
AYA22A Aptamers Mitigate Peanut Allergenicity: Insights from Degranulation Assays and Modulating Immune Responses

 ,
,  , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Initial Aptamer Generation
2.2. Iterative Selection and Mutation
2.3. Secondary and Tertiary Structure Prediction
2.4. Docking Simulation
2.5. Cell Lines
2.6. Characterization of Aptamer Binding to Ara h2 Peanut Protein by ELISA
2.7. Assessment of the Specificity of AYA22A Aptamers to Ara h2 Protein by Competition ELISA
2.8. Aptamer Specificity to Ara Proteins (Ara h1, Ara h2, and Ara h6) Determined by ELISA
2.9. Extraction of Crude Peanut Protein
2.10. Extraction of Crude Protein from Peanut Butter
2.11. RBL-2H3 Cells Degranulation and -Hexosaminidase Assay
2.12. Detection of Aptamer Uptake by Peanut Plant by Real-Time PCR Assay
2.13. Enzyme-Linked Immunosorbent Assay (ELISA)-Based Based Detection of AYA22AR321 in Peanut Crude Samples
2.14. Aptamer Detection in AYA22AR321-Watered Peanut Plants Using ELISA
2.15. PBMC Isolation and Generation of Human Monocyte-Derived Dendritic Cells
2.16. AYA22A Aptamer Attenuates Cross-Presentation of Peanut Allergens by Mo-DCs
2.17. In Vitro Co-Culture Model of Mo-DCs and T Cells to Evaluate the Effect of AYA22A Aptamers on Th2 Cytokines Triggered by Peanut Allergens
2.18. Mechanistic Evaluation of AYA22A Aptamers in Inducing T-Cell Cytokines: Co-Culture Study with Autologous PBMCs
