
Characterization of Gramicidin A in Triblock and Diblock Polymersomes and Hybrid Vesicles via Continuous Wave Electron Paramagnetic Resonance Spectroscopy



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Polymersome Preparation
2.2. Hybrid Vesicle Preparation
2.3. Vesicle Preparation
2.4. Spin-Labeled Lipids
2.5. Transmission Electron Microscopy
2.6. Gramicidin A Preparation
2.7. CW-EPR Measurements
2.8. EPR Spectral Simulations
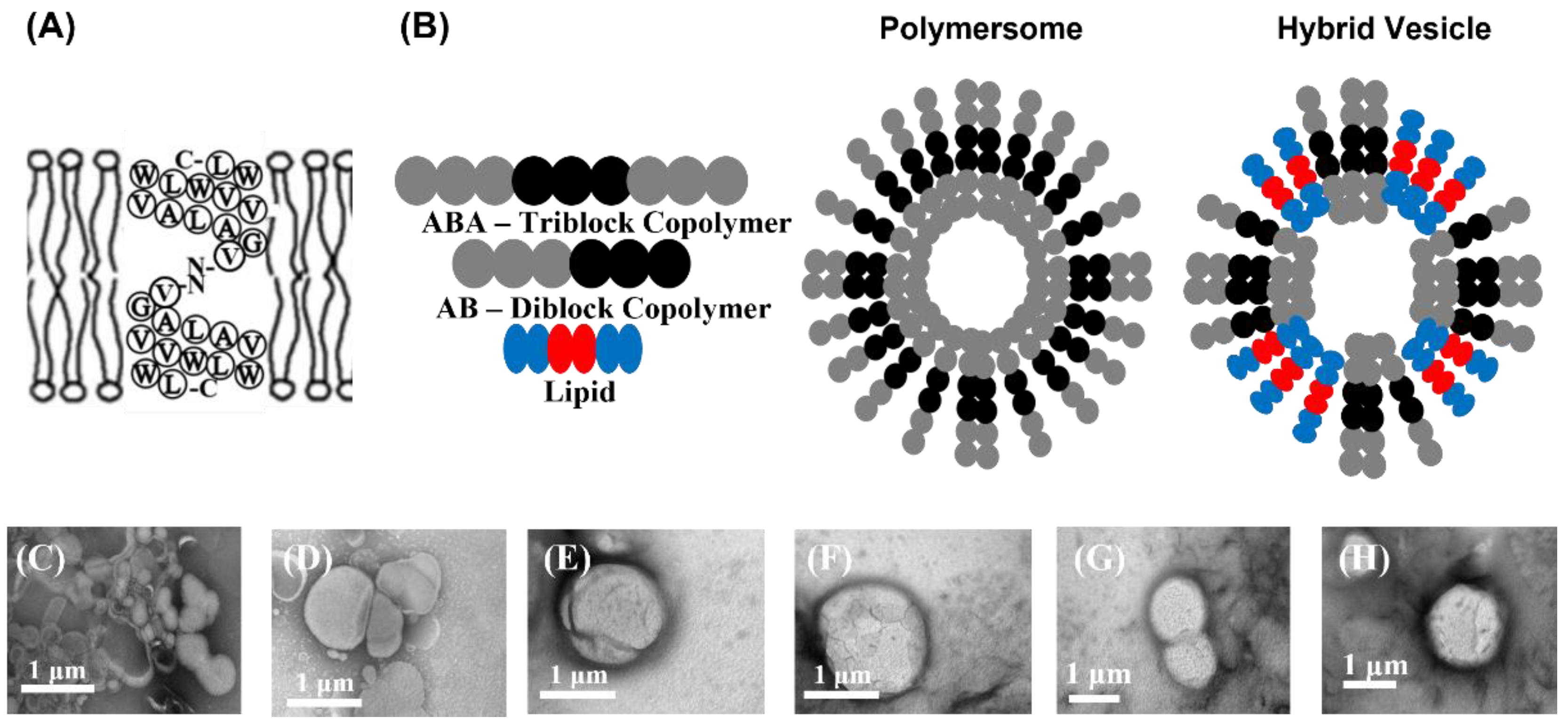
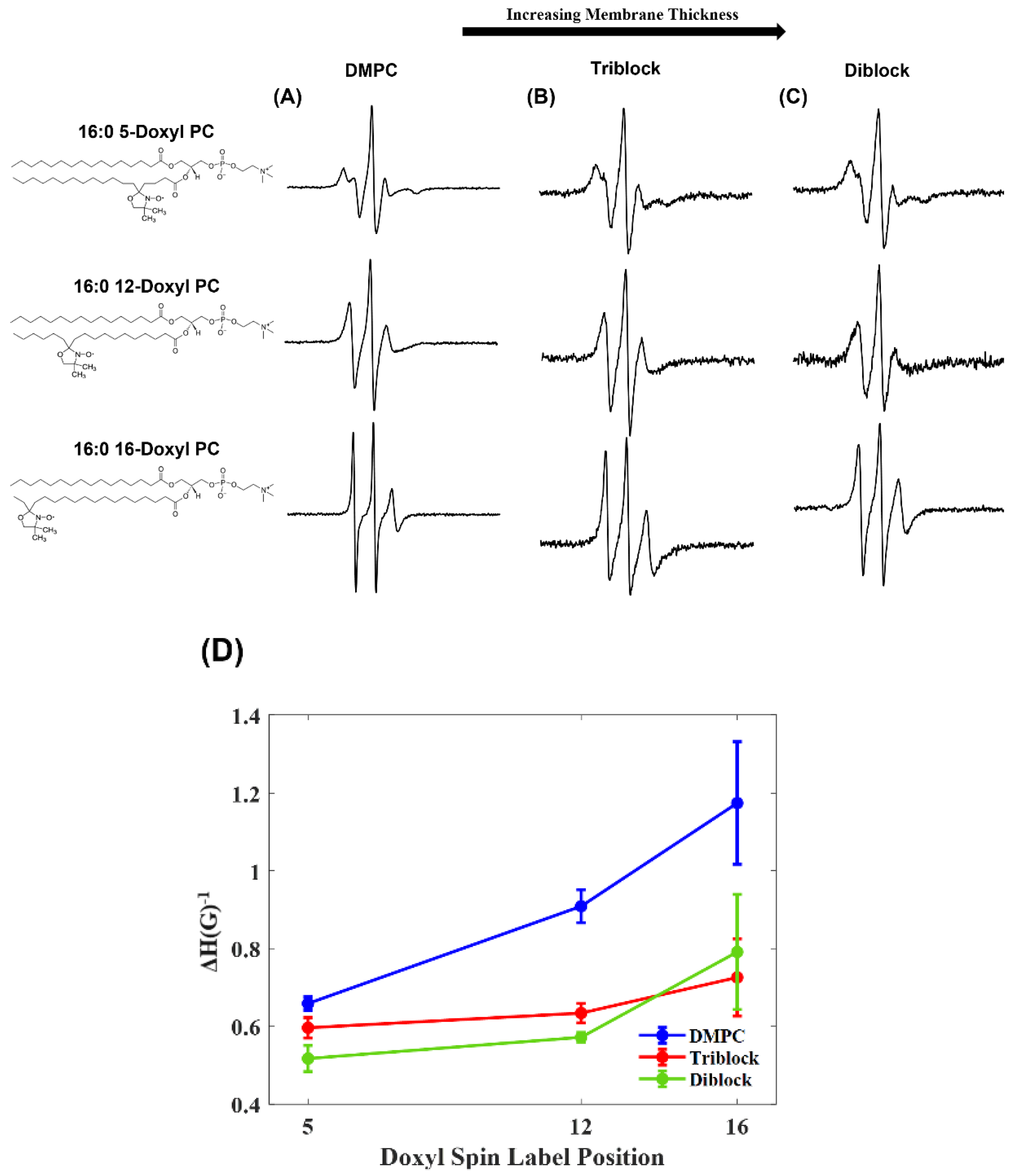
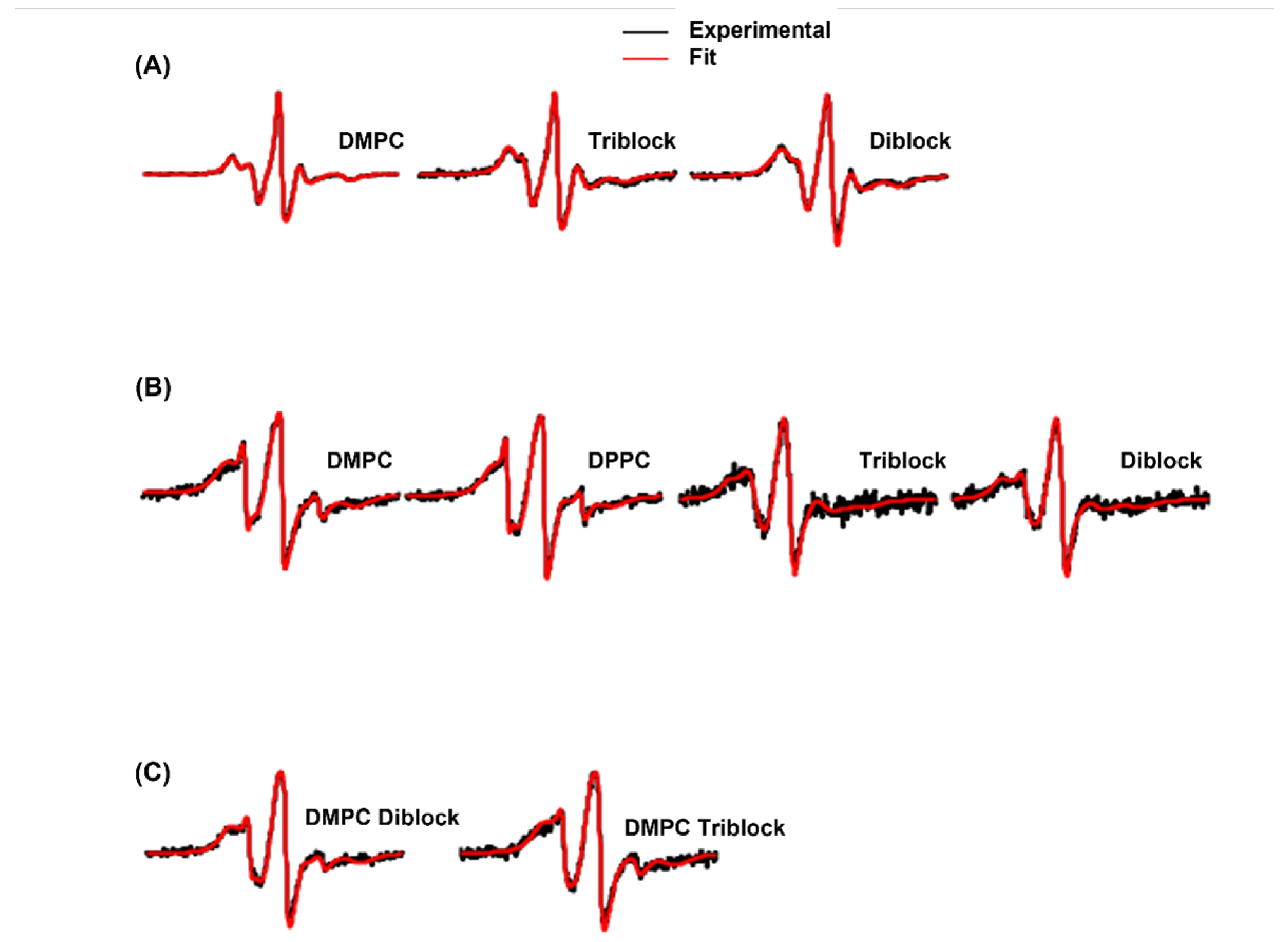
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Majeed, S.; Ahmad, A.B.; Sehar, U.; Georgieva, E.R. Lipid Membrane Mimetics in Functional and Structural Studies of Integral Membrane Proteins. Membranes 2021, 11, 685. [Google Scholar] [CrossRef]
- Yin, H.; Flynn, A.D. Drugging Membrane Protein Interactions. Annu. Rev. Biomed. Eng. 2016, 18, 51–76. [Google Scholar] [CrossRef]
- Beales, P.; Khan, S.; Muench, S.; Jeuken, L. Durable vesicles for reconstitution of membrane proteins in biotechnology. Biochem. Soc. Trans. 2017, 45, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Raschle, T.; Hiller, S.; Etzkorn, M.; Wagner, G. Nonmicellar systems for solution NMR spectroscopy of membrane proteins. Curr. Opin. Struct. Biol. 2010, 20, 471–479. [Google Scholar] [CrossRef] [PubMed]
- Vold, R.R.; Prosser, R.S. Magnetically oriented phospholipid bilayered micelles for structural studies of polypeptides. Does the ideal bicelle exist? J. Magn. Reson. Ser. B 1996, 113, 267–271. [Google Scholar] [CrossRef]
- Duerr, U.H.N.; Gildenberg, M.; Ramamoorthy, A. The Magic of Bicelles Lights Up Membrane Protein Structure. Chem. Rev. 2012, 112, 6054–6074. [Google Scholar] [CrossRef] [PubMed]
