
Forced Biomineralization: A Review





Abstract
1. Introduction
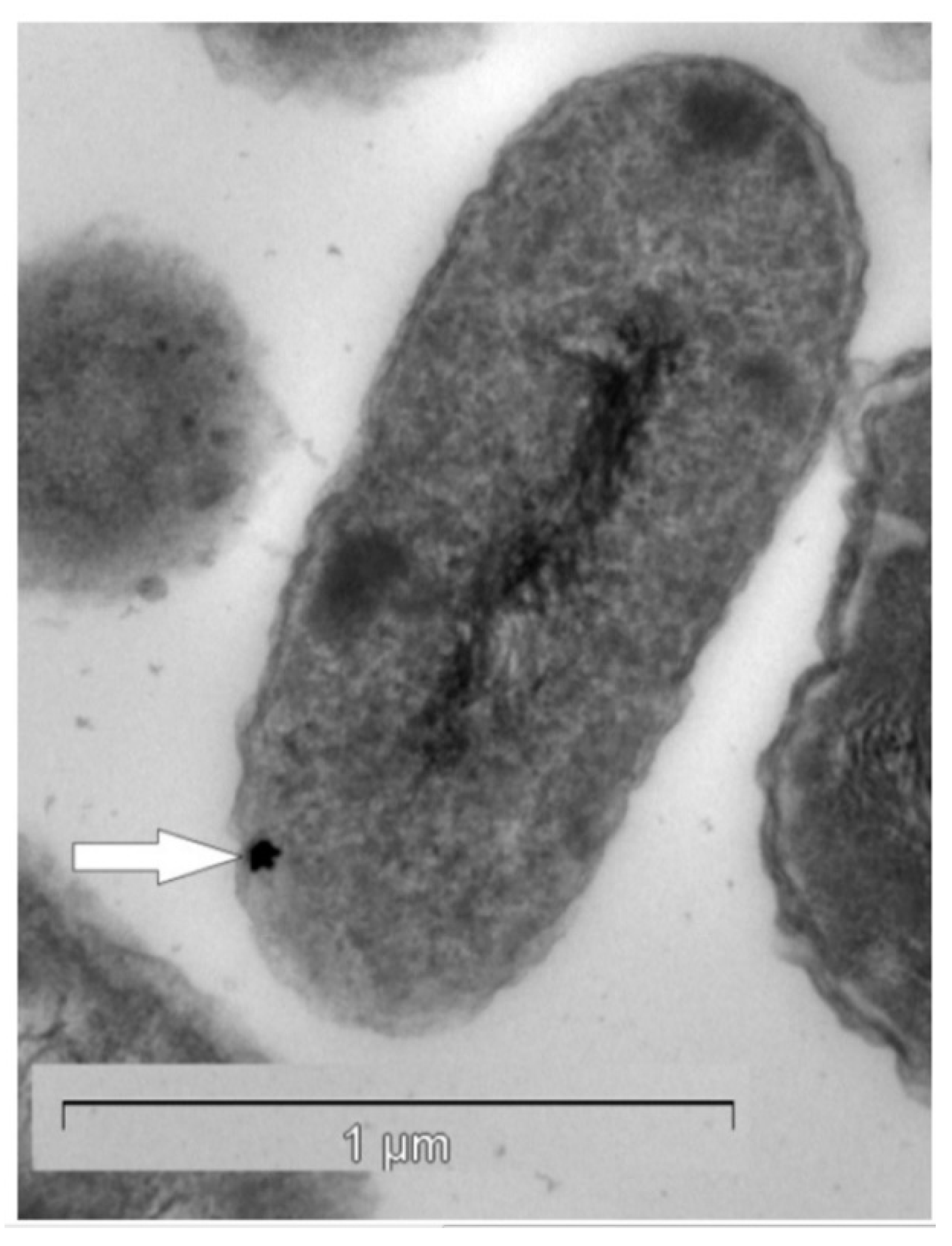
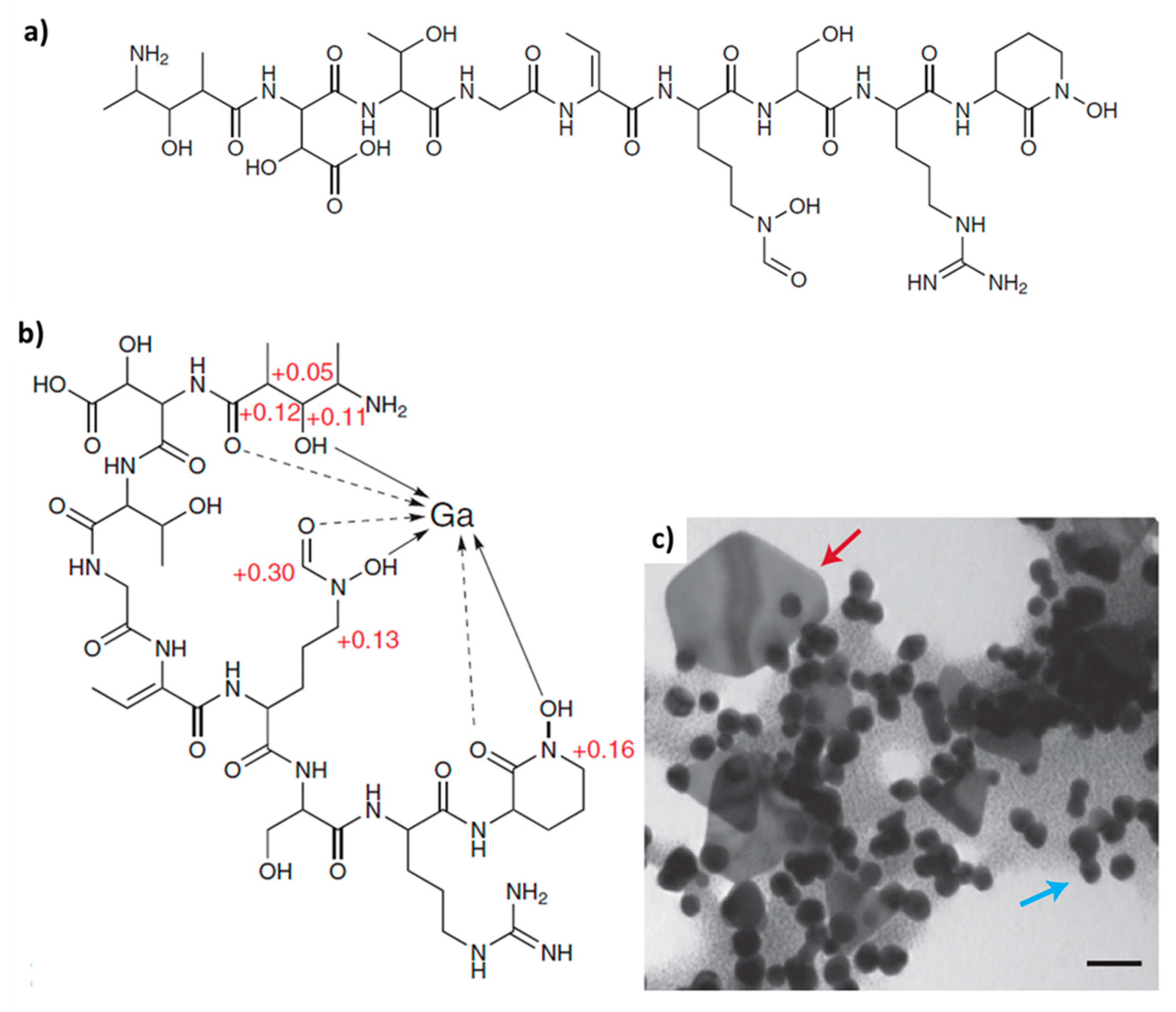
2. Biomineralization of Gold

3. Bioscorodite
- “ the bioscorodite crystal features are very similar to the mineral;
- supersaturation is controlled on a microscale by biological iron oxidation at 80 °C without the use of seed material;
- the biological oxidation does not need the use of strong chemical oxidants;
- arsenic levels of at least 1g/L can be treated (potential future applications of bioscorodite crystallization for metallurgical streams);
- crystal and agglomerates size enable an efficient solid-liquid separation.”
4. Biogenic Hydrozincite
5. Biogenic Manganese Oxides

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biomineral Name | Chemical Composition | Reference |
|---|---|---|
| Switzerite | (Mn, Fe)3(PO4)2 | [118] |
| Bixbyite | (Mn, Fe)2O3 | [118] |
| Hausmannite | Mn2+Mn4+2O4 | [111,118] |
| Pyrolusite | MnO2 | [118] |
| Manganosite | MnO | [118] |
| Romanechite | (Ba, H2O)2(Mn4+, Mn3+)5O10 | [119] |
| Rhodochrosite | MnCO3 | [118] |
| Todorokite | Mn4O7 H2O | [120,121] |
| Birnessite | Na4Mn14O27 9 H2O | [122,123] |
| Bixybyite-like | Mn2O3 | [124] |
6. Biogenic Nickel Minerals
7. Biogenic Vanadate
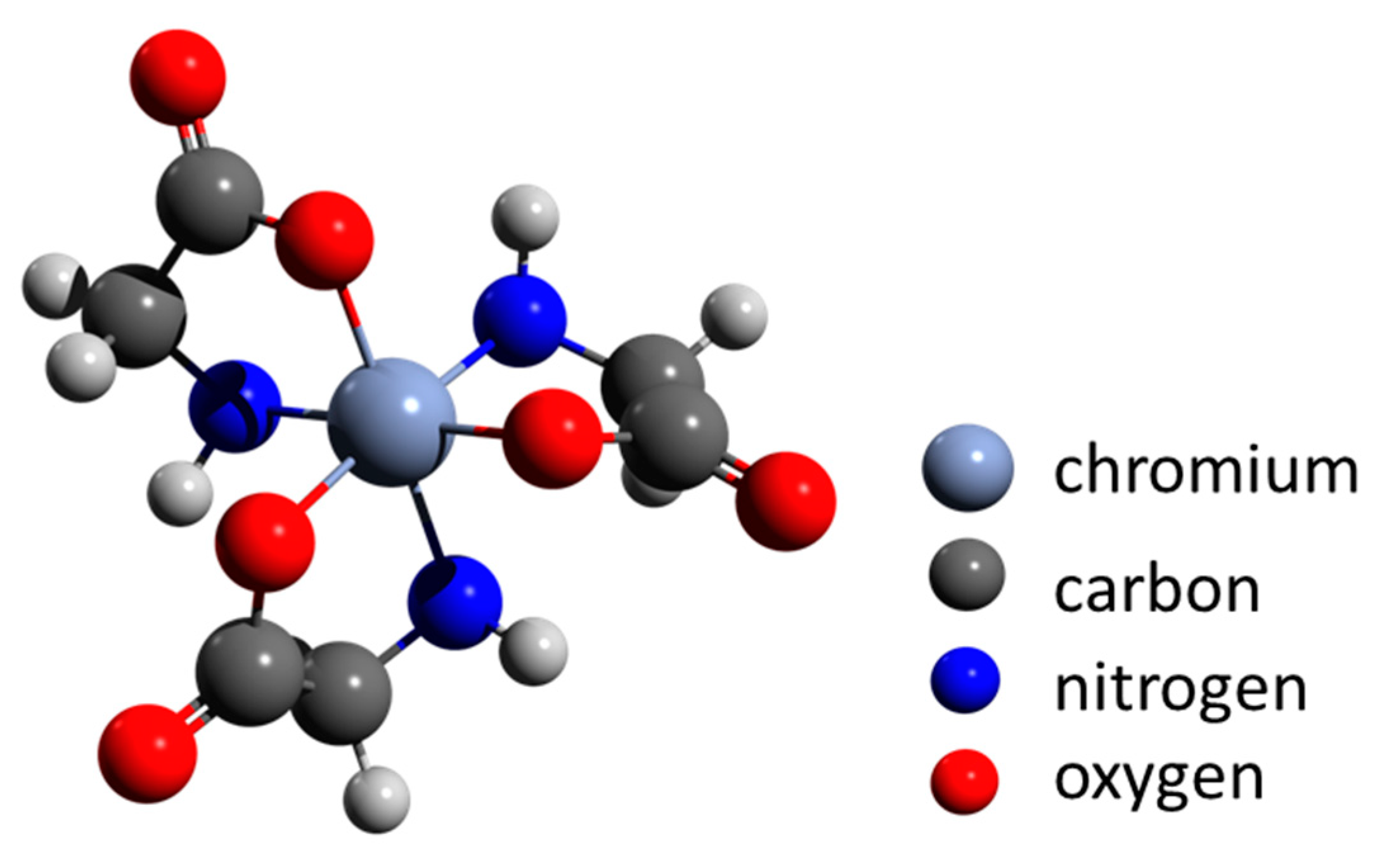
8. Biogenic Chromium Minerals
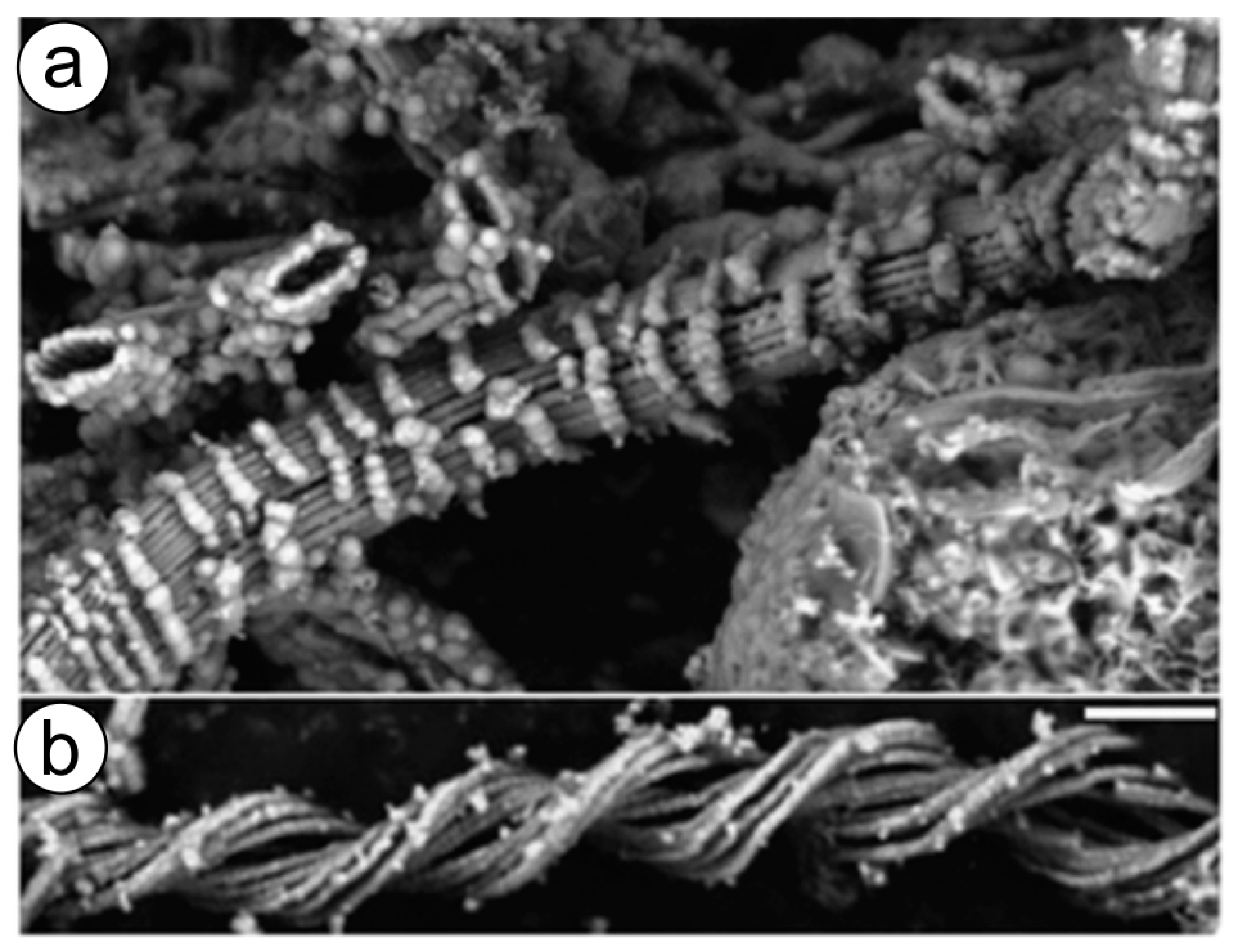
9. Iron Biomineralization and Extremophilic Organisms
10. Tellurium Biomineralization
11. Acidophilic Biomineralization as an Example of Forced Biomineralization
12. Prospects for Practical Use
- The biomineralization of iron-, silica- and calcium-based phases at extreme environmental conditions;
- The survival strategies of pro- and eukaryotes using protective advantages of biomineralization due to the functionalization of their cell envelopes;
- The mechanisms controlling fossilization, as well as exceptional preservation of organic templates which strongly bind to the mineral surface [248];
- The underlying mechanisms used by diverse extremophiles and polyextremophiles to exhibit extreme cold (cryo-), heat (thermo-), and pressure (piezo-) tolerance.