2.19. Mechanistic Evaluation of AYA22A Aptamers in Regulating CD23 and CD63 Expression on B Cells: Co-Culture Study with Autologous PBMCs
2.20. Preventative and Therapeutic Basophil Activation Test
2.21. Statistical Analysis
3. Results
3.1. Design and Molecular Docking Analysis of Aptamers Targeting Ara h1 and Ara h2 Proteins Using In Silico Approaches
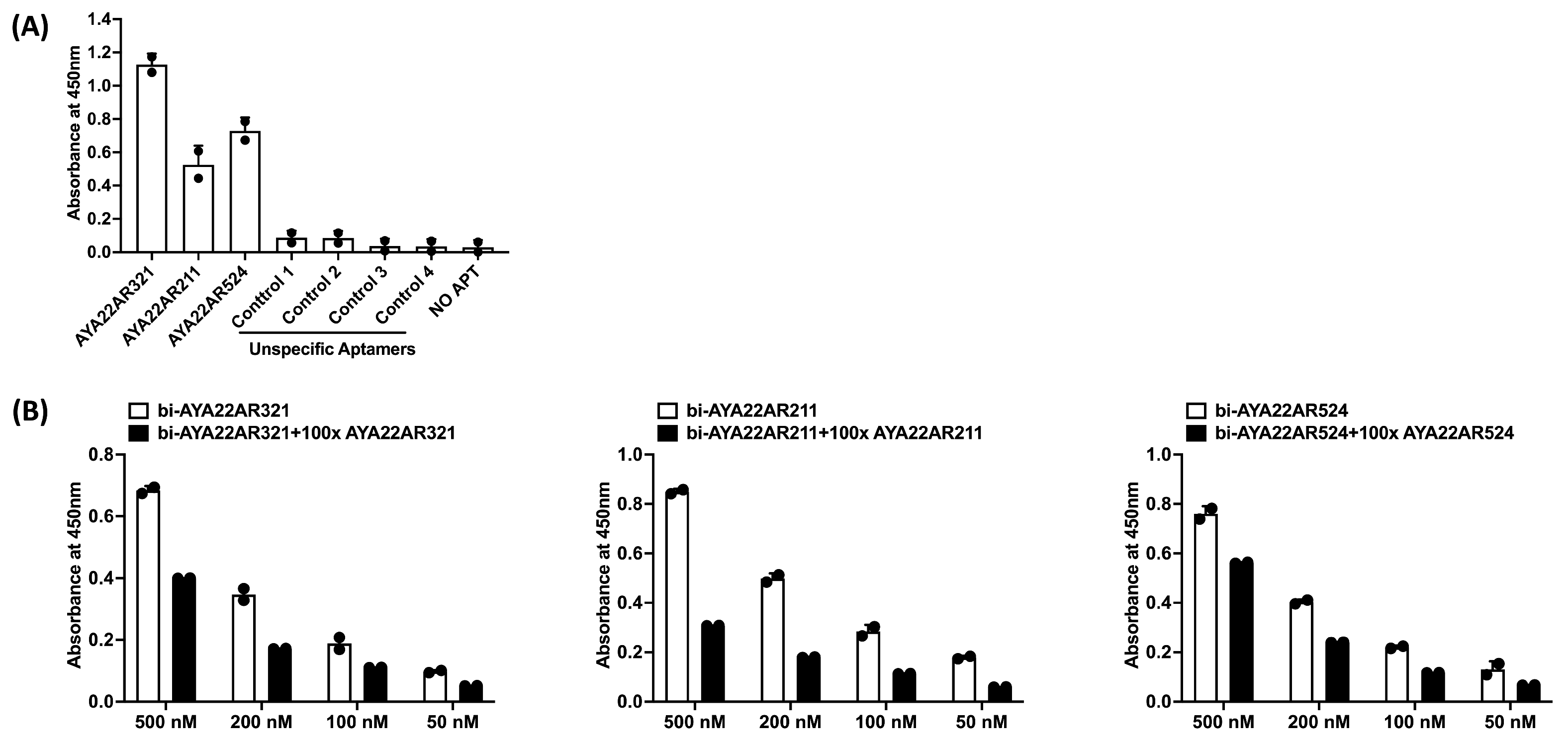
3.2. Aptamers Binding to Peanut Allergens
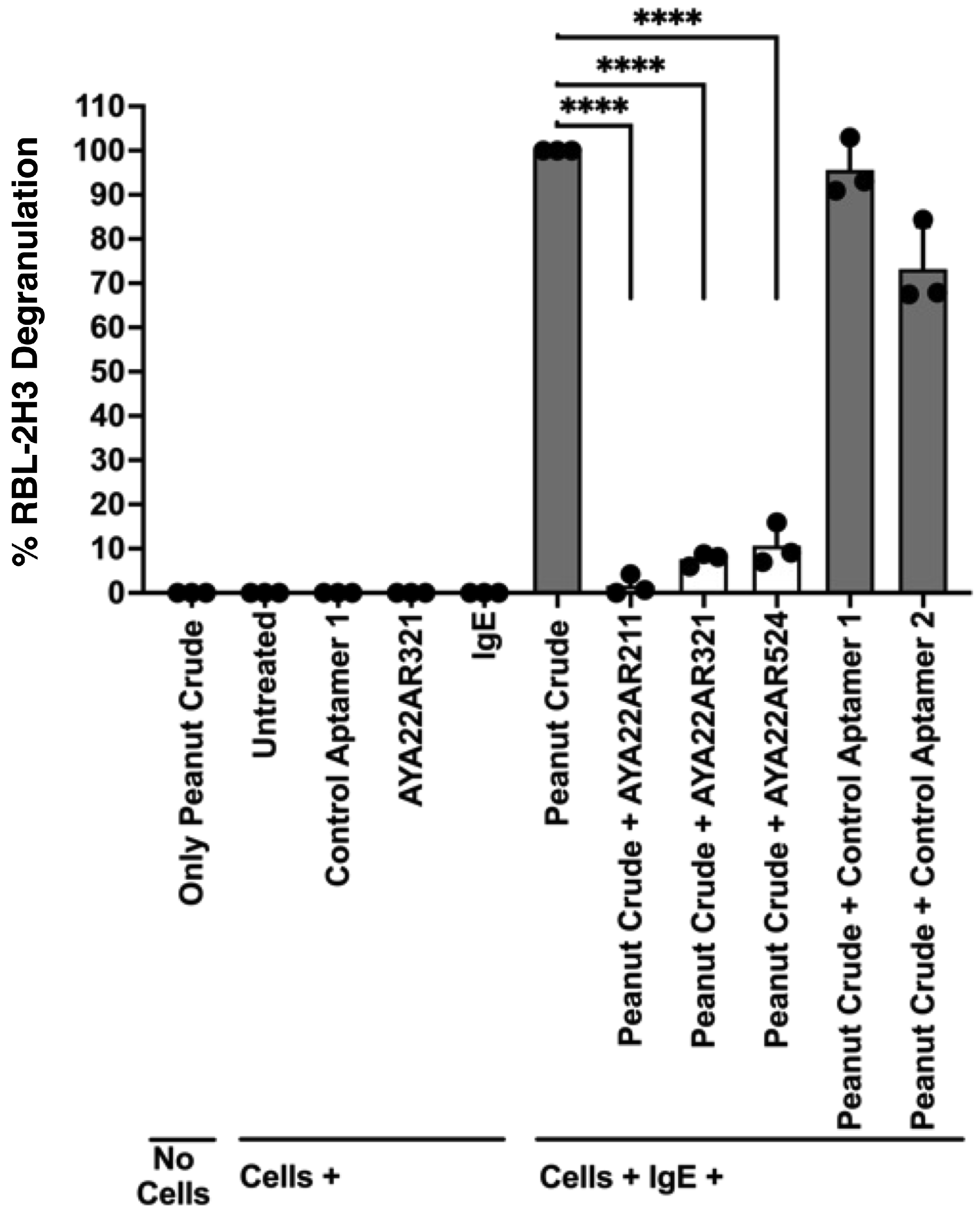
