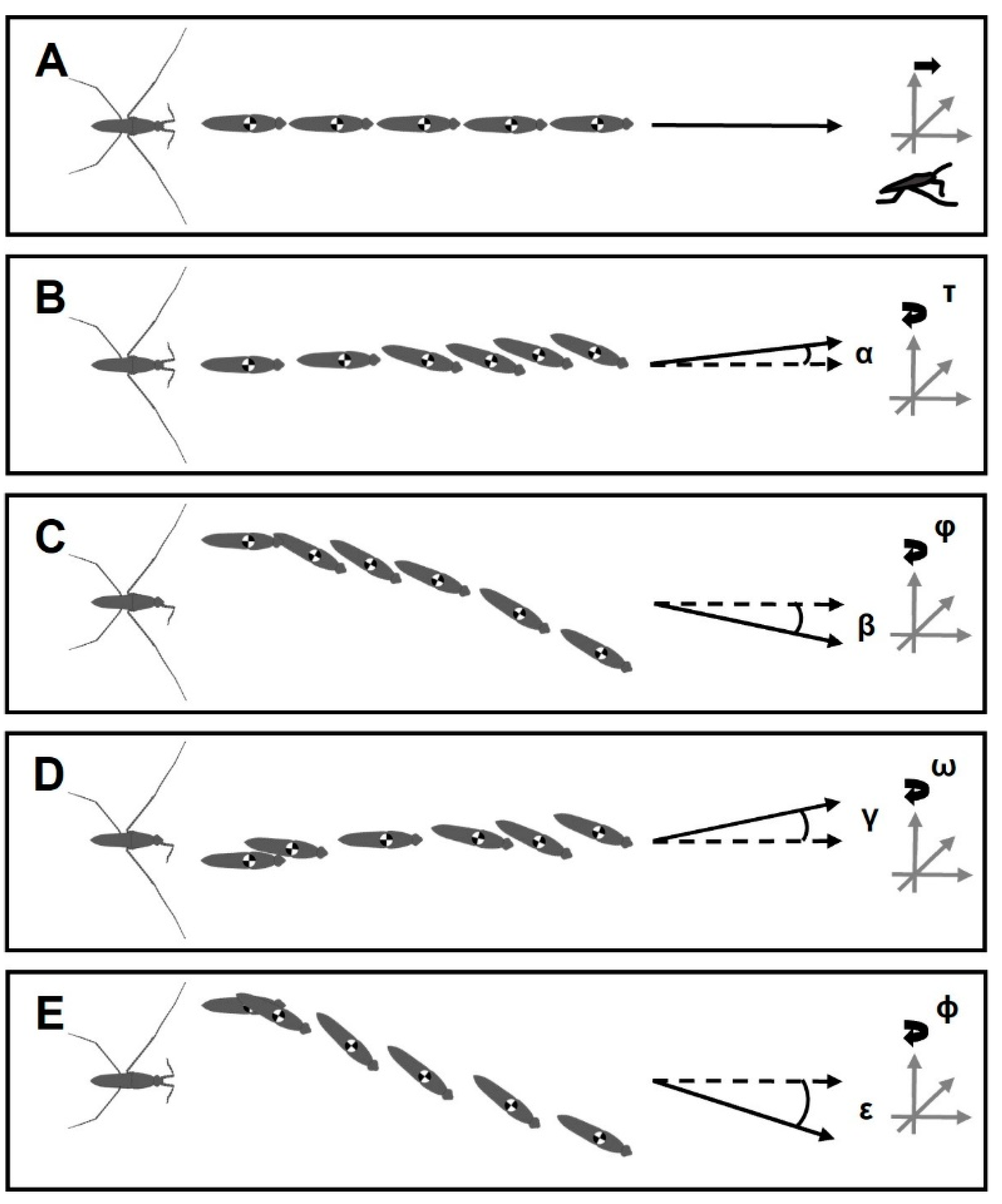
3.4. Positioning of Bodies during Striding Cycle
3.5. Jumping Ability
4. Discussion
4.1. Postural Change after Amputation
4.2. The Presence of All Legs Is Essential for Straightforward Striding
4.3. Asymmetrical Load Changes on the Legs during Locomotion
4.4. Changes in Locomotory Behavior after Amputations
4.5. Characterizations of Striding after Amputation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hughes, G.M. The Co-Ordination of Insect Movements: I The Walking Movements of Insects. J. Exp. Biol. 1952, 29, 267–285. [Google Scholar] [CrossRef]
- Cruse, H. The Function of the Legs in the Free Walking Stick Insect, Carausius morosus. J. Comp. Physiol. A 1976, 112, 235–262. [Google Scholar] [CrossRef]
- Delcomyn, F. Insect Locomotion on Land. In Locomotion and Energetics in Arthropods; Springer US: New York, NY, USA, 1981; pp. 69–91. [Google Scholar] [CrossRef]
- Full, R.J.; Tu, M.S. Mechanics of a Rapid Running Insect: Two-, Four-, and Six-Legged Locomotion. J. Exp. Biol. 1991, 156, 215–231. [Google Scholar] [CrossRef] [PubMed]
- Dürr, V.; Theunissen, L.M.; Dallmann, C.J.; Hoinville, T.; Schmitz, J. Motor Flexibility in Insects: Adaptive Coordination of Limbs in Locomotion and Near-range Exploration. Behav. Ecol. Sociobiol. 2018, 72, 15. [Google Scholar] [CrossRef]
- Schmitt, J.; Holmes, P. Mechanical Models for Insect Locomotion: Dynamics and Stability in the Horizontal Plane I. Theory. Biol. Cybern. 2000, 83, 501–515. [Google Scholar] [CrossRef] [PubMed]
- Weihmann, T. The Smooth Transition From Many-Legged to Bipedal Locomotion-Gradual Leg Force Reduction and its Impact on Total Ground Reaction Forces, Body Dynamics and Gait Transitions. Front. Bioeng. Biotechnol. 2022, 9, 769684. [Google Scholar] [CrossRef] [PubMed]
- Andersen, N.M. The Semiaquatic Bugs; Scandinavian Science Press Ltd.: Klampenborg, Denmark, 1982; Volume 3. [Google Scholar]
- Andersen, N.M. The evolution of marine insects: Phylogenetic, ecological and geographical aspects of species diversity in marine water striders. Ecography 1999, 22, 98–111. [Google Scholar] [CrossRef]
- Crumière, A.J.J.; Santos, M.E.; Sémon, M.; Armisén, D.; Moreira, F.F.F.; Khila, A. Diversity in Morphology and Locomotory Behavior Is Associated with Niche Expansion in the Semi-aquatic Bugs. Curr. Biol. 2016, 26, 3336–3342. [Google Scholar] [CrossRef]
- Ortega-Jimenez, V.M.; von Rabenau, L.; Dudley, R. Escape Jumping by Three Age-Classes of Water Striders from Smooth, Wavy and Bubbling Water Surfaces. J. Exp. Biol. 2017, 220, 2809–2815. [Google Scholar] [CrossRef]