- González-Pérez, A.; Stibius, K.; Vissing, T.; Nielsen, C.; Mouritsen, O. Biomimetic Triblock Copolymer Membrane Arrays: A Stable Template for Functional Membrane Proteins. Langmuir 2009, 25, 10447–10450. [Google Scholar] [CrossRef]
- Meyer, R.; Hussmann, G.; Peterson, N.; Santos, J.; Tuesca, A. A scalable and robust cationic lipid/polymer hybrid nanoparticle platform for mRNA delivery. Int. J. Pharm. 2022, 611, 121314. [Google Scholar] [CrossRef]
- Zhang, X.; Tanner, P.; Graff, A.; Palivan, C.; Meier, W. Mimicking the cell membrane with block copolymer membranes. J. Polym. Sci. Part A-Polym. Chem. 2012, 50, 2293–2318. [Google Scholar] [CrossRef]
- Zheng, C.; Han, L.; Yap, C.W.; Xie, B.; Chen, Y. Progress and problems in the exploration of therapeutic targets. Drug Discov. Today 2006, 11, 412–420. [Google Scholar] [CrossRef]
- Tonge, S.; Tighe, B. Responsive hydrophobically associating polymers: A review of structure and properties. Adv. Drug Deliv. Rev. 2001, 53, 109–122. [Google Scholar] [CrossRef] [PubMed]
- Rideau, E.; Dimova, R.; Schwille, P.; Wurm, F.R.; Landfester, K. Liposomes and polymersomes: A comparative review towards cell mimicking. Chem. Soc. Rev. 2018, 47, 8572–8610. [Google Scholar] [CrossRef]
- Lo, C.; Zeng, J. Application of polymersomes in membrane protein study and drug discovery: Progress, strategies, and perspectives. Bioeng. Transl. Med. 2023, 8, e10350. [Google Scholar] [CrossRef] [PubMed]
- Martin, M.; Dubbs, T.; Fried, J. Planar Bilayer Measurements of Alamethicin and Gramicidin Reconstituted in Biomimetic Block Copolymers. Langmuir 2017, 33, 1171–1179. [Google Scholar] [CrossRef]
- Dao, T.; Brûlet, A.; Fernandes, F.; Er-Rafik, M.; Ferji, K.; Schweins, R.; Chapel, J.; Schmutz, F.; Prieto, M.; Sandre, O.; et al. Mixing Block Copolymers with Phospholipids at the Nanoscale: From Hybrid Polymer/Lipid Wormlike Micelles to Vesicles Presenting Lipid Nanodomains. Langmuir 2017, 33, 1705–1715. [Google Scholar] [CrossRef]