3.3. Aptamers Targeting Ara h1 and Ara h2 Effectively Inhibit IgE-Mediated Degranulation in an In Vitro RBL-2H3 Cell Allergy Model
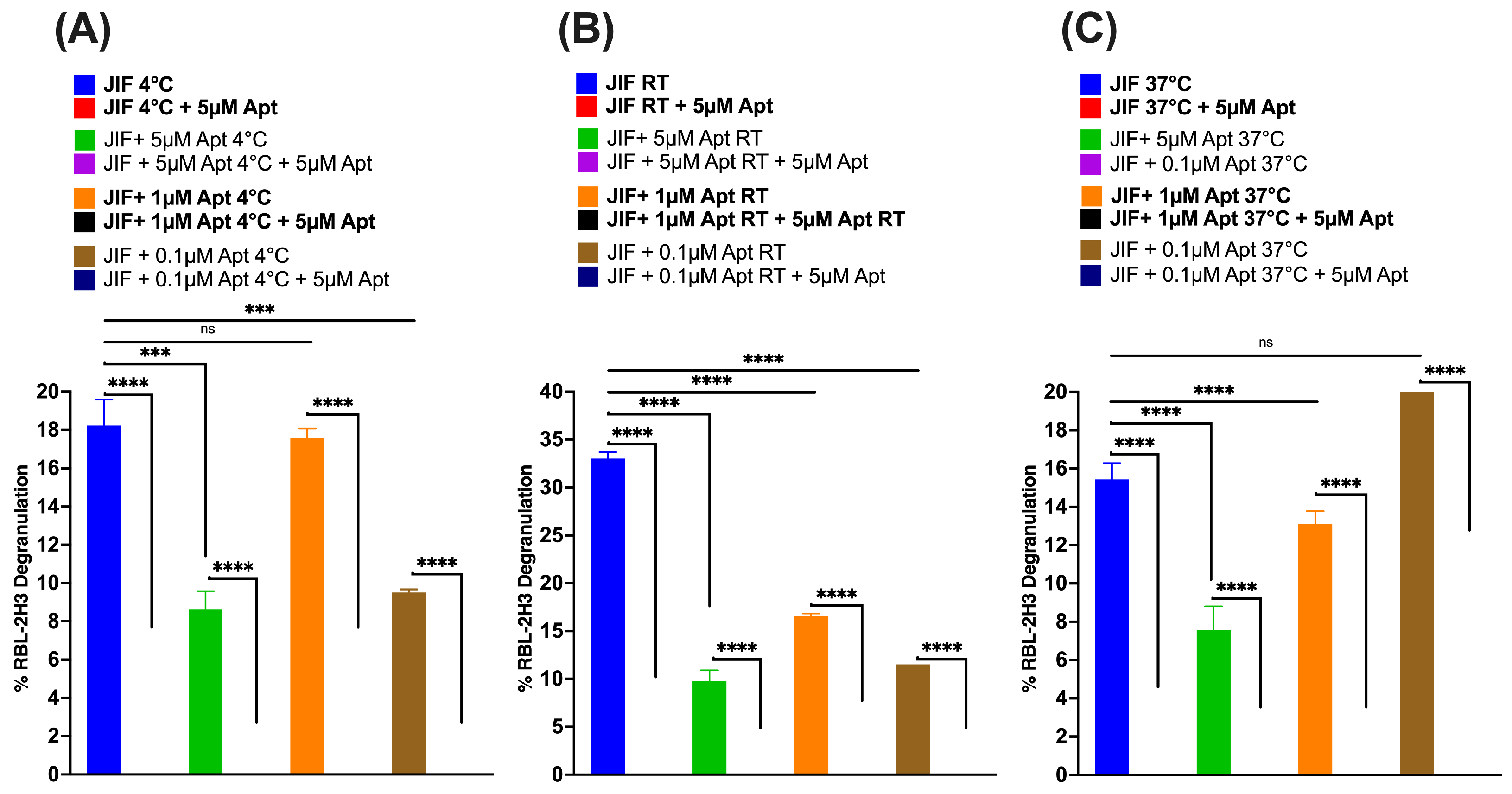
3.4. AYA22A Aptamers Demonstrate Stability and Efficacy in Reducing Degranulation Across Various Temperatures and Processing Conditions
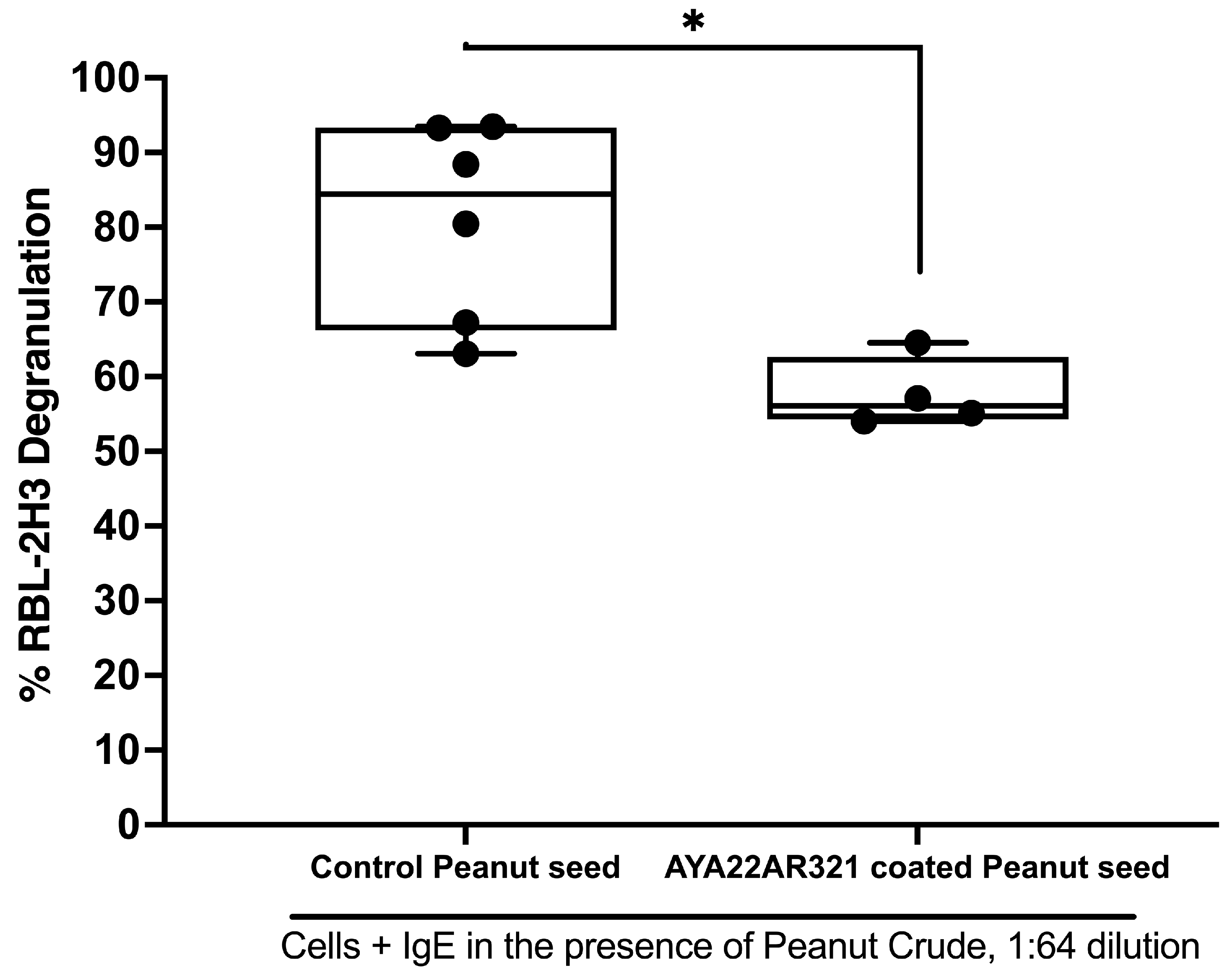
3.5. Peanut Seed Coated with AYA22A Mitigates Degranulation
3.6. AYA22A Aptamer Effectively Inhibits Peanut Butter-Induced Degranulation in RBL-2H3 Cells