- Bush, J.W.M.; Hu, D.; Prakash, M. The Integument of Water-walking Arthropods: Form and Function. Adv. Insect Physiol. 2007, 34, 117–192. [Google Scholar] [CrossRef]
- Denny, M.W. Paradox Lost: Answers and Questions about Walking on Water. J. Exp. Biol. 2004, 207, 1601–1606. [Google Scholar] [CrossRef] [PubMed]
- Hu, D.L.; Chan, B.; Bush, J.W.M. The Hydrodynamics of Water Strider Locomotion. Nature 2003, 424, 663–666. [Google Scholar] [CrossRef]
- White, H.E. Modern College Physics; van Nostrand: New York, NY, USA, 1948; ISBN 978-0-442-29401-4. [Google Scholar]
- Andersen, N.M. A comparative study of locomotion on the water surface in semiaquatic bugs (Insects, Hemiptera, Gerromorpha). Videnskabelige Meddelelser fra Dansk Naturhistorisk Forening 1976, 139, 337–396. [Google Scholar]
- Caponigro, M.A.; Eriksen, C.H. Surface Film Locomotion by the Water Strider, Gerris remigis Say. Am. Midl. Nat. 1976, 95, 268–278. [Google Scholar] [CrossRef]
- Hu, D.L.; Bush, J.W.M. The Hydrodynamics of Water-Walking Arthropods. J. Fluid Mech. 2010, 644, 5–33. [Google Scholar] [CrossRef]
- Kim, H.; Amauger, J.; Jeong, H.; Lee, D.; Yang, E.; Jablonski, P.G. Mechanics of Jumping on Water. Phys. Rev. Fluids 2017, 2, 100505. [Google Scholar] [CrossRef]
- Kovalev, A.; Filippov, A.E.; Gorb, S.N. Numerical Model of the Spatio-Temporal Dynamics in a Water Strider Group. Sci. Rep. 2021, 11, 18047. [Google Scholar] [CrossRef]
- Waldbauer, G.A. Walk around the Pond: Insects in and over the Water; Harvard University Press: Cambridge, MA, USA, 2006; pp. 109–111. [Google Scholar]
- Armisén, D.; Nagui Refki, P.; Crumière, A.J.J.; Viala, S.; Toubiana, W.; Khila, A. Predator strike shapes antipredator phenotype through new genetic interactions in water striders. Nat. Commun. 2015, 6, 8153. [Google Scholar] [CrossRef]
- Haskins, K.; Sih, A.; Krupa, J. Predation Risk and Social Interference as Factors Influencing Habitat Selection in Two Species of Stream-Dwelling Waterstriders. Behav. Ecol. 1997, 8, 351–363. [Google Scholar] [CrossRef]
- Krupa, J.J.; Sih, A. Comparison of Antipredator Responses of Two Related Water Striders to a Common Predator. Ethology 1999, 105, 1019–1033. [Google Scholar] [CrossRef]
- Gruhn, M.; Zehl, L.; Buschges, A. Straight Walking and Turning on a Slippery Surface. J. Exp. Biol. 2008, 212, 194–209. [Google Scholar] [CrossRef] [PubMed]