- Le Meins, J.; Schatz, C.; Lecommandoux, S.; Sandre, O. Hybrid polymer/lipid vesicles: State of the art and future perspectives. Mater. Today 2013, 16, 397–402. [Google Scholar] [CrossRef]
- Meyer, C.E.; Abram, S.L.; Craciun, I.; Palivan, C.G. Biomolecule-polymer hybrid compartments: Combining the best of both worlds. Phys. Chem. Chem. Phys. 2020, 22, 11197–11218. [Google Scholar] [CrossRef] [PubMed]
- Paxton, W.; McAninch, P.; Achyuthan, K.; Shin, S.; Monteith, H. Monitoring and modulating ion traffic in hybrid lipid/polymer vesicles. Colloids Surf. B-Biointerfaces 2017, 159, 268–276. [Google Scholar] [CrossRef]
- Schulz, M.; Binder, W. Mixed Hybrid Lipid/Polymer Vesicles as a Novel Membrane Platform. Macromol. Rapid Commun. 2015, 36, 2031–2041. [Google Scholar] [CrossRef]
- Kelkar, D.; Chattopadhyay, A. The gramicidin ion channel: A model membrane protein. Biochim. Et Biophys. Acta-Biomembr. 2007, 1768, 2011–2025. [Google Scholar] [CrossRef]
- Qin, Z.; Tepper, H.; Voth, G. Effect of membrane environment on proton permeation through gramicidin a channels. J. Phys. Chem. B 2007, 111, 9931–9939. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Fried, J. A hierarchical approach for predicting the transport properties of the gramicidin A channel. Soft Matter 2007, 3, 1041–1052. [Google Scholar] [CrossRef] [PubMed]
- Ahammad, T.; Drew, D.L., Jr.; Sahu, I.D.; Serafin, R.A.; Clowes, K.R.; Lorigan, G.A. Continuous Wave Electron Paramagnetic Resonance Spectroscopy Reveals the Structural Topology and Dynamic Properties of Active Pinholin S2168 in a Lipid Bilayer. J. Phys. Chem. B 2019, 123, 8048–8056. [Google Scholar] [CrossRef]
- Sahu, I.D.; Lorigan, G.A. Site-Directed Spin Labeling EPR for Studying Membrane Proteins. Biomed Res. Int. 2018, 2018, 3248289. [Google Scholar] [CrossRef]