13. Outlook
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zbindn, M.; Martinez, I.; Guyot, F.; Cambon-Bonavita, M.-A.; Gaill, F. Zinc-Iron Sulphide Mineralization in Tubes of Hydrothermal Vent Worms. Eur. J. Mineral. 2001, 13, 653–658. [Google Scholar] [CrossRef]
- Lowenstam, H.A.; Weiner, S. On Biomineralization; Oxford University Press: Oxford, UK, 1989. [Google Scholar]
- Cuif, J.P.; Dauphin, Y.; Sorauf, J.E. Biominerals and Fossils Through Time; Cambridge University Press: New York, NY, USA, 2010. [Google Scholar]
- Gilbert, P.U.P.A.; Porter, S.M.; Sun, C.Y.; Xiao, S.; Gibson, B.M.; Shenkar, N.; Knoll, A.H. Biomineralization by particle attachment in early animals. Proc. Natl. Acad. Sci. USA 2019, 116, 3617659–3617665. [Google Scholar] [CrossRef] [PubMed]
- Murdock, D.J.E. The ‘biomineralization toolkit’ and the origin of animal skeletons. Biol. Rev. 2020, 95, 1372–1392. [Google Scholar] [CrossRef] [PubMed]
- Clark, M. Molecular mechanisms of biomineralization in marine invertebrates. J. Exp. Biol. 2020, 223, jeb.206961. [Google Scholar] [CrossRef] [PubMed]
- Ehrlich, H.; Koutsoukos, P.; Demadis, K.; Pokrovsky, O. Principles of demineralization: Modern strategies for the isolation of organic frameworks. Part II. Decalcification. Micron 2009, 40, 169–193. [Google Scholar] [CrossRef] [PubMed]
- Ehrlich, H.; Bazhenov, V.V.; Debitus, C.; de Voogd, N.; Galli, R.; Tsurkan, M.; Wysokowski, M.; Meissner, H.; Bulut, E.; Kaya, M.; et al. Isolation and identification of chitin from heavy mineralized skeleton of Suberea clavata (Verongida: Demospongiae: Porifera) marine demosponge. Int. J. Biol. Macromol. 2017, 104, 1706–1712. [Google Scholar] [CrossRef] [PubMed]
- Pompe, W.; Worch, H.; Habraken, W.J.E.M.; Simon, P.; Kniep, R.; Ehrlich, H.; Paufler, P. Octacalcium phosphate–a metastable mineral phase controls the evolution of scaffold forming proteins. J. Mat. Chem. B 2015, 3, 5318–5329. [Google Scholar] [CrossRef] [PubMed]
- Polishchuk, I.; Aronhime Bracha, A.; Bloch, L.; Levy, D.; Kauffmann, Y.; Burghammer, M.; Giacobbe, C.; Hendler, G.; Sun, C.Y.; Giuffre, A.J.; et al. Coherently Aligned Nanoparticles Within a Biogenic Single Crystal: A Biological Prestressing Strategy. Science 2017, 358, 1294. [Google Scholar] [CrossRef]
- Zöllner, D.; Zlotnikov, I. Biomineralized tissue formation as an archetype of ideal grain growth. Mater. Horiz. 2019, 6, 751–757. [Google Scholar] [CrossRef]
- Jantschke, A.; Pinkas, I.; Schertel, A.; Addadi, L.; Weiner, S. Biomineralization pathways in calcifying dinoflagellates: Uptake, storage in MgCaP-rich bodies and formation of the shell. Acta Biomater. 2020, 102, 427–439. [Google Scholar] [CrossRef]
- Brownlee, C.; Langer, G.; Wheeler, G.L. Coccolithophore calcification: Changing paradigms in changing oceans. Acta Biomater. 2021, 120, 4–11. [Google Scholar] [CrossRef] [PubMed]
- Ehrlich, H.; Deutzmann, R.; Capellini, E.; Koon, H.; Solazzo, C.; Yang, Y.; Ashford, D.; Thomas–Oates, J.; Lubeck, M.; Baessmann, C.; et al. Mineralization of the meter–long biosilica structures of glass sponges is template on hydroxylated collagen. Nat. Chem. 2010, 2, 1084–1088. [Google Scholar] [CrossRef]
- Ehrlich, H.; Demadis, K.; Pokrovsky, O.; Koutsoukos, P. Modern views on desilicification: Biosilica and abiotic silica dissolution in natural and artificial environments. Chem. Rev. 2010, 110, 4656–4689. [Google Scholar] [CrossRef]
- Wysokowski, M.; Jesionowski, T.; Ehrlich, H. Biosilica as source for inspiration in biological materials science. Am. Mineral. 2018, 103, 665–691. [Google Scholar] [CrossRef]
- Kopáni, M.; Hlinková, J.; Ehrlich, H.; Valigura, D.; Boča, R. Magnetic properties of iron oxides in the human Globus pallidus. J. Bioanal. Biomed. 2017, 9, 2. [Google Scholar] [CrossRef]
- McCausland, H.C.; Komeili, A. Magnetic genes: Studying the genetics of biomineralization in magnetotactic bacteria. PLoS Genet. 2020, 16, e1008499. [Google Scholar] [CrossRef]
- Ehrlich, H.; Simon, P.; Carrillo–Cabrera, W.; Bazhenovm, V.V.; Botting, J.; Ilan, M.; Ereskovsky, A.V.; Muricy, G.; Worch, H.; Mensch, A.; et al. Insights into chemistry of biological materials: Newly discovered silica–aragonite–chitin biocomposites in Demosponges. Chem. Mater. 2010, 22, 1462–1471. [Google Scholar] [CrossRef]
- Ehrlich, H.; Motylenko, M.; Sundareshwar, P.V.; Ereskovsky, A.V.; Zglobicka, I.; Noga, T.; Plocinski, T.; Tsurkan, M.; Wyroba, E.; Suski, S.; et al. Multiphase biomineralization: Enigmatic invasive siliceous diatoms produce crystalline calcite. Adv. Funct. Mater. 2016, 26, 2503–2510. [Google Scholar] [CrossRef]
- Gal, A.; Hirsch, A.; Siegel, S.; Li, C.H.; Aichmayer, B.; Politi, Y.; Fratzl, P.; Weiner, S.; Addadi, L. Plant cystoliths: A complex functional biocomposite of four distinct silica and amorphous calcium carbonate phases. Chem. Easton 2012, 18, 10262–10270. [Google Scholar] [CrossRef] [PubMed]
- Pierantoni, M.; Tenne, R.; Rephael, B.; Brumfeld, V.; van Casteren, A.; Kupczik, K.; Oron, D.; Addadi, L.; Weiner, S. Mineral Deposits in Ficus Leaves: Morphologies and Locations in Relation to Function. Plant Physiol. 2018, 176, 1751–1763. [Google Scholar] [CrossRef] [PubMed]
- Khrunyk, Y.; Lach, S.; Petrenko, I.; Ehrlich, H. Progress in Modern Marine Biomaterials Research. Mar. Drugs 2020, 18, 589. [Google Scholar] [CrossRef]
- Connors, M.; Ehrlich, H.; Hog, M.; Godeffroy, C.; Araya, S.; Kallai, I.; Gazit, D.; Boyce, M.C.; Ortiz, C. Three–dimensional structure of the shell plate assembly of the chiton Tonicella marmorea and its biomechanical consequences. J. Struct. Biol. 2012, 177, 314–328. [Google Scholar] [CrossRef] [PubMed]
- Schoeppler, V.; Gránásy, L.; Reich, E.; Poulsen, N.; de Kloe, R.; Cook, P.; Rack, A.; Pusztai, T.; Zlotnikov, I. Biomineralization as a paradigm of directional solidification: A physical model for molluscan shell ultrastructural morphogenesis. Adv. Mater. 2018, 30, 1803855. [Google Scholar] [CrossRef]
- Machałowski, T.; Wysokowski, M.; Petrenko, I.; Langer, E.; Tsurkan, D.; Jesionowski, T.; Ehrlich, H. In vivo biomimetic calcification of selected organic scaffolds using snail shell regeneration: A new methodological approach. Appl. Phys. A 2020, 126, 469. [Google Scholar] [CrossRef]
- Wysokowski, M.; Machalowski, T.; Petrenko, I.; Schimpf, C.; Rafaja, D.; Galli, R.; Ziętek, J.; Pantović, S.; Voronkina, A.; Kovalchuk, V.; et al. 3D chitin scaffolds of marine demosponge origin for biomimetic mollusk hemolymph-associated biomineralization ex-vivo. Mar. Drug 2020, 18, 123. [Google Scholar] [CrossRef] [PubMed]
- Ehrlich, H.; Martinovic, R.; Joksimovic, D.; Petrenko, I.; Schiaparelli, S.; Wysokowski, M.; Tsurkan, D.; Stelling, A.L.; Springer, A.; Gelinsky, M.; et al. Conchixes: Organic scaffolds which resemble the size and shapes of mollusks shells, their isolation and potential multifunctional applications. Appl. Phys. A 2020, 126, 562. [Google Scholar] [CrossRef]
- Kahil, K.; Varsano, N.; Sorrentino, A.; Pereiro, E.; Rez, P.; Weiner, S.; Addadi, L. Cellular pathways of calcium transport and concentration toward mineral formation in sea urchin larvae. Proc. Natl. Acad. Sci. USA 2020, 117, 30957–30965. [Google Scholar] [CrossRef] [PubMed]
- Gautron, J.; Stapane, L.; Le Roy, N.; Nys, Y.; Rodriguez-Navarro, A.B.; Hincke, M.T. Avian eggshell biomineralization: An update on its structure, mineralogy and protein tool kit. BMC Mol. Cell Biol. 2021, 22, 11. [Google Scholar]
- He, L.; Hao, Y.; Zhen, L.; Liu, H.; Shao, M.; Xu, X.; Liang, K.; Gao, Y.; Yuan, H.; Li, J.; et al. Biomineralization of dentin. J. Struct. Biol. 2019, 207, 115–122. [Google Scholar] [CrossRef]
- Amini, S.; Razi, H.; Seidel, R.; Werner, D.; White, W.T.; Weaver, J.C.; Dean, M.N.; Fratzl, P. Shape-preserving erosion controlled by the graded microarchitecture of shark tooth enameloid. Nat. Comm. 2020, 11, 5971. [Google Scholar] [CrossRef]
- Wysokowski, M.; Petrenko, I.; Galli, R.; Schimpf, C.; Rafaja, D.; Hubalkova, J.; Aneziris, C.G.; Dyshlovoy, S.; von Amsberg, G.; Meissner, H.; et al. Extreme biomineralization: The case of the hypermineralized ear bone of gray whale (Eschrichtius robustus). Appl. Phys. A 2020, 126, 727. [Google Scholar] [CrossRef]
- Wysokowski, M.; Zaslansky, P.; Ehrlich, H. Macrobiomineralogy: Insights and Enigmas in Giant Whale Bones and Perspectives for Bioinspired Materials Science ACS Biomat. Sci. Eng. 2020, 6, 5357–5367. [Google Scholar] [CrossRef] [PubMed]
- Thomas, J.; Worch, H.; Kruppke, B.; Gemming, T. Contribution to understand the biomineralization of bones. J. Bone Miner. Metab. 2020, 38, 456–468. [Google Scholar] [CrossRef] [PubMed]
- Sharma, V.; Srinivasan, A.; Nikolajeff, F.; Kumar, S. Biomineralization process in hard tissues: The interaction complexity within protein and inorganic counterparts. Acta Biomater. 2021, 120, 20–37. [Google Scholar] [CrossRef]
- Weiner, S.; Dove, P.M. An Overview of Biomineralization Processes and the Problem of the Vital Effect. Rev. Mineral. Geochem. 2003, 54, 1–29. [Google Scholar] [CrossRef]
- Benzerara, K. Biomineralization. In Encyclopedia of Astrobiology; Gargaud, M., Irvine, W.M., Amils, R., James, H., Cleaves, J., II, Pinti, D.L., Quintanilla, J.C., Rouan, D., Spohn, T., Tirard, S., et al., Eds.; Springer: Berlin/Heidelberg, Germany, 2011. [Google Scholar]
- Maleke, M.; Valverde, A.; Vermeulen, J.-G.; Cason, E.; Gomez-Arias, A.; Moloantoa, K.; Coetsee-Hugo, L.; Swart, H.; van Heerden, E.; Castillo, J. Biomineralization and Bioaccumulation of Europium by a Thermophilic Metal Resistant Bacterium. Front. Microbiol. 2019, 10, 81. [Google Scholar] [CrossRef]
- Reith, F.; Fairbrother, L.; Nolze, G.; Wilhelmi, O.; Clode, P.L.; Gregg, A.; Parsons, J.E.; Wakelin, S.A.; Pring, A.; Hough, R.; et al. Nanoparticle Factories: Biofilms Hold the Key to Gold Dispersion and Nugget Formation. Geology 2010, 38, 843–846. [Google Scholar] [CrossRef]
- Reith, F.; Lengke, M.F.; Falconer, D.; Craw, D.; Southam, G. The Geomicrobiology of Gold. ISME J. Multidiscip. J. Microb. Ecol. 2007, 1, 567–584. [Google Scholar] [CrossRef]