- Merienne, H.; Latil, G.; Moretto, P.; Fourcassié, V. Walking Kinematics in the Polymorphic Seed Harvester Ant Messor barbarus: Influence of Body Size and Load Carriage. J. Exp. Biol. 2020, 223, jeb205690. [Google Scholar] [CrossRef]
- Merienne, H.; Latil, G.; Moretto, P.; Fourcassié, V. Dynamics of Locomotion in the Seed Harvesting Ant Messor barbarus: Effect of Individual Body Mass and Transported Load Mass. PeerJ 2021, 9, e10664. [Google Scholar] [CrossRef] [PubMed]
- Ting, L.H.; Blickhan, R.; Full, R.J. Dynamic and Static Stability in Hexapedal Runners. J. Exp. Biol. 1994, 197, 251–269. [Google Scholar] [CrossRef]
- Baek, M.; Lawin, K.M.; Codden, C.J.; Lim, H.; Yang, E.; Kim, H.; Lee, S.; Jablonski, P.G. Water strider females use individual experience to adjust jumping behaviour to their weight within physical constraints of water surface tension. Sci. Rep. 2020, 10, 18657. [Google Scholar] [CrossRef]
- Koh, J.S.; Yang, E.; Jung, G.P.; Jung, S.P.; Son, J.H.; Lee, S.I.; Jablonski, P.G.; Wood, R.J.; Kim, H.Y.; Cho, K.J. Jumping on Water: Surface Tension-Dominated Jumping of Water Striders and Robotic Insects. Science 2015, 349, 517–521. [Google Scholar] [CrossRef]
- Meshkani, J.; Rajabi, H.; Kovalev, A.; Gorb, S.N. Patterns of Load Distribution among the Legs in Small Water Striders during Standing and Striding. J. Zool. 2023, 320, 84–95. [Google Scholar] [CrossRef]
- Yang, E.; Son, J.H.; Lee, S.I.; Jablonski, P.G.; Kim, H.Y. Water Striders Adjust Leg Movement Speed to Optimize Takeoff Velocity for Their Morphology. Nat. Commun. 2016, 7, 13698. [Google Scholar] [CrossRef]
- Perez Goodwyn, P.; Maezono, Y.; Takamatsu, H.; Fujisaki, K. Semiaquatic Heteroptera Locomotion: Coral Treaders (Hermatobates weddi, Hermatobatidae), Sea Skaters (Halovelia septentrionalis, Veliidae), and Water Striders (Metrocoris histrio, Gerridae). Usual and Unusual Gaits. Hydrobiologia 2009, 630, 219–229. [Google Scholar] [CrossRef]
- Fleming, P.; Bateman, P. Just Drop It and Run: The Effect of Limb Autotomy on Running Distance and Locomotion Energetics of Field Crickets (Gryllus bimaculatus). J. Exp. Biol. 2007, 210, 1446–1454. [Google Scholar] [CrossRef]
- Lu, H.; Zheng, Y.; Yin, W.; Tao, D.; Pesika, N.; Meng, Y.; Tian, Y. Propulsion Principles of Water Striders in Sculling Forward through Shadow Method. J. Bionic Eng. 2018, 15, 516–525. [Google Scholar] [CrossRef]
- Yin, W.; Zheng, Y.L.; Lu, H.Y.; Zhang, X.J.; Tian, Y. Three-Dimensional Topographies of Water Surface Dimples Formed by Superhydrophobic Water Strider Legs. Appl. Phys. Lett. 2016, 109, 163701. [Google Scholar] [CrossRef]