- Sahu, I.D.; Lorigan, G.A. Electron paramagnetic resonance as a tool for studying membrane proteins. Biomolecules 2020, 10, 763. [Google Scholar] [CrossRef]
- Dzikovski, B.G.; Borbat, P.P.; Freed, J.H. Spin-labeled gramicidin a: Channel formation and dissociation. Biophys. J. 2004, 87, 3504–3517. [Google Scholar] [CrossRef] [PubMed]
- GE, M.; Freed, J. An electron-spin-resonance study of interactions between gramicidin-A’ and phosphatidylcholine bilayers. Biophys. J. 1993, 65, 2106–2123. [Google Scholar] [CrossRef]
- Patyal, B.; Crepeau, R.; Freed, J. Lipid-gramicidin interactions using two-dimensional Fourier-transform electron spin resonance. Biophys. J. 1997, 73, 2201–2220. [Google Scholar] [CrossRef]
- Bouchard, M.; Auger, M. Solvent history dependence of gramicidin-lipid interactions—A raman and infrared spectroscopic STUDY. Biophys. J. 1993, 65, 2484–2492. [Google Scholar] [CrossRef]
- Strauba, J.; Nowotarskib, M.; Luc, J.; Shethd, T.; Jiaod, S.; Fishera, M.; Shelld, M.; Helgesond, M.; Jerschowc, A.; Han, S. Phosphates form spectroscopically dark state assemblies in common aqueous solutions. Proc. Natl. Acad. Sci. USA 2023, 120, e2206765120. [Google Scholar] [CrossRef]
- Lu, J.; Straub, J.; Nowotarski, M.; Han, S.; Xu, X.; Jerschow, A. Spectroscopically dark phosphate features revealed by chemical exchange saturation transfer. NMR Biomed. 2024, 37, e5057. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Peyear, T.; Bennett, W.; Andersen, O.; Lightstone, F.; Ingólfsson, H. Molecular Mechanism for Gramicidin Dimerization and Dissociation in Bilayers of Different Thickness. Biophys. J. 2019, 117, 1831–1844. [Google Scholar] [CrossRef]
- Itel, F.; Chami, M.; Najer, A.; Lörcher, S.; Wu, D.; Dinu, I.; Meier, W. Molecular Organization and Dynamics in Polymersome Membranes: A Lateral Diffusion Study. Macromolecules 2014, 47, 7588–7596. [Google Scholar] [CrossRef]