- Southam, G.; Lengke, M.F.; Fairbrother, L.; Reith, F. The Biogeochemistry of Gold. Elements 2009, 5, 303–307. [Google Scholar] [CrossRef]
- Bischoff, G.C. The Biological Origin of Bacterioform Gold from Australia. Neues Jahrb. Geol. Paläontol. Mon. 1997, 6, 329–338. [Google Scholar] [CrossRef]
- Jiang, X.; Zhao, C.; Fan, X.; Wu, G. Gold Biomineralization on Bacterial Biofilms for Leaching of Au3+ Damages Eukaryotic Cells. ACS Omega 2019, 4, 16667–16673. [Google Scholar] [CrossRef] [PubMed]
- Nies, D.H. Microbial Heavy Metal Resistance. Appl. Microbiol. Biotechnol. 1999, 51, 730–750. [Google Scholar] [CrossRef] [PubMed]
- Brugger, J.; Etschmann, B.; Grosse, C.; Plumridge, C.; Kaminski, J.; Paterson, D.; Shar, S.S.; Ta, C.; Howard, D.L.; de Jonge, M.D.; et al. Can Biological Toxicity Drive the Contrasting Behavior of Platinum and Gold in Surface Environments? Chem. Geol. 2013, 343, 99–110. [Google Scholar] [CrossRef]
- Usher, A.; McPhail, D.C.; Brugger, J. A Spectrophotometric Study of Aqueous Au(III) Halide-Hydroxide Complexes at 25–80 °C. Geochim. Cosmochim. Acta 2009, 73, 3359–3380. [Google Scholar] [CrossRef]
- Reith, F.; Rogers, S.L.; McPhail, D.C.; Webb, D. Biomineralization of Gold: Biofilms on Bacterioform Gold. Science 2006, 313, 233–236. [Google Scholar] [CrossRef] [PubMed]
- Reith, F.; Etschmann, B.; Grosse, C.; Moors, H.; Benotmane, M.A.; Monsieurs, P.; Grass, G.; Doonan, C.; Vogt, S.; Lai, B.; et al. Mechanisms of Gold Biomineralization in the Bacterium Cupriavidus metallidurans. Proc. Natl. Acad. Sci. USA 2009, 106, 17757–17762. [Google Scholar] [CrossRef]
- Wiesemann, N.; Mohr, J.; Grosse, C.; Herzberg, M.; Hause, G.; Reith, F.; Nies, D.H. Influence of Copper Resistance Determinants on Gold Transformation by Cupriavidus metallidurans Strain CH34. J. Bacteriol. 2013, 195, 2298–2308. [Google Scholar] [CrossRef]
- Fairbrother, L.; Brugger, J.; Shapter, J.; Laird, J.S.; Southam, G.; Reith, F. Supergene Gold Transformation: Biogenic Secondary and Nano-Particulate Gold from Arid Australia. Chem. Geol. 2012, 320–321, 17–31. [Google Scholar] [CrossRef]
- Fairbrother, L.; Etschmann, B.; Brugger, J.; Shapter, J.; Southam, G.; Reith, F. Cupriavidus metallidurans and the Biomineralization of Gold. Environ. Sci. Technol. 2013, 47, 2628–2635. [Google Scholar] [CrossRef]
- Ahmad, A.; Senapati, S.; Khan, M.I.; Kumar, R.; Sastry, M. Extracellular Biosynthesis of Monodisperse Gold Nanoparticles by a Novel Extremophilic Actinomycete, Thermomonospora sp. Langmuir 2003, 19, 3550–3553. [Google Scholar] [CrossRef]
- Maliszewska, I.; Tylus, W.; Chęcmanowski, J.; Szczygieł, B.; Pawlaczyk-Graja, I.; Pusz, W.; Baturo-Cieśniewska, A. Biomineralization of gold by Mucor plumbeus: The progress in understanding the mechanism of nanoparticles’ formation. Biotechnol. Prog. 2017, 33, 1381–1392. [Google Scholar] [CrossRef] [PubMed]
- Johnston, C.W.; Wyatt, M.A.; Li, X.; Ibrahim, A.; Shuster, J.; Southam, G.; Magarvey, N.A. Gold Biomineralization by a Metallophore from a Gold-Associated Microbe. Nat. Chem. Biol. 2013, 9, 241–243. [Google Scholar] [CrossRef]
- Lengke, M.F.; Fleet, M.E.; Southam, G. Bioaccumulation of Gold by Filamentous Cyanobacteria Between 25 and 200 °C. Geomicrobiol. J. 2006, 23, 591–597. [Google Scholar] [CrossRef]
- Lengke, M.F.; Fleet, M.E.; Southam, G. Morphology of Gold Nanoparticles Synthesized by Filamentous Cyanobacteria from gold(I)-Thiosulfate and gold(III)-Chloride Complexes. Langmuir 2006, 22, 2780–2787. [Google Scholar] [CrossRef]
- Verma, V.C.; Ahand, S.; Ulrichs, C.; Singh, S.K. Biogenic Gold Nanotriangles from Saccharomonospora sp.; an Endophytic Actinomycetes of Azadirachta indica A. Juss. Int. Nano Lett. 2013, 3, 21. [Google Scholar] [CrossRef]
- Das, S.K.; Liang, J.; Schmidt, M.; Laffir, F.; Marsili, E. Biomineralization Mechanism of Gold by Zygomycete Fungi Rhizopous oryzae. ACS Nano 2012, 6, 6165–6173. [Google Scholar] [CrossRef]
- Das, S.K.; Das, A.R.; Guha, A.K. Microbial Synthesis of Multishaped Gold Nanostructures. Small 2010, 6, 1012–1021. [Google Scholar] [CrossRef]
- Raveendran, S.; Palaninathan, V.; Nagaoka, Y.; Fukuda, T.; Iwai, S.; Higashi, T.; Mizuki, T.; Sakamoto, Y.; Mohanan, P.V.; Maekawa, T.; et al. Extremophilic Polysaccharide Nanoparticles for Cancer Nanotherapy and Evaluation of Antioxidant Properties. Int. J. Biol. Macromol. 2015, 76, 310–319. [Google Scholar] [CrossRef]
- Jones, B.; Renaut, R.W.; Rosen, M.R. Biogenicity of Gold- and Silver-Bearing Siliceous Sinters Forming in Hot (75 °C) Anaerobic Spring-Waters of Champagne Pool, Waiotapu, North Island, New Zealand. J. Geol. Soc. Lond. 2001, 158, 895–911. [Google Scholar] [CrossRef]
- Hartings, M.R.; Benjamin, N.; Briere, F.; Briscione, M.; Choudary, O.; Fisher, T.L.; Flynn, L.; Ghias, E.; Harper, M.; Khamis, N.; et al. Concurrent Zero-Dimensional and One-Dimensional Biomineralization of Gold from a Solution of Au3+ and Bovine Serum Albumin. Sci. Technol. Adv. Mater. 2013, 14, 65004. [Google Scholar] [CrossRef][Green Version]
- Chakraboroty, N.; Banerjee, A.; Lahiri, S.; Panda, A.; Ghosh, A.N.; Pal, R. Biorecovery of Gold Using Cyanobacteria and an Eukaryotic Alga with Special Reference to Nanogold Formation; A Novel Phenomenon. J. Appl. Phycol. 2009, 21, 145–152. [Google Scholar] [CrossRef]
- He, S.; Guo, Z.; Zhang, Y.; Zhang, S.; Wang, J.; Gu, N. Biosynthesis of Gold Nanoparticles Using the Bacteria Rhodopseudomonas capsulata. Mater. Lett. 2007, 61, 3984–3987. [Google Scholar] [CrossRef]
- Du, L.; Xian, L.; Feng, J.-X. Rapid Extra-/Intracellular Biosynthesis of Gold Nanoparticles by the Fungus Penicillium sp. J. Nanoparticle Res. 2011, 13, 921–930. [Google Scholar] [CrossRef]
- Roh, Y.; Kang, S.; Park, B.; Kim, Y. Microbially Induced Precipitation of Gold(0) Nanoparticles. J. Nanosci. Nanotechnol. 2015, 15, 408–411. [Google Scholar] [CrossRef]
- Schwartz-Duval, A.S.; Konopka, C.J.; Moitra, P.; Daza, E.A.; Srivastava, I.; Johnson, E.V.; Kampert, T.L.; Fayn, S.; Haran, A.; Dobrucki, L.W.; et al. Intratumoral generation of photothermal gold nanoparticles through a vectorized biomineralization of ionic gold. Nat. Comm. 2020, 11, 4530. [Google Scholar] [CrossRef]
- Shamaila, S.; Zafar, N.; Riaz, S.; Sharif, R.; Nazir, J.; Naseem, S. Gold Nanoparticles: An Efficient Antimicrobial Agent Against Enteric Bacterial Human Pathogen. Nanomaterials 2016, 6, 71. [Google Scholar] [CrossRef]
- Li, X.; Robinson, S.M.; Gupta, A.; Saha, K.; Jiang, Z.; Moyano, D.F.; Sahar, A.; Riley, M.A.; Rotello, V.M. Functional Gold Nanoparticles as Potent Antimicrobial Agents against Multi-Drug-Resistant Bacteria. ACS Nano 2014, 8, 10682–10686. [Google Scholar] [CrossRef]
- Rai, A.; Prabhuneb, A.; Perry, C.C. Antibiotic Mediated Synthesis of Gold Nanoparticles with Potent Antimicrobial Activity and Their Application in Antimicrobial Coatings. J. Mater. Chem. 2010, 20, 6789–6798. [Google Scholar] [CrossRef]
- Gwynne, P. Microbiology: There’s Gold in Them There Bugs. Nature 2013, 495, S12–S13. [Google Scholar] [CrossRef]
- Houngaloune, S. On-Site Synthesis of Schwertmannite and Its Application for Arsenic Immobilization at Copper Mines. Ph.D. Thesis, Hokkaido University, Hokkaido, Japan, 2015. [Google Scholar]
- Rodriguez-Freire, L.; Sierra-Alvarez, R.; Root, R.; Chorover, J.; Field, J.A. Biomineralization of Arsenate to Arsenic Sulfides Is Greatly Enhanced at Mildly Acidic Conditio. Water Res. 2015, 66, 242–253. [Google Scholar] [CrossRef]
- Ledbetter, R.N.; Connon, S.A.; Neal, A.L.; Dohnalkova, A.; Magnuson, T.S. Biogenic Mineral Production by a Novel Arsenic-Metabolizing Thermophilic Bacterium from the Alvord Basin, Oregon. Appl. Environ. Microbiol. 2007, 73, 5928–5936. [Google Scholar] [CrossRef]
- Newman, D.K.; Beveridge, T.J.; Morel, F.M.M. Precipitation of Arsenic Trisulfide by Desulfotomaculum auripigmentum. Appl. Environ. Microbiol. 1997, 63, 2022–2028. [Google Scholar] [CrossRef]
- Lee, J.-H.; Kim, M.-G.; Yoo, B.; Myung, N.V.; Maeng, J.; Lee, T.; Dohnalkova, A.C.; Fredrickson, J.K.; Sadowsky, M.J.; Hur, H.-G. Biogenic Formation of Photoactive Arsenic-Sulfide Nanotubes by Shewanella sp. Strain HN-41. Proc. Natl. Acad. Sci. USA 2007, 104, 20410–20415. [Google Scholar] [CrossRef]
- Gonzalez-Contreras, P.; Weijma, J.; Van Der Weijden, R.; Buisman, C.J.N. Biogenic Scorodite Crystallization by Acidianus sulfidivorans for Arsenic Removal. Environ. Sci. Technol. 2010, 44, 675–680. [Google Scholar] [CrossRef]
- Gonzalez-Contreras, P.; Weijma, J.; Buisman, C.J.N. Bioscorodite Crystallization in an Airlift Reactor for Arsenic Removal. Cryst. Growth Des. 2012, 12, 2699–2706. [Google Scholar] [CrossRef]
- Gonzalez-Contreras, P.; Weijma, J.; Buisman, C.J.N. Continuous Bioscorodite Crystallization in CSTRs for Arsenic Removal and Disposal. Water Res. 2012, 46, 5883–5892. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Contreras, P. BIOSCORODITE: Biological Crystallization of Scorodite for Arsenic Removal; Wageningen University: Wageningen, The Netherlands, 2012. [Google Scholar]
- Okibe, N.; Morishita, S.; Tanaka, M.; Hirajima, T.; Sasaki, K. Effect of Cu(II) on Bio-Scorodite Crystallization Using Acidianus brierleyi. Adv. Mater. Res. 2015, 1130, 101–104. [Google Scholar] [CrossRef]
- Okibe, N.; Morishita, S.; Tanaka, M.; Sasaki, K.; Hirajima, T.; Hatano, K.; Ohata, A. Bioscorodite Crystallization Using Acidianus brierleyi Effects Caused by Cu(II) Present in As(III)-Bearing Copper Refinery Wastewaters. Hydrometallurgy 2016, 168, 121–126. [Google Scholar] [CrossRef]
- Tanaka, M.; Okibe, N. Factors to Enable Crystallization of Environmentally Stable Bioscorodite from Dilute As(III)-Contaminated Waters. Minerals 2018, 8, 23. [Google Scholar] [CrossRef]
- Tec-Caamal, E.N.; Rodríguez-Vázquez, R.; Aguilar-López, R. Kinetic analysis of arsenic and iron oxidation by Acidianus brierleyi for biogenic scorodite formation. Chem. Pap. 2019, 73, 811–820. [Google Scholar] [CrossRef]