- Zheng, Y.; Lu, H.; Yin, W.; Tao, D.; Shi, L.; Tian, Y. Elegant Shadow Making Tiny Force Visible for Water-Walking Arthropods and Updated Archimedes’ Principle. Langmuir 2016, 32, 10522–10528. [Google Scholar] [CrossRef] [PubMed]
- Steinmann, T.; Arutkin, M.; Cochard, P.; Raphaël, E.; Casas, J.; Benzaquen, M. Unsteady Wave Pattern Generation by Water Striders. J. Fluid Mech. 2018, 848, 370–387. [Google Scholar] [CrossRef]
- Schneider, C.; Rasband, W.; Eliceiri, K. NIH Image to ImageJ: 25 Years of Image Analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Pearson, K.G.; Franklin, R. Characteristics of Leg Movements and Patterns of Coordination in Locusts Walking on Rough Terrain. Int. J. Robot. Res. 1984, 3, 101–112. [Google Scholar] [CrossRef]
- Binder, M.D.; Hirokawa, N.; Windhorst, U. Encyclopedia of Neuroscience; Springer: Berlin/Heidelberg, Germany, 2008. [Google Scholar] [CrossRef]
- Alexander, R.M.N. Size and Shape; Edward Arnold: London, UK, 1971. [Google Scholar]
- Mahadik, G.; Hernández-Sánchez, J.F.; Arunachalam, S.; Gallo Jr, A.; Cheng, L.; Farinha, A.S.; Thoroddsen, S.T.; Mishra, H.; Duarte, C.M. Superhydrophobicity and Size Reduction Enabled Halobates (Insecta: Heteroptera, Gerridae) to Colonize the Open Ocean. Sci. Rep. 2020, 10, 7785. [Google Scholar] [CrossRef]
- Walker, E.D.; Archer, W.E. Sequential Organization of Grooming Behaviors of the Mosquito, Aedes triseriatus. J. Insect Behav. 1988, 1, 97–109. [Google Scholar] [CrossRef]
- Hughes, G.M. The Co-Ordination of Insect Movements: II The Effect of Limb Amputation and the Cutting of Commissures in The Cockroach (Blatta Oiuentalis). J. Exp. Biol. 1957, 34, 306–333. [Google Scholar] [CrossRef]
- Noah, A.J.; Quimby, L.; Frazier, F.S.; Zill, S.N. Force Detection in Cockroach Walking Reconsidered: Discharges of Proximal Tibial Campaniform Sensilla When Body Load Is Altered. J. Neurosci. 2001, 187, 769–784. [Google Scholar] [CrossRef] [PubMed]
- Noah, J.A.; Quimby, L.; Frazier, S.F.; Zill, S.N. Sensing the Effect of Body Load in Legs: Responses of Tibial Campaniform Sensilla to Forces Applied to the Thorax in Freely Standing Cockroaches. J. Comp. Physiol. A Neuroethol. Sens. Neural Behav. Physiol. 2004, 190, 201–215. [Google Scholar] [CrossRef]
- Hu, D.L.; Prakash, M.; Chan, B.; Bush, W.M. Water-Walking Devices. Exp. Fluids 2007, 43, 769–778. [Google Scholar] [CrossRef]
- Song, Y.S.; Sitti, M. STRIDE: A Highly Maneuverable and Non-Tethered Water Strider Robot. In Proceedings of the IEEE International Conference on Robotics and Automation, Rome, Italy, 10–14 April 2007; pp. 980–984. [Google Scholar] [CrossRef]