- Lomora, M.; Garni, M.; Itel, F.; Tanner, P.; Spulber, M.; Palivan, C. Polymersomes with engineered ion selective permeability as stimuli-responsive nanocompartments with preserved architecture. Biomaterials 2015, 53, 406–414. [Google Scholar] [CrossRef] [PubMed]
- Steinkühler, J.; Jacobs, M.; Boyd, M.; Villaseñor, C.; Loverde, S.; Kamat, N. PEO-b-PBD Diblock Copolymers Induce Packing Defects in Lipid/Hybrid Membranes and Improve Insertion Rates of Natively Folded Peptides. Biomacromolecules 2022, 23, 4756–4765. [Google Scholar] [CrossRef] [PubMed]
- Itel, F.; Najer, A.; Palivan, C.; Meier, W. Dynamics of Membrane Proteins within Synthetic Polymer Membranes with Large Hydrophobic Mismatch. Nano Lett. 2015, 15, 3871–3878. [Google Scholar] [CrossRef]
- Bali, A.P.; Sahu, I.D.; Craig, A.F.; Clark, E.E.; Burridge, K.M.; Dolan, M.T.; Dabney-Smith, C.; Konkolewicz, D.; Lorigan, G.A. Structural characterization of styrene-maleic acid copolymer-lipid nanoparticles (SMALPs) using EPR spectroscopy. Chem. Phys. Lipids 2019, 220, 6–13. [Google Scholar] [CrossRef]
- Rizzolo, F.; Sabatino, G.; Chelli, M.; Rovero, P.; Papini, A. A convenient microwave-enhanced solid-phase synthesis of difficult peptide sequences: Case study of Gramicidin A and CSF114(Glc). Int. J. Pept. Res. Ther. 2007, 13, 203–208. [Google Scholar] [CrossRef]
- Schneider, D.J.; Freed, J.H. Calculating slow motional magnetic resonance spectra: A user’s guide. In Biological Magnetic Resonance; Berlinger, L.J., Ed.; Plenum Publishing: New York, NY, USA, 1989. [Google Scholar]
- Budil, D.E.; Lee, S.; Saxena, S.; Freed, J.H. Nonlinear-Least-Squares Analysis of Slow-Motion EPR Spectra in One and Two Dimensions Using a Modified Levenberg–Marquardt Algorithm. J. Magn. Reson. Ser. A 1996, 120, 155–189. [Google Scholar] [CrossRef]