- Admiraal, W.; Blanck, H.; Buckert-De Jong, M.; Guasch, H.; Ivorra, N.; Lehmann, V.; Nyström, B.A.H.; Paulsson, M.; Sabater, S. Short-Term Toxicity of Zinc to Microbenthic Algae and Bacteria in a Metal Polluted Stream. Water Res. 1999, 33, 1989–1996. [Google Scholar] [CrossRef]
- Eisler, R. Zinc Hazards to Fish, Wildlife, and Invertebrates: A Synoptic Review; U.S. Department of the Interior, Fish and Wildlife Service: Laurel, MI, USA, 1993. [Google Scholar]
- Mangold, S.; Potrykus, J.; Björn, E.; Lövgren, L.; Dopson, M. Extreme Zinc Tolerance in Acidophilic Microorganisms from the Bacterial and Archaeal Domains. Extremophiles 2013, 17, 75–85. [Google Scholar] [CrossRef]
- Ghose, S. The Crystal Structure of Hydrozincite, Zn5(OH)6(CO3)2. Acta Crystallogr. 1964, 17, 1051–1057. [Google Scholar] [CrossRef]
- Zuddas, P.; Podda, F. Variations in Physicochemical Properties of Water Associated with Bio-Precipitation of Hydrozincite [Zn5(OH)6(CO3)2] in the Waters of Rio Naracauli, Sardinia (Italy). Appl. Geochem. 2005, 20, 507–517. [Google Scholar] [CrossRef]
- Medas, D.; Cidu, R.; Lattanzi, P.; Podda, F.; Wanty, R.B.; De Giudici, G. Hydrozincite Seasonal Precipitation at Naracauli (Sardinia-Italy): Hydrochemical Factors and Morphological Features of the Biomineralization Process. Appl. Geochem. 2012, 27, 1814–1820. [Google Scholar] [CrossRef]
- Medas, D.; Lattanzi, P.; Podda, F.; Meneghini, C.; Trapananti, A.; Sprocati, A.; Casu, M.A.; Musu, E.; De Giudici, G. The Amorphous Zn Biomineralization at Naracauli Stream, Sardinia: Electron Microscopy and X-Ray Absorption Spectroscopy. Environ. Sci. Pollut. Res. 2014, 21, 6775–6782. [Google Scholar] [CrossRef]
- Podda, F.; Zuddas, P.; Minacci, A.; Pepi, M.; Baldi, F. Heavy Metal Coprecipitation with Hydrozincite [Zn5(OH)6(CO3)2] from Mine Waters Caused by Photosynthetic Microorganisms. Appl. Environ. Microbiol. 2000, 66, 5092–5098. [Google Scholar] [CrossRef]
- De Giudici, G.; Wanty, R.B.; Podda, F.; Kimball, B.A.; Verplanck, P.L.; Lattanzi, P.; Cidu, R.; Medas, D. Quantifying Biomineralization of Zinc in the Rio Naracauli (Sardinia, Italy), Using a Tracer Injection and Synoptic Sampling. Chem. Geol. 2014, 384, 110–119. [Google Scholar] [CrossRef]
- Wanty, R.B.; Podda, F.; De Giudici, G.; Cidu, R.; Lattanzi, P. Zinc Isotope and Transition-Element Dynamics Accompanying Hydrozincite Biomineralization in the Rio Naracauli, Sardinia, Italy. Chem. Geol. 2013, 337–338, 1–10. [Google Scholar] [CrossRef]
- Podda, F.; Medas, D.; De Giudici, G.; Ryszka, P.; Wolowski, K.; Turnau, K. Zn Biomineralization Processes and Microbial Biofilm in a Metal-Rich Stream (Naracauli, Sardinia). Environ. Sci. Pollut. Res. 2014, 21, 6793–6808. [Google Scholar] [CrossRef]
- De Giudici, G.; Podda, F.; Sanna, R.; Musu, E.; Tombolini, R.; Cannas, C.; Musinu, A.; Casu, M. Structural Properties of Biologically Controlled Hydrozincite: An HRTEM and NMR Spectroscopic Study. Am. Mineral. 2009, 94, 1698–1706. [Google Scholar] [CrossRef]
- Ngwenya, B.T.; Magennis, M.; Podda, F.; Gromov, A. Self-Preservation Strategies during Bacterial Biomineralization with Reference to Hydrozincite and Implications for Fossilization of Bacteria. J. R. Soc. Interface 2014, 11, 20140845. [Google Scholar] [CrossRef]
- Labrenz, M.; Druschel, G.K.; Thomsen-Ebert, T.; Gilbert, B.; Welch, S.; Kemner, K.M.; Logan, G.A.; Summons, R.E.; De Stasio, G.; Bond, P.L.; et al. Formation of Sphalerite (ZnS) Deposits in Natural Biofilms of Sulfate-Reducing Bacteria. Science 2000, 290, 1744–1747. [Google Scholar] [CrossRef]
- Moon, J.-W.; Phelps, T.J.; Fitzgerald, C.L., Jr.; Lind, R.F.; Elkins, J.G.; Jang, G.G.; Joshi, P.C.; Kidder, M.; Armstrong, B.L.; Watkins, T.R.; et al. Manufacturing Demonstration of Microbially Mediated Zinc Sulfide Nanoparticles in Pilot-Plant Scale Reactors. Appl. Microbiol. Biotechnol. 2016, 100, 7921–7931. [Google Scholar] [CrossRef]
- Medas, D.; Carlomagno, I.; Meneghini, C.; Aquilanti, G.; Araki, T.; Bedolla, D.E.; Buosi, C.; Casu, M.A.; Gianoncelli, A.; Kuncser, A.C.; et al. Zinc incorporation in marine bivalve shells grown in mine-polluted seabed sediments: A case study in the Malfidano mining area (SW Sardinia, Italy). Environ. Sci. Pollut. Res. 2018, 25, 36645–36660. [Google Scholar] [CrossRef]
- Ehrlich, H.L. Manganese as an Energy Source for Bacteria. In Environmental Biogeochemistry; Nriagu, J.O., Ed.; Ann Arbor Science: Ann Arbor, MI, USA, 1976; Volume 2, pp. 633–644. [Google Scholar]
- Ehrlich, H.L. Geomicrobiology of Manganese. In Geomicrobiology; CRC Press: Boca Raton, FL, USA, 1996; pp. 429–528. [Google Scholar]
- Skinner, H.C.W.; Fitzpatrick, R.W. Biomineralization: Processes of Iron and Maganese: Modern and Ancient Environments; Catena Verlag: Drestedt, Germany, 1992; Volume 21. [Google Scholar]
- Tebo, B.M.; Johnson, H.A.; McCarthy, J.K.; Templeton, A.S. Geomicrobiology of manganese(II) Oxidation. Trends Microbiol. 2005, 13, 421–428. [Google Scholar] [CrossRef]
- Spiro, T.G.; Bargar, J.R.; Sposito, G.; Tebo, B.M. Bacteriogenic Manganese Oxides. Acc. Chem. Res. 2010, 43, 2–9. [Google Scholar] [CrossRef]
- Droz, B.; Dumas, N.; Duckworth, O.W.; Pena, J. A Comparison of the Sorption Reactivity of Bacteriogenic and Mycogenic Mn Oxide Nanoparticles. Environ. Sci. Technol. 2015, 49, 4200–4208. [Google Scholar] [CrossRef]
- Blöthe, M.; Wegorzewski, A.; Müller, C.; Simon, F.; Kuhn, T.; Schippers, A. Manganese-Cycling Microbial Communities Inside Deep-Sea Manganese Nodules. Environ. Sci. Technol. 2015, 49, 7692–7700. [Google Scholar] [CrossRef]
- Tu, J.; Yang, Z.; Hu, C.; Qu, J. Characterization and Reactivity of Biogenic Manganese Oxides for Ciprofloxacin Oxidation. J. Environ. Sci. 2014, 26, 1154–1161. [Google Scholar] [CrossRef]
- Keim, C.N.; Nalini, H.A.; Lena, J.C.D. Manganese Oxide Biominerals from Freshwater Environments in Quadrilatero Ferrifero, Minas Gerais, Brazil. Geomicrobiol. J. 2014, 31, 37–41. [Google Scholar] [CrossRef]
- Zerfass, C.; Christie-Oleza, J.A.; Soyer, O.S. Manganese Oxide Biomineralization Provides Protection against Nitrite Toxicity in a Cell-Density-Dependent Manner. Appl. Environ. Microbiol. 2019, 85, e02129-18. [Google Scholar] [CrossRef] [PubMed]
- Brouwers, G.J.; Vijgenboom, E.; Corstjens, P.; De Vrind, J.P.M.; de Vrind-de Jong, E.W. Bacterial Mn2+ Oxidizing Systems and Multicopper Oxidases: An Overview of Mechanisms and Functions. Geomicrobiol. J. 2000, 17, 1–24. [Google Scholar]
- Rosson, R.A.; Nealson, K.H. Manganese Binding and Oxidation by Spores of a Marine Bacillus. J. Bacteriol. 1982, 151, 1027–1034. [Google Scholar] [CrossRef]
- De Vrind, J.P.; de Vrind-de Jong, E.W.; de Voogt, J.W.; Westbroek, P.; Boogerd, F.C.; Rosson, R.A. Manganese Oxidation by Spores and Spore Coats of a Marine Bacillus Species. Appl. Environ. Microbiol. 1986, 52, 1096–1100. [Google Scholar] [CrossRef] [PubMed]
- Mann, S.; Sparks, N.H.; Scott, G.H.; de Vrind-de Jong, E.W. Oxidation of Manganese and Formation of Mn3O4 (Hausmannite) by Spore Coats of a Marine Bacillus sp. Appl. Environ. Microbiol. 1988, 54, 2140–2143. [Google Scholar] [CrossRef] [PubMed]
- Hastings, D.; Emerson, S. Oxidation of Manganese by Spores of a Marine Bacillus: Kinetic and Thermodynamic Considerations. Geochim. Cosmochim. Acta 1986, 50, 1819–1824. [Google Scholar] [CrossRef]
- Butterfield, C.N.; Soldatova, A.V.; Lee, S.-W.; Spiro, T.G.; Tebo, B.M. Mn(II,III) Oxidation and MnO2 Mineralization by an Expressed Bacterial Multicopper Oxidase. Proc. Natl. Acad. Sci. USA 2013, 110, 11731–11735. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Liu, D.; Jia, Z.; Csetenyi, L.; Gadd, G.M. Fungal Biomineralization of Manganese as a Novel Source of Electrochemical Materials. Curr. Biol. 2016, 26, 950–955. [Google Scholar] [CrossRef]
- Parikh, S.J.; Chorover, J. FTIR Spectroscopic Study of Biogenic Mn-Oxide Formation by Pseudomonas Putida GB-1. Geomicrobiol. J. 2005, 22, 207–218. [Google Scholar] [CrossRef]
- Ivarsson, M.; Broman, C.; Gustafsson, H.; Holm, N.G. Biogenic Mn-Oxides in Subseafloor Basalts. PLoS ONE 2015, 10, e0128863. [Google Scholar] [CrossRef]
- Akob, D.M.; Bohu, T.; Beyer, A.; Schäffner, F.; Händel, M.; Johnson, C.A.; Merten, D.; Büchel, G.; Totsche, K.U.; Küsel, K. Identification of Mn(II)-Oxidizing Bacteria from a Low-pH Contaminated Former Uranium Mine. Appl. Environ. Microbiol. 2014, 80, 5086–5097. [Google Scholar] [CrossRef]
- Villalobos, M.; Toner, B.; Bargar, J.; Sposito, G. Characterization of the Manganese Oxide Produced by Pseudomonas putida Strain MnB1. Geochim. Cosmochim. Acta 2003, 67, 2649–2662. [Google Scholar] [CrossRef]
- Saratovsky, I.; Wightman, P.G.; Pastén, P.A.; Gaillard, J.F.; Poeppelmeier, K.R. Manganese Oxides: Parallels between Abiotic and Biotic Structures. J. Am. Chem. Soc. 2006, 128, 11188–11198. [Google Scholar] [CrossRef]
- Hosseinkhani, B.; Emtiazi, G. Synthesis and Characterization of a Novel Extracellular Biogenic Manganese Oxide (Bixbyite-like Mn2O3) Nanoparticle by Isolated Acinetobacter sp. Curr. Microbiol. 2011, 63, 300–305. [Google Scholar] [CrossRef] [PubMed]
- Bohu, T.; Akob, D.M.; Abratis, M.; Lazar, C.S.; Küsel, K. Biological Low pH Mn(II) Oxidation in a Manganese Deposit Influenced by Metal-Rich Groundwater. Appl. Environ. Microbiol. 2016, 82. [Google Scholar] [CrossRef]
- Greene, A.C.; Madgwick, J.C. Microbial Formation of Manganese Oxides. Appl. Environ. Microbiol. 1991, 57, 1114–1120. [Google Scholar] [CrossRef]
- Takeda, M.; Kamagata, Y.; Ghiorse, W.C.; Hanada, S.; Koizumi, J.I. Caldimonas manganoxidans gen. nov., sp. nov., a poly(3-Hydroxybutyrate)-Degrading, Manganese-Oxidizing Thermophile. Int. J. Syst. Evol. Microbiol. 2002, 52, 895–900. [Google Scholar]
- Dick, G.J.; Lee, Y.E.; Tebo, B.M. Manganese (II)-Oxidizing Bacillus Spores in Guaymas Basin Hydrothermal Sediments and Plumes. Appl. Environ. Microbiol. 2006, 72, 3184–3190. [Google Scholar] [CrossRef] [PubMed]
- Galezowski, L.; Recham, N.; Larcher, D.; Miot, J.; Skouri-Panet, F.; Guyot, F. Microbially Induced Mineralization of Layered Mn Oxides Electroactive in Li Batteries. Front. Microbiol. 2020, 11, 2031. [Google Scholar] [CrossRef] [PubMed]