- Alexander, R.M.N. Principles of Animal Locomotion; Princeton University Press: Princeton, NJ, USA, 2013. [Google Scholar] [CrossRef]
- Delcomyn, F. Perturbation of the Motor System in Freely Walking Cockroaches. I. Rear Leg Amputation and the Timing of Motor Activity in Leg Muscles. J. Exp. Biol. 1991, 156, 483–502. [Google Scholar] [CrossRef]
- Delcomyn, F. Perturbation of the Motor System in Freely Walking Cockroaches. II. The Timing of Motor Activity in Leg Muscles after Amputation of a Middle Leg. J. Exp. Biol. 1991, 156, 503–517. [Google Scholar] [CrossRef]
- Graham, D. The Effect of Amputation and Leg Restraint on the Free Walking Coordination of the Stick Insect Carausius morosus. J. Comp. Physiol. 1977, 116, 91–116. [Google Scholar] [CrossRef]
- Pearson, K.G.; Iles, J.F. Nervous Mechanisms Underlying Intersegmental Co-ordination of Leg Movements during Walking in the Cockroach. J. Exp. Biol. 1973, 58, 725–744. [Google Scholar] [CrossRef]
- Kong, X.Q.; Liu, J.L.; Zhang, W.J.; Qu, Y.D. Load-Bearing Ability of the Mosquito Tarsus on Water Surfaces Arising from Its Flexibility. AIP Adv. 2015, 5, 037101. [Google Scholar] [CrossRef]
- Bush, J.W.M.; Hu, D.L. Walking on Water: Biolocomotion at the Interface. Annu. Rev. Fluid Mech. 2006, 38, 339–369. [Google Scholar] [CrossRef]
- Darnhofer-Demar, B. Zur Fortbewegung des Wasserl~iufers Gerris lacustris L. auf der Wasseroberfl~che. Verh Dtsch. Zool Ges. 1968, 28, 430–439. [Google Scholar]
- Grabowska, M.; Godlewska, E.; Schmidt, J.; Daun-Gruhn, S. Quadrupedal Gaits in Hexapod Animals—Inter-Leg Coordination in Free-Walking Adult Stick Insects. J. Exp. Biol. 2012, 215, 4255–4266. [Google Scholar] [CrossRef]
- Pratt, R.Y. Striding Habits in the Gerridae. Pan-Pac.Entomol. 1938, 14, 157. [Google Scholar]
- Tseng, M.; Rowe, L. Sexual Dimorphism and Allometry in the Giant Water Strider Gigantometra gigas. Can. J. Zool. 1999, 77, 923–929. [Google Scholar] [CrossRef]
- Gao, P.; Feng, J. A Numerical Investigation of the Propulsion of Water Walkers. J. Fluid Mech. 2011, 668, 363–383. [Google Scholar] [CrossRef]
- Zill, S.N.; Keller, B.R.; Duke, E.R. Sensory Signals of Unloading in One Leg Follow Stance Onset in Another Leg: Transfer of Load and Emergent Coordination in Cockroach Walking. J. Neurophysiol. 2008, 101, 2297–2304. [Google Scholar] [CrossRef]