- Camargos, H.S.; Alonso, A. Electron paramagnetic resonance (epr) spectral components of spin-labeled lipids in saturated phospholipid bilayers. effect of cholesterol. Quim. Nova 2013, 36, 815–U152. [Google Scholar] [CrossRef]
- Sahu, I.D.; Zhang, R.; Dunagan, M.M.; Craig, A.F.; Lorigan, G.A. Characterization of KCNE1 inside lipodisq nanoparticles for EPR spectroscopic studies of membrane proteins. J Phys. Chem. B 2017, 121, 5312–5321. [Google Scholar] [CrossRef] [PubMed]
- Weil, J.A.; Bolton, J.R. Electron Paramagnetic Resonance: Elementary Theory and Practical Applications; Wiley-Interscience; John Wiley & Sons: Hoboken, NJ, USA, 2007. [Google Scholar]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gordon, E.A.; Sahu, I.D.; Fried, J.R.; Lorigan, G.A. Characterization of Gramicidin A in Triblock and Diblock Polymersomes and Hybrid Vesicles via Continuous Wave Electron Paramagnetic Resonance Spectroscopy. Biomimetics 2025, 10, 154. https://doi.org/10.3390/biomimetics10030154
Gordon EA, Sahu ID, Fried JR, Lorigan GA. Characterization of Gramicidin A in Triblock and Diblock Polymersomes and Hybrid Vesicles via Continuous Wave Electron Paramagnetic Resonance Spectroscopy. Biomimetics. 2025; 10(3):154. https://doi.org/10.3390/biomimetics10030154
Chicago/Turabian StyleGordon, Emma A., Indra D. Sahu, Joel R. Fried, and Gary A. Lorigan. 2025. "Characterization of Gramicidin A in Triblock and Diblock Polymersomes and Hybrid Vesicles via Continuous Wave Electron Paramagnetic Resonance Spectroscopy" Biomimetics 10, no. 3: 154. https://doi.org/10.3390/biomimetics10030154
APA StyleGordon, E. A., Sahu, I. D., Fried, J. R., & Lorigan, G. A. (2025). Characterization of Gramicidin A in Triblock and Diblock Polymersomes and Hybrid Vesicles via Continuous Wave Electron Paramagnetic Resonance Spectroscopy. Biomimetics, 10(3), 154. https://doi.org/10.3390/biomimetics10030154