- Macomber, L.; Hausinger, R.P. Mechanisms of Nickel Toxicity in Microorganisms. Metallomics 2011, 3, 1153–1162. [Google Scholar] [CrossRef]
- Amoroso, M.J.; Schubert, D.; Mitscherlich, P.; Schumann, P.; Kothe, E. Evidence for High Affinity Nickel Transporter Genes in Heavy Metal Resistant Streptomyces Spec. J. Basic Microbiol. 2000, 40, 295–301. [Google Scholar] [CrossRef]
- Schmidt, A.; Haferburg, G.; Sineriz, M.; Merten, D.; Büchel, G.; Kothe, E. Heavy Metal Resistance Mechanisms in Actinobacteria for Survival in AMD Contaminated Soils. Chemie Der Erde-Geochem. 2005, 65, 131–144. [Google Scholar] [CrossRef]
- Haferburg, G.; Kloess, G.; Schmitz, W.; Kothe, E. “Ni-Struvite”—A New Biomineral Formed by a Nickel Resistant Streptomyces acidiscabies. Chemosphere 2008, 72, 517–523. [Google Scholar] [CrossRef]
- Sar, P.; Kazy, S.K.; Singh, S.P. Intracellular Nickel Accumulation by Pseudomonas aeruginosa and Its Chemical Nature. Lett. Appl. Microbiol. 2001, 32, 257–261. [Google Scholar] [CrossRef]
- Evans, H.T.; White, J.S. The Colorful Vanadium Minerals: A Brief Review and a New Classification. Mineral. Rec. 1987, 18, 333–340. [Google Scholar]
- Ceci, Y.J.; Kierans, M.; Hillier, S.; Pendlowski, H.; Gray, N.; Persiani, A.M.; Gadd, G.M. Transformation of Vanadinite (Pb5(VO4)3Cl) by Fungi. Environ. Microbiol. 2015, 17, 2018–2034. [Google Scholar] [CrossRef] [PubMed]
- Crans, D.C.; Amin, S.S.; Keramidas, A.D. Chemistry of Relevance to Vanadium in the Environment. In Vanadium in the Environment. Part 1: Chemistry and Biochemistry; Nriagu, J.O., Ed.; John Wiley & Sons, Inc.: New York, NY, USA, 1998; pp. 73–95. [Google Scholar]
- Lyalikova, N.N.; Yurkova, N.A. Role of Microorganisms in Vanadium Concentration and Dispersion. Geomicrobiol. J. 1992, 10, 15–26. [Google Scholar] [CrossRef]
- Zhang, J.; Dong, H.; Zhao, L.; McCarrick, R.; Agrawal, A. Microbial Reduction and Precipitation of Vanadium by Mesophilic and Thermophilic Methanogens. Chem. Geol. 2014, 370, 29–39. [Google Scholar] [CrossRef]
- Xu, Y.H.; Brandl, H.; Osterwalder, S.; Elzing, E.J.; Huang, J.H. Vanadium-basidiomycete fungi interaction and its impact on vanadium biogeochemistry. Environ. Int. 2019, 130, 104891. [Google Scholar] [CrossRef] [PubMed]
- Coker, V.S.; van der Laan, G.; Telling, N.D.; Lloyd, J.R.; Byrne, J.M.; Arenholz, E.; Pattrick, R.A.D. Bacterial production of vanadium ferrite spinel (Fe,V)3O4 nanoparticles. Mineral. Mag. 2020, 84, 554–562. [Google Scholar] [CrossRef]
- Baillet, F.; Magnin, J.P.; Cheruy, A.; Ozil, P. Chromium Precipitation by the Acidophilic Bacterium Thiobacillus ferrooxidans. Biotechnol. Lett. 1998, 20, 95–99. [Google Scholar] [CrossRef]
- Narayani, M.; Shetty, K.V. Chromium-Resistant Bacteria and Their Environmental Condition for Hexavalent Chromium Removal: A Review. Crit. Rev. Environ. Sci. Technol. 2013, 43, 955–1009. [Google Scholar] [CrossRef]
- Thatoi, H.; Das, S.; Mishra, J.; Rath, B.P.; Das, N. Bacterial Chromate Reductase, a Potential Enzyme for Bioremediation of Hexavalent Chromium: A Review. J. Environ. Manag. 2014, 146, 383–399. [Google Scholar] [CrossRef]
- Joutey, N.T.; Sayel, H.; Bahafid, W.; Ghachtouli, N.E. Mechanisms of Hexavalent Chromium Resistance and Removal by Microorganisms. Rev. Environ. Contam. Toxicol. 2015, 233, 45–69. [Google Scholar]
- Zhang, C.; Liu, S.; Logan, J.; Mazumder, R.; Phelps, T.J. Enhancement of Fe(III), Co(III), and Cr(VI) Reduction at Elevated Temperatures and by a Thermophilic Bacterium. Appl. Biochem. Biotechnol. 1996, 57–58, 923–932. [Google Scholar] [CrossRef]
- Kashefi, K.; Holmes, D.E.; Reysenbach, A.L.; Lovley, D.R. Use of Fe(III) as an Electron Acceptor to Recover Previously Uncultured Hyperthermophiles: Isolation and Characterization of Geothermobacterium ferrireducens gen. nov.; sp. nov. Appl. Environ. Microbiol. 2002, 68, 1735–1742. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Pan, H.; Xiao, T.; Barbier, G.; Wang, Z.; Yue, H.; Sun, S.; Nitsche, S.; Bernadac, A.; Pradel, N.; et al. Isolation and Characterization of Novel Marine Roseobacter Clade Members Producing Unique Intracellular Chromium-Rich Aggregates. Res. Microbiol. 2006, 157, 714–719. [Google Scholar] [CrossRef] [PubMed]
- Mohite, P.T.; Kumar, A.R.; Zinjarde, S.S. Biotransformation of Hexavalent Chromium into Extracellular chromium(III) Oxide Nanoparticles Using Schwanniomyces occidentalis. Biotechnol. Lett. 2016, 38, 441–446. [Google Scholar] [CrossRef] [PubMed]
- Jobby, R.; Jha, P.; Gupta, A.; Gupte, A.; Desai, N. Biotransformation of chromium by root nodule bacteria Sinorhizobium sp. SAR1. PLoS ONE 2019, 14, e0219387. [Google Scholar]
- Cheng, Y.; Xu, X.; Lin, W.; Han, R.; Liu, M. Transformation from Organo-Cr(III) to Trivalent Chromium Mineral (Guyanaite/Grimaldiite) and Its Environmental Implication. Geomicrobiol. J. 2016, 33, 948–953. [Google Scholar] [CrossRef]
- Gajdardziska-Josifovska, M.; McClean, R.G.; Schofield, M.A.; Sommer, C.V.; Kean, W.F. Discovery of Nanocrystalline Botanical Magnetite. Eur. J. Mineral. 2001, 13, 863–870. [Google Scholar] [CrossRef]
- Bazylinski, D.A.; Frankel, R.B. Magnetosome Formation in Prokaryotes. Nat. Rev. Microbiol. 2004, 2, 217–230. [Google Scholar] [CrossRef] [PubMed]
- Bazylinski, D.A.; Frankel, R.B.; Konhauser, K.O. Modes of Biomineralization of Magnetite by Microbes. Geomicrobiol. J. 2007, 24, 465–475. [Google Scholar] [CrossRef]
- Ledderhose, G.; Glykosamin, U. Zeitschrift für Physiol. Chemie 1880, 4, 139–159. [Google Scholar]
- Lower, B.H.; Bazylinski, D.A. The Bacterial Magnetosome: A Unique Prokaryotic Organelle. J. Mol. Microbiol. Biotechnol. 2013, 23, 63–80. [Google Scholar] [CrossRef]
- Uebe, R.; Schüler, D. Magnetosome Biogenesis in Magnetotactic Bacteria. Nat. Rev. Microbiol. 2016, 14, 621–637. [Google Scholar] [CrossRef]
- Barber-Zucker, S.; Zarivach, R. A Look into the Biochemistry of Magnetosome Biosynthesis in Magnetotactic Bacteria. ACS Chem. Biol. 2017, 12, 13–22. [Google Scholar] [CrossRef]
- Miot, J.; Benzerara, K.; Morin, G.; Kappler, A.; Bernard, S.; Obst, M.; Férard, C.; Skouri-Panet, F.; Guigner, J.M.; Posth, N.; et al. Iron Biomineralization by Anaerobic Neutrophilic Iron-Oxidizing Bacteria. Geochim. Cosmochim. Acta 2009, 73, 696–711. [Google Scholar] [CrossRef]
- Chan, C.S.; Fakra, S.C.; Edwards, D.C.; Emerson, D.; Banfield, J.F. Iron Oxyhydroxide Mineralization on Microbial Extracellular Polysaccharides. Geochim. Cosmochim. Acta 2009, 73, 3807–3818. [Google Scholar] [CrossRef]
- Krepski, S.T.; Hanson, T.E.; Chan, C.S. Isolation and Characterization of a Novel Biomineral Stalk-Forming Iron-Oxidizing Bacterium from a Circumneutral Groundwater Seep. Environ. Microbiol. 2012, 14, 1671–1680. [Google Scholar] [CrossRef] [PubMed]
- Miot, J.; Jézéquel, D.; Benzerara, K.; Cordier, L.; Rivas-Lamelo, S.; Skouri-Panet, F.; Férard, C.; Poinsot, M.; Duprat, E. Mineralogical Diversity in Lake Pavin: Connections with Water Column Chemistry and Biomineralization Processes. Minerals 2016, 6, 24. [Google Scholar] [CrossRef]
- Ehrenberg, C.G. Vorläufige Mitteilungen Über Das Wirkliche Vorkommen Fossiler Infusorien Und Ihre Große Verbreitung. Poggendorffs Ann. Phys. Chem. 1836, 38, 213–227. [Google Scholar] [CrossRef]
- Pringsheim, E.G. Iron Bacteria. Biol. Rev. Camb. Philos. Soc. 1949, 24, 200–245. [Google Scholar] [CrossRef] [PubMed]
- Fleming, E.J.; Davis, R.E.; Mcallister, S.M.; Chan, C.S.; Moyer, C.L.; Tebo, B.M.; Emerson, D. Hidden in Plain Sight: Discovery of Sheath-Forming, Iron-Oxidizing Zetaproteobacteria at Loihi Seamount, Hawaii, USA. FEMS Microbiol. Ecol. 2013, 85, 116–127. [Google Scholar] [CrossRef]
- Saini, G.; Chan, C.S. Near-Neutral Surface Charge and Hydrophilicity Prevent Mineral Encrustation of Fe-Oxidizing Micro-Organisms. Geobiology 2013, 11, 191–200. [Google Scholar] [CrossRef]
- Scott, J.J.; Breier, J.A.; Luther, G.W.; Emerson, D. Microbial Iron Mats at the Mid-Atlantic Ridge and Evidence That Zetaproteobacteria May Be Restricted to Iron-Oxidizing Marine Systems. PLoS ONE 2015, 10, e0119284. [Google Scholar] [CrossRef]
- Handley, K.M.; Boothman, C.; Mills, R.A.; Pancost, R.D.; Lloyd, J.R. Functional Diversity of Bacteria in a Ferruginous Hydrothermal Sediment. ISME J. 2010, 4, 1193–1205. [Google Scholar] [CrossRef]
- Frankel, R.B. Iron Biominerals: An Overview. In Iron Biominerals; Frankel, R.B., Blakemore, R.P., Eds.; Plenum: New York, NY, USA, 1990; pp. 1–6. [Google Scholar]
- Melton, E.D.; Swanner, E.D.; Behrens, S.; Schmidt, C.; Kappler, A. The Interplay of Microbially Mediated and Abiotic Reactions in the Biogeochemical Fe Cycle. Nat. Rev. Microbiol. 2014, 12, 797–809. [Google Scholar] [CrossRef]
- Mirabello, G.; Lenders, J.J.M.; Sommerdijk, N.A.J.M. Bioinspired Synthesis of Magnetite Nanoparticles. Chem. Soc. Rev. 2016, 45, 12–14. [Google Scholar] [CrossRef]
- Fuente, V.; Rufo, L.; Juárez, B.H.; Menéndez, N.; García-Hernández, M.; Salas-Colera, E.; Espinosa, A. Formation of Biomineral Iron Oxides Compounds in a Fe Hyperaccumulator Plant: Imperata cylindrica (L.) P. Beauv. J. Struct. Biol. 2016, 193, 23–32. [Google Scholar] [CrossRef]
- Faivre, D.; Godec, T.U. From Bacteria to Mollusks: The Principles Underlying the Biomineralization of Iron Oxide Materials. Angew. Chem. Int. Ed. 2015, 54, 4728–4747. [Google Scholar] [CrossRef] [PubMed]
- Holden, J.F.; Feinberg, L.F. Microbial Iron Respiration near 100 °C. Proc. SPIE 2005, 5906, 590608-4. [Google Scholar]
- Abreu, F.; Cantão, M.E.; Nicolás, M.F.; Barcellos, F.G.; Morillo, V.; Almeida, L.G.; do Nascimento, F.F.; Lefèvre, C.T.; Bazylinski, D.A.; de Vasconcelos, A.T.R.; et al. Common Ancestry of Iron Oxide- and Iron-Sulfide-Based Biomineralization in Magnetotactic Bacteria. ISME J. 2011, 5, 1634–1640. [Google Scholar] [CrossRef] [PubMed]
- Miot, J.; Remusat, L.; Duprat, E.; Gonzalez, A.; Pont, S.; Poinsot, M. Fe Biomineralization Mirrors Individual Metabolic Activity in a Nitrate-Dependent Fe(II)-Oxidizer. Front. Microbiol. 2015, 6, 879. [Google Scholar] [CrossRef]