- Akay, T.; Ludwar, B.C.; Goritz, M.L.; Schmitz, J.; Buschges, A. Segment specificity of load signal processing depends on walking direction in the stick insect leg muscle control system. J. Neurosci. 2007, 27, 3285–3294. [Google Scholar] [CrossRef] [PubMed]
- Bässler, U. Sensory control of leg movement in the stick insect Carausius morosus. Biol. Cybern. 1977, 25, 61–72. [Google Scholar] [CrossRef] [PubMed]
- Dean, J. Control of Leg Protraction in the Stick Insect: A Targeted Movement Showing Compensation for Externally Applied Forces. J. Comp. Physiol. A 1984, 155, 771–781. [Google Scholar] [CrossRef]
- Delcomyn, F. Walking Robots and the Central and Peripheral Control of Locomotion in Insects. J. Comp. Physiol. A Sens. Neural Behav. Physiol. 1999, 185, 259–270. [Google Scholar] [CrossRef]
- Schmitz, J. Load-Compensating Reactions in the Proximal Leg Joints of Stick Insects During Standing and Walking. J. Exp. Biol. 1993, 183, 15–33. [Google Scholar] [CrossRef]
- Owaki, D.; Aonuma, H.; Sugimoto, Y.; Ishiguro, A. Leg Amputation Modifies Coordinated Activation of the Middle Leg Muscles in the Cricket Gryllus bimaculatus. Sci. Rep. 2021, 11, 1327. [Google Scholar] [CrossRef]
- Feng, X.Q.; Gao, X.F.; Wu, Z.N.; Jiang, L.; Zheng, Q.S. Superior Water Repellency of Water Strider Legs with Hierarchical Structures: Experiments and Analysis. Langmuir 2007, 23, 4892–4896. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Jiang, L. Biophysics: Water-Repellent Legs of Water Striders. Nature 2004, 432, 36. [Google Scholar] [CrossRef] [PubMed]
- Full, R.J.; Zuccarello, D.A.; Tullis, A. Effect of Variation in Form on the Cost of Terrestrial Locomotion. J. Exp. Biol. 1990, 150, 233–246. [Google Scholar] [CrossRef]
- Delcomyn, F. The Effect of Limb Amputation on Locomotion in the Cockroach Periplaneta americana. J. Exp. Biol. 1971, 54, 453–469. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, J.; Ren, L. The Terrestrial Locomotion of a Mole Cricket with Foreleg Amputation. Sci. China Technol. Sci. 2015, 58, 999–1006. [Google Scholar] [CrossRef]
- Bateman, P.W.; Fleming, P.A. Direct and indirect costs of limb autotomy in field crickets, Gryllus bimaculatus. Anim. Behav. 2005, 69, 151–159. [Google Scholar] [CrossRef]
- Zill, S.; Schmitz, J.; Büschges, A. Load Sensing and Control of Posture and Locomotion. Arthropod Struct. Dev. 2004, 33, 273–286. [Google Scholar] [CrossRef]
- Wittlinger, M.; Wehner, R.; Wolf, H. The Ant Odometer: Stepping on Stilts and Stumps. Science 2006, 312, 1965–1967. [Google Scholar] [CrossRef]