- Kirschvink, J.L.; Hagadorn, J.W. A Grand Unified Theory of Biomineralization. In The Biomineralization of Nano and Micro Structures; Bäuerlein, E., Ed.; Wiley-VCH Verlag GmbH: Weinheim, Germany, 2000; pp. 139–150. [Google Scholar]
- Kirschvink, J.L.; Jones, D.S. Magnetite Biomineralization and Magnctoreception in Organisms: A New Biomagnetism; Plenum: New York, NY, USA, 1985. [Google Scholar]
- Seckbach, J.; Oren, A.; Stan-Lotter, H. Polyextremophiles; Springer Science & Business Media: Dordrecht, The Netherlands, 2012. [Google Scholar]
- Bazylinski, D.A.; Frankel, R.B.; Jannasch, H.W. Anaerobic Magnetite Production by a Marine, Magnetotactic Bacterium. Nature 1988, 334, 518–519. [Google Scholar] [CrossRef]
- Yan, L.; Zhang, S.; Chen, P.; Wang, W.; Wang, Y.; Li, H. Magnetic Properties of Acidithiobacillus ferrooxidans. Mater. Sci. Eng. C 2013, 33, 4026–4031. [Google Scholar] [CrossRef]
- Yan, L.; Zhang, S.; Liu, H.; Wang, W.; Chen, P.; Li, H. Optimization of Magnetosome Production by Acidithiobacillus ferrooxidans Using Desirability Function Approach. Mater. Sci. Eng. C 2016, 59, 731–739. [Google Scholar] [CrossRef]
- Lefèvre, C.T.; Frankel, R.B.; Pósfai, M.; Prozorov, T.; Bazylinski, D.A. Isolation of Obligately Alkaliphilic Magnetotactic Bacteria from Extremely Alkaline Environments. Environ. Microbiol. 2011, 13, 2342–2350. [Google Scholar] [CrossRef]
- Abreu, F.; Martins, J.L.; Souza Silveira, T.; Keim, C.N.; de Barros, H.G.P.L.; Filho, F.J.G.; Lins, U. “Candidatus Magnetoglobus multicellularis”, a Multicellular, Magnetotactic Prokaryote from a Hypersaline Environment. Int. J. Syst. Evol. Microbiol. 2007, 57, 1318–1322. [Google Scholar] [CrossRef]
- Revathy, T.; Jacob, J.; Jayasri, M.; Suthindhiran, K. Isolation and Characterization of Magnetospirillum from Saline Lagoon. World J. Microbiol. Biotechnol. 2016, 32, 109. [Google Scholar] [CrossRef]
- Wu, W.; Li, B.; Hu, J.; Li, J.; Wang, F.; Pan, Y. Iron Reduction and Magnetite Biomineralization Mediated by a Deep-Sea Iron-Reducing Bacterium Shewanella piezotolerans WP3. J. Geophys. Res. 2011, 116, G04034. [Google Scholar]
- Dong, Y.; Li, J.; Zhang, W.; Zhang, W.; Zhao, Y.; Xiao, T.; Wu, L.; Pan, H. The Detection of Magnetotactic Bacteria in Deep Sea Sediments from the East Pacific Manganese Nodule Province. Environ. Microbiol. Rep. 2016, 8, 239–249. [Google Scholar] [CrossRef]
- Stapleton, R.D.; Sabree, Z.L.; Palumbo, A.V.; Moyer, C.; Devol, A.; Roh, Y.; Zhou, J.-Z. Metal Reduction at in Situ Temperatures by Shewanella Isolates from Diverse Marine Environments. Aquat. Microb. Ecol. 2005, 38, 81–91. [Google Scholar] [CrossRef]
- Abreu, F.; Carolina, A.; Araujo, V.; Silva, K.T.; Cunha, O.D.L.; Almeida, L.G.; Geurink, C.; Farina, M.; Rodelli, D.; Jovane, L.; et al. Culture-Independent Characterization of Novel Psychrophilic Magnetotactic Cocci from Antarctic Marine Sediments. Environ. Microbiol. 2016, 18, 4426–4441. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Vali, H.; Romaner, C.S.; Phelps, T.J.; Liu, S.V. Formation of Single-Domain Magnetite by a Thermophilic Bacterium. Am. Mineral. 1998, 83, 1409–1418. [Google Scholar] [CrossRef]
- Lefèvre, C.T.; Abreu, F.; Schmidt, M.L.; Lins, U.; Frankel, R.B.; Hedlund, B.P.; Bazylinski, D.A. Moderately Thermophilic Magnetotactic Bacteria from Hot Springs in Nevada. Appl. Environ. Microbiol. 2010, 76, 3740–3743. [Google Scholar] [CrossRef]
- Bazylinski, D.A.; Lefèvre, C.T. Magnetotactic Bacteria from Extreme Environments. Life 2013, 3, 295–307. [Google Scholar] [CrossRef]
- Belkova, N.L.; Zakharova, J.R.; Tazaki, K.; Okrugin, V.M.; Parfenova, V.V. Fe-Si Biominerals in the Vilyuchinskie Hot Springs, Kamchatka Peninsula, Russia. Int. Microbiol. 2004, 7, 193–198. [Google Scholar]
- Edwards, K.J.; Glazer, B.T.; Rouxel, O.J.; Bach, W.; Emerson, D.; Davis, R.E.; Toner, B.M.; Chan, C.S.; Tebo, B.M.; Staudigel, H.; et al. Ultra-Diffuse Hydrothermal Venting Supports Fe-Oxidizing Bacteria and Massive Umber Deposition at 5000 M off Hawaii. ISME J. 2011, 5, 1748–1758. [Google Scholar] [CrossRef]
- Kashefi, K.; Lovley, D.R. Extending the Upper Temperature Limit for Life. Science 2003, 301, 934. [Google Scholar] [CrossRef]
- Wiedenheft, B.; Mosolf, J.; Willits, D.; Yeager, M.; Dryden, K.A.; Young, M.; Douglas, T. An Archaeal Antioxidant: Characterization of a Dps-like Protein from Sulfolobus solfataricus. Proc. Natl. Acad. Sci. USA 2005, 102, 10551–10556. [Google Scholar] [CrossRef]
- Ilbert, M.; Bonnefoy, V. Insight into the Evolution of the Iron Oxidation Pathways. Biochim. Biophys. Acta 2013, 1827, 161–175. [Google Scholar] [CrossRef]
- Tazaki, K. Formation of Banded Iron-Manganese Structures by Natural Microbal Communities. Clays Clay Miner. 2000, 48, 511–520. [Google Scholar] [CrossRef]
- Araujo, A.C.; Abreu, F.; Silva, K.T.; Bazylinski, D.A.; Lins, U. Magnetotactic Bacteria as Potential Sources of Bioproducts. Mar. Drugs 2015, 13, 389–430. [Google Scholar] [CrossRef] [PubMed]
- Schüler, D.; Frankel, R.B. Bacterial Magnetosomes: Microbiology, Biomineralization and Biotechnological Applications. Appl. Microb. Biotechnol. 1999, 52, 464–473. [Google Scholar] [CrossRef] [PubMed]
- Peigneux, A.; Valverde-Tercedor, C.; Lopez-Moreno, R.; Perez-Gonzalez, T.; Fernandez-Vivas, M.; Jimenez-Lopez, C. Learning from Magnetotactic Bacteria: A Review on the Synthesis of Biomimetic Nanoparticles Mediated by Magnetosome-Associated Proteins. J. Struct. Biol. 2016, 196, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Bakhshi, P.; Bain, J.; Gul, M.; Stride, E.; Edirisinghe, M.; Staniland, S. Manufacturing Man-Made Magnetosomes: High-Throughput in Situ Synthesis of Biomimetic Magnetite Loaded Nanovesicles. Macromol. Biosci. 2016, 16, 1555–1561. [Google Scholar] [CrossRef] [PubMed]
- Alphandéry, E. Applications of Magnetosomes Synthesized by Magnetotactic Bacteria in Medicine. Front. Bioeng. Biotechnol. 2014, 2, 5. [Google Scholar]
- Bain, J.; Staniland, S. Bioinspired Nanoreactors for the Biomineralisation of Metallic-Based Nanoparticles for Nanomedicine. Phys. Chem. Chem. Phys. 2015, 17, 15508–15521. [Google Scholar] [CrossRef]
- Matsunaga, T.; Okamura, Y.; Tanaka, T. Biotechnological Application of Nano-Scale Engineered Bacterial Magnetic Particles. J. Mater. Chem. 2004, 14, 2099–2105. [Google Scholar] [CrossRef]
- Mathuriya, A. Magnetotactic Bacteria: Nanodrivers of the Future. Crit. Rev. Biotechnol. 2015, 13, 389–430. [Google Scholar] [CrossRef]
- Mériaux, S.; Bouchner, M.; Marty, B.; Lalatonne, Y.; Prévéral, S.; Motte, L.; Lefèvre, C.; Geffroy, F.; Lethimonnier, F.; Péan, M.; et al. Magnetosomes, Biogenic Magnetic Nanomaterials for Brain Molecular Imaging with 17.2 T MRI Scanner. Adv. Healthc. Mater. 2015, 4, 1076–1083. [Google Scholar] [CrossRef] [PubMed]
- Jacob, J.J.; Suthindhiran, K. Magnetotactic Bacteria and Magnetosomes—Scope and Challenges. Mater. Sci. Eng. C 2016, 68, 919–928. [Google Scholar] [CrossRef]
- Picard, A.; Kappler, A.; Schmid, G.; Quaroni, L.; Obst, M. Experimental Diagenesis of Organo-Mineral Structures Formed by Microaerophilic Fe(II)-Oxidizing Bacteria. Nat. Commun. 2015, 6, 6277. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, Y.; Chen, R.; Anikeeva, P.; Jasanoff, A. Engineering Intracellular Biomineralization and Biosensing by a Magnetic Protein. Nat. Commun. 2015, 6, 8721. [Google Scholar] [CrossRef]
- Baesman, S.M.; Bullen, T.D.; Dewald, J.; Zhang, D.; Curran, S.; Islam, F.S.; Beveridge, T.J.; Oremland, R.S. Formation of Tellurium Nanocrystals during Anaerobic Growth of Bacteria That Use Te Oxyanions as Respiratory Electron Acceptors. Appl. Environ. Microbiol. 2007, 73, 2135–2143. [Google Scholar] [CrossRef]
- Munar, M.P.; Matsuo, T.; Kimura, H.; Takahashi, H.; Okamura, Y. Biomineralization of Metallic Tellurium by Bacteria Isolated From Marine Sediment Off Niigata Japan. In Biomineralization; Endo, K., Kogure, T., Nagasawa, H., Eds.; Springer: Singapore, 2018. [Google Scholar] [CrossRef]
- Espinosa-Ortiz, E.J.; Rene, E.R.; Guyot, F.; van Hullebusch, E.D.; Lens, P.N.L. Biomineralization of tellurium and selenium-tellurium nanoparticles by the white-rot fungus Phanerochaete chrysosporium. Int. Biodeterior. Biodegrad. 2017, 124, 258–266. [Google Scholar] [CrossRef]
- Walker, J.J.; Spear, J.R.; Pace, N.R. Geobiology of a Microbial Endolithic Community in the Yellowstone Geothermal Environment. Nature 2005, 434, 1011–1014. [Google Scholar] [CrossRef]
- Reitz, T.; Merroun, M.L.; Rossberg, A.; Steudtner, R.; Selenska-Pobell, S. Bioaccumulation of U(VI) by Sulfolobus acidocaldarius under Moderate Acidic Conditions. Radiochim. Acta 2011, 99, 543–553. [Google Scholar] [CrossRef]
- Dopson, M.; Ossandon, F.J.; Lövgren, L.; Holmes, D.S. Metal Resistance or Tolerance? Acidophiles Confront High Metal Loads via Both Abiotic and Biotic Mechanisms. Front. Microbiol. 2014, 5, 157. [Google Scholar] [CrossRef]
- Dopson, M.; Holmes, D.S. Metal Resistance in Acidophilic Microorganisms and Its Significance for Biotechnologies. Appl. Microbiol. Biotechnol. 2014, 98, 8133–8144. [Google Scholar] [CrossRef]
- Inskeep, W.P.; Macur, R.E.; Harrison, G.; Bostick, B.C.; Fendorf, S. Biomineralization of As(V)-Hydrous Ferric Oxyhydroxide in Microbial Mats of an Acid-Sulfate-Chloride Geothermal Spring, Yellowstone National Park. Geochim. Cosmochim. Acta 2004, 68, 3141–3155. [Google Scholar] [CrossRef]
- Mori, J.F.; Lu, S.; Händel, M.; Totsche, K.U.; Neu, T.R.; Iancu, V.V.; Tarcea, N.; Popp, J.; Küsel, K. Schwertmannite Formation at Cell Junctions by a New Filament-Forming Fe(II)-Oxidizing Isolate Affiliated with the Novel Genus Acidithrix. Microbiology 2016, 162, 62–71. [Google Scholar] [CrossRef]
- Gherman, V.D.; Boboescu, I.Z.; Pap, B.; Kondorosi, E.; Gherman, G.; Maroti, G. An Acidophilic Bacterial-Archaeal-Fungal Ecosystem Linked to Formation of Ferruginous Crusts and Stalactites. Geomicrobiol. J. 2014, 31, 407–418. [Google Scholar] [CrossRef][Green Version]
- Amils, R.; Fernández-Remolar, D.; The IPBSL Team. Río Tinto: A Geochemical and Mineralogical Terrestrial Analogue of Mars. Life 2014, 4, 511–534. [Google Scholar] [CrossRef]
- Sánchez-Román, M.; Fernández-Remolar, D.; Amils, R.; Sánchez-Navas, A.; Schmid, T.; San Martin-Uriz, P.; Rodríguez, N.; McKenzie, J.A.; Vasconcelos, C. Microbial Mediated Formation of Fe-Carbonate Minerals under Extreme Acidic Conditions. Sci. Rep. 2014, 4, 4767. [Google Scholar] [CrossRef]
- Ziegler, S.; Ackermann, S.; Majzlan, J.; Gescher, J. Matrix Composition and Community Structure Analysis of a Novel Bacterial Pyrite Leaching Community. Environ. Microbiol. 2009, 11, 2329–2338. [Google Scholar] [CrossRef] [PubMed]
- Oggerin, M.; Tornos, F.; Rodríguez, N.; del Moral, C.; Sanchez-Roman, M.; Amils, R. Specific Jarosite Biomineralization by Purpureocillium lilacinum, an Acidophilic Fungi Isolated from Río Tinto. Environ. Microbiol. 2013, 15, 2228–2237. [Google Scholar] [CrossRef]
- Oggerin, M.; Rodríguez, N.; del Moral, C.; Amils, R. Fungal Jarosite Biomineralization in Río Tinto. Res. Microbiol. 2014, 165, 719–725. [Google Scholar] [CrossRef]
- Wills, A.S.; Harrison, A.; Ritter, C.; Smith, R.I. Magnetic Properties of Pure and Diamagnetically Doped Jarosites: Model Kagomé Antiferromagnets with Variable Coverage of the Magnetic Lattice. Phys. Rev. B 2000, 61, 6156–6169. [Google Scholar] [CrossRef]
- Harrison, A. First Catch Your Hare: The Design and Synthesis of Frustrated Magnets. J. Phys. Condens. Matter 2004, 16, S553–S572. [Google Scholar] [CrossRef]
- Asokan, P.; Saxena, M.; Asolekar, S.R. Hazardous Jarosite Use in Developing Non-Hazardous Product for Engineering Application. J. Hazard. Mater. 2006, 137, 1589–1599. [Google Scholar] [CrossRef] [PubMed]
- DiGregorio, B.E. Rock Varnish as a Habitat for Extant Life on Mars. Proc. SPIE 2002, 4495, 120. [Google Scholar]
- Krinsley, D.; Dorn, R.I.; DiGregorio, B. Astrobiological Implications of Rock Varnish in Tibet. Astrobiology 2009, 9, 551–562. [Google Scholar] [CrossRef] [PubMed]
- Azua-Bustos, A.; Urrejola, C.; Vicuña, R. Life at the Dry Edge: Microorganisms of the Atacama Desert. FEBS Lett. 2012, 586, 2939–2945. [Google Scholar] [CrossRef] [PubMed]
- Dorn, R.I.; Krinsley, D.H.; Liu, T.; Anderson, S.; Clark, J.; Cahill, T.A.; Gill, T.E. Manganese-Rich Rock Varnish Does Occur in Antarctica. Chem. Geol. 1992, 99, 289–298. [Google Scholar] [CrossRef]
- Hoover, B.; Levin, G.V.; Rozanov, A.Y.; Gladstone, G.R.; Perry, R.S.; Kolb, V.M.; Lynne, B.Y.; Sephton, M.; Mcloughlin, N.; Engel, M.H.; et al. How Desert Varnish Forms? Proc. SPIE 2005, 5906, 276–287. [Google Scholar]
- Perry, R.S.; Kolb, V.M. Biological and Organic Constiuents of Desert Varnish: Review and New Hypotheses. Proc. SPIE 2004, 5163, 202–217. [Google Scholar]
- Perry, R.; Sephton, M. Solving the Mystery of Desert Varnish with Microscopy. Infocus Mag. 2008, 11, 62–76. [Google Scholar] [CrossRef]
- White, C.H. Desert Varnish. Am. J. Sci. 1924, 9, 413–420. [Google Scholar] [CrossRef]
- Dorn, R.I.; Oberlander, T.M. Microbial Origin of Desert Varnish. Science 1981, 213, 1245–1247. [Google Scholar] [CrossRef] [PubMed]
- Laudermilk, J.D. On the Origin of Desert Varnish. Am. J. Sci. 1931, 21, 51–66. [Google Scholar] [CrossRef]
- Raymond, R., Jr.; Guthrie, G.D., Jr.; Bish, D.L.; Reneau, S.L.; Chipera, S.J. Biomineralization of Manganese in Rock Varnish. Catena Suppl. 1992, 21, 321–335. [Google Scholar]
- Hungate, B.; Danin, A.; Pellerin, N.B.; Stemmler, J.; Kjellander, P.; Adams, J.B.; Staley, J.T. Characterization of Manganese-Oxidizing (MnII→MnIV) Bacteria from Negev Desert Rock Varnish: Implications in Desert Varnish Formation. Can. J. Microbiol. 1987, 33, 939–943. [Google Scholar] [CrossRef]
- Malherbe, C.; Ingley, R.; Hutchinson, I.; Edwards, H.; Carr, A.S.; Harris, L.; Boom, A. Biogeological Analysis of Desert Varnish Using Portable Raman Spectrometers. Astrobiology 2015, 15, 442–452. [Google Scholar] [CrossRef]
- Macholdt, D.S.; Jochum, K.P.; Pöhlker, C.; Arangio, A.; Förster, J.-D.; Stoll, B.; Weis, U.; Webe, B.; Müller, M.; Kappl, M.; et al. Characterization and differentiation of rock varnish types from different environments by microanalytical techniques. Chem. Geol. 2017, 459, 91–118. [Google Scholar] [CrossRef]
- Thakkar, K.N.; Mhatre, S.S.; Parikh, R.Y. Biological Synthesis of Metallic Nanoparticles. Nanomedicine 2010, 6, 257–262. [Google Scholar] [CrossRef]
- Narayanan, K.B.; Sakthivel, N. Biological Synthesis of Metal Nanoparticles by Microbes. Adv. Colloid Interface Sci. 2010, 156, 1–13. [Google Scholar] [CrossRef]
- Ghodake, G.; Eom, C.Y.; Kim, S.W.; Jin, E. Biogenic Nano-Synthesis; Towards the Efficient Production of the Biocompatible Gold Nanoparticles. Bull. Korean Chem. Soc. 2010, 31, 2771–2775. [Google Scholar] [CrossRef][Green Version]
- Iravani, S. Bacteria in Nanoparticle Synthesis: Current Status and Future Prospects. Int. Sch. Res. Not. 2014, 2014, 359316. [Google Scholar] [CrossRef] [PubMed]
- Weghuis, M.O. High Rate Biotechnology for the Metal and Mining Industry; Environmental Science & Engineering Magazine: Aurora, ON, Canada, 2013; pp. 28–31. [Google Scholar]
- Suresh, A.K.; Pelletier, D.A.; Wang, W.; Broich, M.L.; Moon, J.W.; Gu, B.; Allison, D.P.; Joy, D.C.; Phelps, T.J.; Doktycz, M.J. Biofabrication of Discrete Spherical Gold Nanoparticles Using the Metal-Reducing Bacterium Shewanella oneidensis. Acta Biomater. 2011, 7, 2148–2152. [Google Scholar] [CrossRef] [PubMed]
- Suresh, A.K.; Pelletier, D.A.; Wang, W.; Moon, J.W.; Gu, B.; Mortensen, N.P.; Allison, D.P.; Joy, D.C.; Phelps, T.J.; Doktycz, M.J. Silver Nanocrystallites: Biofabrication Using Shewanella oneidensis, and an Evaluation of Their Comparative Toxicity on Gram-Negative and Gram-Positive Bacteria. Environ. Sci. Technol. 2010, 44, 5210–5215. [Google Scholar] [CrossRef]
- Hennebel, T.; Van Nevel, S.; Verschuere, S.; De Corte, S.; De Gusseme, B.; Cuvelier, C.; Fitts, J.P.; Van Der Lelie, D.; Boon, N.; Verstraete, W. Palladium Nanoparticles Produced by Fermentatively Cultivated Bacteria as Catalyst for Diatrizoate Removal with Biogenic Hydrogen. Appl. Microbiol. Biotechnol. 2011, 91, 1435–1445. [Google Scholar] [CrossRef]
- Coker, V.S.; Telling, N.D.; Van Der Laan, G.; Pattrick, R.A.D.; Pearce, C.I.; Arenholz, E.; Tuna, F.; Winpenny, R.E.P.; Lloyd, J.R. Harnessing the Extracellular Bacterial Production of Nanoscale Cobalt Ferrite with Exploitable Magnetic Properties. ACS Nano 2009, 3, 1922–1928. [Google Scholar] [CrossRef] [PubMed]
- Kumari, D.; Qian, X.Y.; Pan, X.; Achal, V.; Li, Q.; Gadd, G.M. Microbially-Induced Carbonate Precipitation for Immobilization of Toxic Metals; Elsevier Ltd.: Amsterdam, The Netherlands, 2016; Volume 94. [Google Scholar]
- Ehrlich, H.; Simon, P.; Motylenko, M.; Wysokowski, M.; Bazhenov, V.V.; Galli, R.; Stelling, A.L.; Stawski, D.; Ilan, M.; Stöcker, H.; et al. Extreme Biomimetics: Formation of zirconium dioxide nanophase using chitinous scaffolds under hydrothermal conditions. J. Mater. Chem. B 2013, 1, 5092–5099. [Google Scholar] [CrossRef]
- Wysokowski, M.; Petrenko, I.; Stelling, A.L.; Stawski, D.; Jesionowski, T.; Ehrlich, H. Chitin as a versatile template for extreme biomimetics. Polymers 2015, 7, 235–265. [Google Scholar] [CrossRef]
- Szatkowski, T.; Wysokowski, M.; Lota, G.; Peziak, D.; Bazhenov, V.V.; Nowaczyk, G.; Walter, J.; Molodtsov, S.L.; Stöcker, H.; Himcinschi, C.; et al. Novel nanostructured hematite–spongin composite developed using extreme biomimetic approach. RSC Adv. 2015, 5, 79031–79040. [Google Scholar] [CrossRef]
- Szatkowski, T.; Siwińska–Stefańska, K.; Wysokowski, M.; Stelling, A.L.; Joseph, Y.; Ehrlich, H.; Jesionowski, T. Immobilization of titanium (IV) oxide onto 3D spongin scaffolds of marine sponge origin according to extreme biomimetics principles for removal of C.I. Basic Blue 9. Biomimetics 2017, 2, 4. [Google Scholar] [CrossRef]
- Szatkowski, T.; Kopczyński, K.; Motylenko, M.; Borrmann, H.; Mania, B.; Graś, M.; Lota, G.; Bazhenov, V.V.; Rafaja, D.; Roth, F.; et al. Extreme biomimetics: Carbonized 3D spongin scaffold as a novel support for nanostructured manganese oxide (IV) and its electrochemical applications. Nano Res. 2018, 11, 4199–4214. [Google Scholar] [CrossRef]
- Petrenko, I.; Summers, A.P.; Simon, P.; Żółtowska-Aksamitowska, S.; Motylenko, M.; Schimpf, C.; Rafaja, D.; Roth, F.; Kummer, K.; Brendler, E.; et al. Extreme Biomimetics: Preservation of molecular detail in centimetre scale samples of biological meshes laid down by sponges. Sci. Adv. 2019, 5, eaax2805. [Google Scholar] [CrossRef]
- Rothschild, L.J.; Mancinelli, R.L. Life in Extreme Environments. Nature 2001, 409, 1092–1101. [Google Scholar] [CrossRef]
- Ehrlich, H. (Ed.) Extreme Biomimetics; Springer International Publishing: Cham, Switzerland, 2019; p. 276. [Google Scholar]
- Demarchi, B.; Hall, S.; Roncal-Herrero, T.; Freeman, C.L.; Woolley, J.; Crisp, M.K.; Wilson, J.; Fotakis, A.; Fischer, R.; Kessler, B.M.; et al. Protein Sequences Bound to Mineral Surfaces Persist into Deep Time. Elife 2016, 5, e17092. [Google Scholar] [CrossRef]
- Govarthanan, M.; Lee, K.J.; Cho, M.; Kim, J.S.; Kamala-Kannan, S.; Oh, B.T. Significance of Autochthonous bacillus sp. KK1 on Biomineralization of Lead in Mine Tailings. Chemosphere 2013, 90, 2267–2272. [Google Scholar] [CrossRef] [PubMed]
- Acharya, C.; Chandwadkar, P.; Nayak, C. Unusual Versatility of the Filamentous, Diazotrophic Cyanobacterium Anabaena Torulosa Revealed for Its Survival during Prolonged Uranium Exposure. Appl. Environ. Microbiol. 2017, 83, e03356-16. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, Y.; Kopp, R.; Kogure, T.; Suga, A.; Takai, K.; Tsuchida, S.; Ozaki, N.; Endo, K.; Hashimoto, J.; Kato, Y. Sclerite Formation in the Hydrothermal-Vent “Scaly-Foot” Gastropod—possible Control of Iron Sulfide Biomineralization by the Animal. Earth Planet. Sci. Lett. 2006, 242, 39–50. [Google Scholar] [CrossRef]