- Wittlinger, M.; Wehner, R.; Wolf, H. The Desert Ant Odometer: A Stride Integrator That Accounts for Stride Length and Walking Speed. J. Exp. Biol. 2007, 210, 198–207. [Google Scholar] [CrossRef]
- Berendes, V.; Zill, S.N.; Büschges, A.; Bockemühl, T. Speed-dependent interplay between local pattern-generating activity and sensory signals during walking in Drosophila. J. Exp. Biol. 2016, 219, 3781–3793. [Google Scholar] [CrossRef]
- Wosnitza, A.; Bockemühl, T.; Dubbert, M.; Scholz, H.; Buschges, A. Inter-Leg Coordination in the Control of Walking Speed in Drosophila. J. Exp. Biol. 2013, 216, 480–491. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.; Pham, T.H.; Nguyen, P.D.; Tran, A.D.; Ha, J.; Jablonski, P.G.; Lee, S. Locomotion and Flow Speed Preferences in Natural Habitats by Large Water Striders, Ptilomera tigrina, with Micro-Morphological Adaptations for Rowing. J. Ethol. 2022, 40, 211–221. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Symbols | Amputated Leg(s) |
|---|---|
| +/− RH | Right-hindleg |
| −/+ LF | Left-foreleg |
| −/− HL | Hindlegs pair |
| −/− ML | Midlegs pair |
| −/− FL | Forelegs pair |
| +/− RH & +/− RM | Right-hindleg and Right-midleg |
| +/− RF & +/− RM | Right-foreleg and Right-midleg |
| Grade I | Grade II | Grade III |
|---|---|---|
| +/− RH | +/− RH & −/+ LF | −/− HL |
| −/+ LF | −/− FL | −/− ML |
| +/− RM | +/− RH & +/− RM | |
| +/− RF & +/− RM |
| Animal Model | Load Value on Legs (%) | BOS (mm2) | |||||
|---|---|---|---|---|---|---|---|
| R1 | R2 | R3 | L1 | L2 | L3 | ||
| Intact * | 19 | 10 | 21 | 19 | 10 | 21 | 274 |
| +/− RH | 21 | 18 | 0 | 20 | 13 | 28 | 177 |
| −/+ LF | 30 | 10 | 22 | 0 | 11 | 27 | 288 |
| +/− RH & −/+ LF | 34 | 22 | 0 | 0 | 15 | 29 | 144 |
| +/− RM | 21 | 0 | 27 | 20 | 13 | 19 | 220 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meshkani, J.; Rajabi, H.; Kovalev, A.; Gorb, S.N. Locomotory Behavior of Water Striders with Amputated Legs. Biomimetics 2023, 8, 524. https://doi.org/10.3390/biomimetics8070524
Meshkani J, Rajabi H, Kovalev A, Gorb SN. Locomotory Behavior of Water Striders with Amputated Legs. Biomimetics. 2023; 8(7):524. https://doi.org/10.3390/biomimetics8070524
Chicago/Turabian StyleMeshkani, Javad, Hamed Rajabi, Alexander Kovalev, and Stanislav N. Gorb. 2023. "Locomotory Behavior of Water Striders with Amputated Legs" Biomimetics 8, no. 7: 524. https://doi.org/10.3390/biomimetics8070524
APA StyleMeshkani, J., Rajabi, H., Kovalev, A., & Gorb, S. N. (2023). Locomotory Behavior of Water Striders with Amputated Legs. Biomimetics, 8(7), 524. https://doi.org/10.3390/biomimetics8070524