- González-Muñoz, M.T.; De Linares, C.; Martínez-Ruiz, F.; Morcillo, F.; Martin-Ramos, D.; Arias, J.M. Ca-Mg Kutnahorite and Struvite Production by Idiomarina Strains at Modern Seawater Salinities. Chemosphere 2008, 72, 465–472. [Google Scholar] [CrossRef] [PubMed]
- Rhee, Y.J.; Hillier, S.; Gadd, G.M. A New Lead Hydroxycarbonate Produced During Transformation of Lead Metal by the Soil Fungus Paecilomyces javanicus. Geomicrobiol. J. 2016, 33, 250–260. [Google Scholar] [CrossRef]
- Li, Q.; Csetenyi, L.; Gadd, G.M. Biomineralization of Metal Carbonates by Neurospora crassa. Environ. Sci. Technol. 2014, 48, 14409–14416. [Google Scholar] [CrossRef]
- Liang, X.; Kierans, M.; Ceci, A.; Hillier, S.; Gadd, G.M. Phosphatase-Mediated Bioprecipitation of Lead by Soil Fungi. Environ. Microbiol. 2016, 18, 219–231. [Google Scholar] [CrossRef]
- Yao, S.; Jin, B.; Liu, Z.; Shao, C.; Zhao, R.; Wang, X.; Tang, R. Biomineralization: From Material Tactics to Biological Strategy. Adv. Mater. 2017, 29, 1605903. [Google Scholar] [CrossRef] [PubMed]
- Knoll, A.H. Biomineralization and Evolutionary History. Rev. Mineral. Geochem. 2003, 54, 329–356. [Google Scholar] [CrossRef]
- Wood, R. Exploring the drivers of early biomineralization. Emergy Top. Life Sci. 2018, 2, 201–212. [Google Scholar]
- Evans, J.S. The Biomineralization Proteome: Protein Complexity for a Complex Bioceramic Assembly Process. Proteomics 2019, 19, e1900036. [Google Scholar] [CrossRef] [PubMed]
- Oggerin, M.; Tornos, F.; Rodriguez, N.; Pascual, L.; Amils, R. Fungal Iron Biomineralization in Río Tinto. Minerals 2016, 6, 37. [Google Scholar] [CrossRef]
- Choi, Y.; Lee, S.Y. Biosynthesis of inorganic nanomaterials using microbial cells and bacteriophages. Nat. Rev. Chem. 2020, 4, 638–656. [Google Scholar] [CrossRef]
- Benzerara, K.; Miot, J.; Morin, G.; Ona-Nguema, G.; Skouri-Panet, F.; Ferard, C. Significance, mechanisms and environmental implications of microbial biomineralization. Importance, mécanismes et implications environnementales de la biominéralisation par les microorganisms. Comptes Rendus Geosci. 2011, 343, 160–167. [Google Scholar] [CrossRef]
- Brasier, M. Why Do Lower Plants and Animals Biomineralize? Paleobiology 1986, 12, 241–250. [Google Scholar] [CrossRef]
- Domingues, V.S.; de Souza Monteiro, A.; Júlio, A.D.L.; Queiroz, A.L.L.; dos Santos, V.L. Diversity of Metal-Resistant and Tensoactive-Producing Culturable Heterotrophic Bacteria Isolated from a Copper Mine in Brazilian Amazonia. Sci. Rep. 2020, 10, 617. [Google Scholar] [CrossRef]
- Chen, J.; Li, J.; Zhang, H.; Shi, W.; Liu, Y. Bacterial Heavy-Metal and Antibiotic Resistance Genes in a Copper Tailing Dam Area in Northern China. Front. Microbiol. 2019, 10, 1916. [Google Scholar] [CrossRef]
- De Lima e Silva, A.; Carvalho, M.A.; Souza, S.; Dias, P.; Silva, F.R.; Saramago, C.; Bento, C.; Hofer, E. Heavy metal tolerance (Cr, Ag AND Hg) in bacteria isolated from sewage. Braz. J. Microbiol. 2012, 43, 1620–1631. [Google Scholar] [CrossRef]
- Trevors, J.T.; Oddie, K.M.; Belliveau, B.H. Metal resistance in bacteria. FEMS Microbiol. Rev. 1985, 32, 39–54. [Google Scholar] [CrossRef]
- Davis, I.J. Characterization of Heavy Metal and Antibiotic-Resistance Genes in Oral Bacteria. Ph.D. Thesis, University College London, London, UK, 2003. Available online: https://discovery.ucl.ac.uk/id/eprint/10104640 (accessed on 1 March 2021).
- Jackson, D.J.; Macis, L.; Reitner, J.; Wörheide, G. A horizontal gene transfer supported the evolution of an early metazoan biomineralization strategy. BMC Evol. Biol. 2011, 11, 238. [Google Scholar] [CrossRef] [PubMed]
- Marvasi, M.; Casillas-Santiago, L.M.; Henríquez, T.; Casillas-Martinez, L. Involvement of etfA gene during CaCO3 pre-cipitation in Bacillus subtilis biofilm. Geomicrobiol J. 2017, 34, 722–728. [Google Scholar] [CrossRef]
- Perito, B.; Biagiotti, L.; Daly, S.; Galizzi, A.; Tiano, P.; Mastromei, G. Bacterial Genes Involved in Calcite Crystal Precipitation. In Of Microbes and Art; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 2000; pp. 219–230. [Google Scholar]
- Zhu, Y.; Ma, N.; Jin, W.; Wu, S.; Sun, C. Genomic and Transcriptomic Insights into Calcium Carbonate Biomineralization by Marine Actinobacterium brevibacterium linens BS258. Front. Microbiol. 2017, 8, 602. [Google Scholar] [CrossRef] [PubMed]
- Zaquin, T.; Malik, A.; Drake, J.L.; Putnam, H.M.; Mass, T. Evolution of Protein-Mediated Biomineralization in Scleractinian Corals. Front. Genet. 2021, 12, 618517. [Google Scholar] [CrossRef]
- Livingston, B.T.; Killian, C.E.; Wilt, F.; Cameron, A.; Landrum, M.J.; Ermolaeva, O.; Sapojnikov, V.; Maglott, D.R.; Bu-chanan, A.M.; Ettensohn, C.A. A genome-wide analysis of biomineralization-related proteins in the sea urchin Strongylo-centrotus purpuratus. Dev. Biol. 2006, 300, 335–348. [Google Scholar] [CrossRef]
- Cameron, C.B.; Bishop, C.D. Biomineral ultrastructure, elemental constitution and genomic analysis of biomineralization-related proteins in hemichordates. Proc. R. Soc. B Boil. Sci. 2012, 279, 3041–3048. [Google Scholar] [CrossRef]
- Sleight, V.A.; Antczak, P.; Falciani, F.; Clark, M.S. Computationally predicted gene regulatory networks in molluscan bi-omineralization identify extracellular matrix production and ion transportation pathways. Bioinformatics 2020, 36, 1326–1332. [Google Scholar]
- Hildebrand, M. Diatoms, Biomineralization Processes, and Genomics. Chem. Rev. 2008, 108, 4855–4874. [Google Scholar] [CrossRef]
- Shimizu, K.; Kobayashi, H.; Nishi, M.; Tsukahara, M.; Bito, T.; Arima, J. Exploration of Genes Associated with Sponge Silicon Biomineralization in the Whole Genome Sequence of the Hexactinellid Euplectella curvistellata. In Biomineralization; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 2018; pp. 147–153. [Google Scholar]
- Moriceau, B.; Gehlen, M.; Tréguer, P.; Baines, S.; Livage, J.; André, L. Editorial: Biogeochemistry and Genomics of Silicification and Silicifiers. Front. Mar. Sci. 2019, 6, 57. [Google Scholar] [CrossRef]
- Pohl, A.; Berger, F.; Sullan, R.M.A.; Valverde-Tercedor, C.; Freindl, K.; Spiridis, N.; Lefevre, C.; Menguy, N.; Klumpp, S.; Blank, K.G.; et al. Decoding Biomineralization: Interaction of a Mad10-Derived Peptide with Magnetite Thin Films. Nano Lett. 2019, 19, 8207–8215. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Liu, X.; Xiao, Y.; Hao, H.; Zhang, Y.; Tang, R. Biomineralization State of Viruses and Their Biological Potential. Chemistry 2018, 24, 11518–11529. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Dash, H.R.; Chakraborty, J. Genetic basis and importance of metal resistant genes in bacteria for bioremediation of contaminated environments with toxic metal pollutants. Appl. Microbiol. Biotechnol. 2016, 100, 2967–2984. [Google Scholar] [CrossRef] [PubMed]
- Young, T.R.; Xiao, Z. Principles and practice of determining metal–protein affinities. Biochem. J. 2021, 478, 1085–1116. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Hu, X.; Zhang, X.; Gao, S.; Ding, C.; Feng, Y.; Bao, W. Identification of metal ion binding sites based on amino acid sequences. PLoS ONE 2017, 12, e0183756. [Google Scholar] [CrossRef] [PubMed]
- Yannone, S.M.; Hartung, S.; Menon, A.L.; Adams, M.W.; A Tainer, J. Metals in biology: Defining metalloproteomes. Curr. Opin. Biotechnol. 2012, 23, 89–95. [Google Scholar] [CrossRef]
- Maret, W. The Metals in the Biological Periodic System of the Elements: Concepts and Conjectures. Int. J. Mol. Sci. 2016, 17, 66. [Google Scholar] [CrossRef]
- Maret, W. Metallomics: The Science of Biometals and Biometalloids. Adv. Exp. Med. Biol. 2018, 1055, 1–20. [Google Scholar] [CrossRef]
- Kosman, D.J. A holistic view of mammalian (vertebrate) cellular iron uptake. Metallomics 2020, 12, 1323–1334. [Google Scholar] [CrossRef]




| Biomineral | Chemical Formula | Organism | Metal/Metalloid | Ref. |
|---|---|---|---|---|
| Alamosite | PbSiO3 | Bacillus sp. KK1 | Pb | [249] |
| Bioscorodite | FeAsO4·2H2O | Acidianus suljidivorans, Sulfolobales spp. (70 °C, pH 1.2) | FeAs | [118,250,251] |
| Chernikovite | H2(UO2)2(PO4)2·8H2O | Anabaena torulosa (cyanobacteria) | U | [118] |
| Eskaloite | Cr2O3 | Schwanniomyces occidentalis (50 °C) | Cr(VI) | [252] |
| Greigite | Fe3S4 | Chrysomallon squamiferum (thermophilic gastropod) | Fe | [253] |
| Hydro- cerussite | (Pb3(CO3)2(OH)2) | Paecilomyces javanicus | Pb | [254] |
| Kutnahorite | (Ca(Mn2+,Mg,Fe2+)(CO3)2) | Idiomarina sp. (Halophilic) | Ca, Mn, Mg, Fe | [255] |
| Ni-struvite | Ni(NH4)(PO4)∙6H2O | Streptomyces acidiscabies | Ni | [256] |
| Orpiment | As2S3 | Shewanella sp. Desulfosporosinus auripigmenti (anaerobic) | As | [257,258] |
| Otavite | CdCO3 | Neurospora crassa (fungus) | Cd | [259] |
| Plumbonacrite | (Pb10(CO3)6O(OH)6) | Paecilomyces javanicus | Pb | [259] |
| Pyrite | FeS2 | Chrysomallon squamiferum (thermophilic gastropod) | Fe | [260] |
| Pyromorphite | Pb5(PO4)3Cl | Paecilomyces javanicus | Pb | [261] |
| Realgar | AsS | As-reducing bacterium closely related to Caloramator and Thermobrachium. (Anaerobic, moderately thermophilic) | As | [262] |
| Sphalerite/Wurtzite | (Zn0.88Fe0.12)S | Alvinella pompejana (thermophilic worm) | Zn, Fe | [78] |
| Tellurium-based biominerals | Te(0) | Sulfurospirillum barnesii (anaerobic) | Te | [211] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ehrlich, H.; Bailey, E.; Wysokowski, M.; Jesionowski, T. Forced Biomineralization: A Review. Biomimetics 2021, 6, 46. https://doi.org/10.3390/biomimetics6030046
Ehrlich H, Bailey E, Wysokowski M, Jesionowski T. Forced Biomineralization: A Review. Biomimetics. 2021; 6(3):46. https://doi.org/10.3390/biomimetics6030046
Chicago/Turabian StyleEhrlich, Hermann, Elizabeth Bailey, Marcin Wysokowski, and Teofil Jesionowski. 2021. "Forced Biomineralization: A Review" Biomimetics 6, no. 3: 46. https://doi.org/10.3390/biomimetics6030046
APA StyleEhrlich, H., Bailey, E., Wysokowski, M., & Jesionowski, T. (2021). Forced Biomineralization: A Review. Biomimetics, 6(3), 46. https://doi.org/10.3390/biomimetics6030046