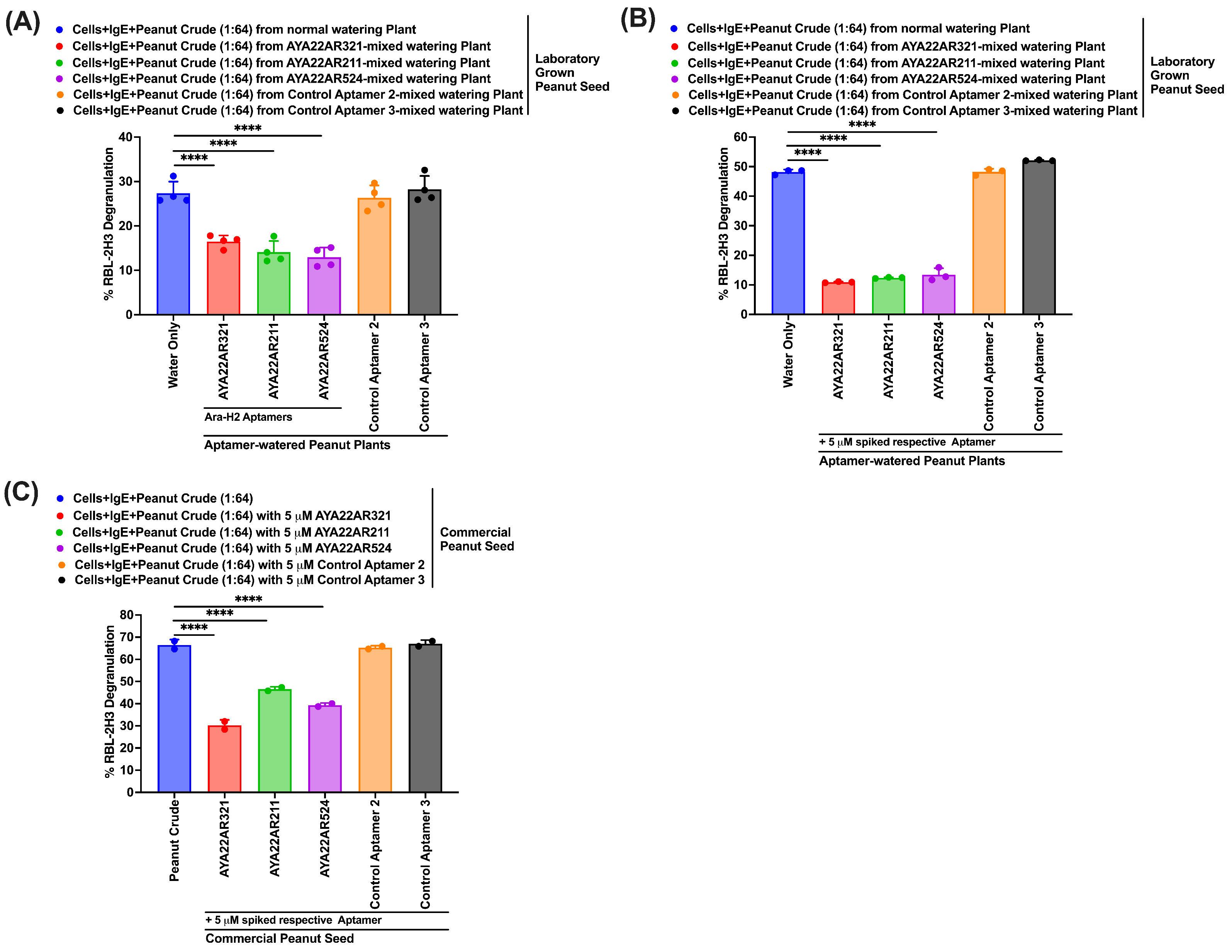
3.7. Peanut Crude Extract from the Seed of AYA22A-Watered Laboratory Peanut Plants Decreases Degranulation
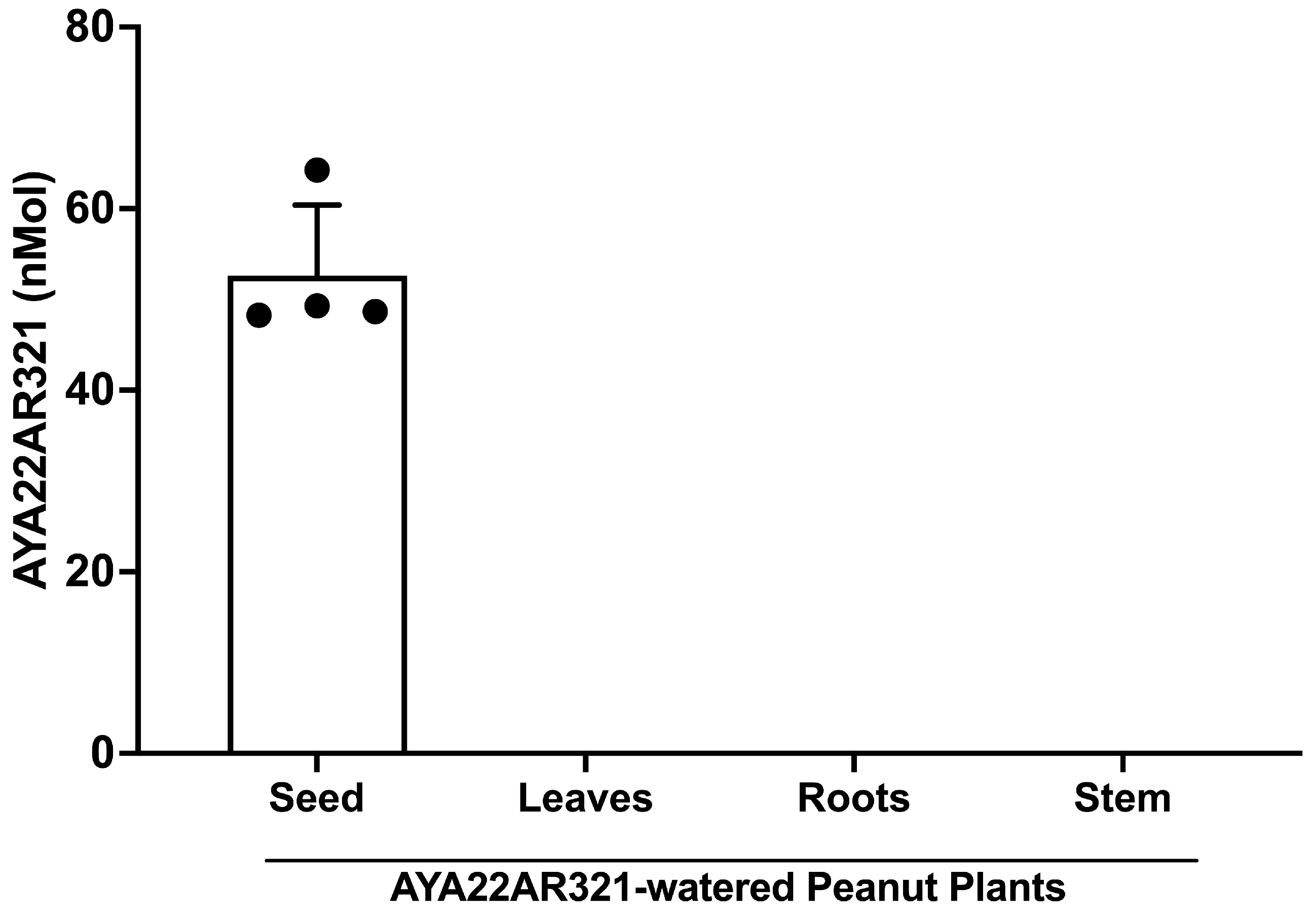
3.8. AYA22AR321 Was Detected in Peanut Crude Extracted from the Seed of AYA22AR321-Watered Laboratory Peanut Plants
3.9. AYA22A Aptamers Downregulate the Production of Th2 Cytokines and Upregulate Th1 Cytokines in an In Vitro Allergy Model
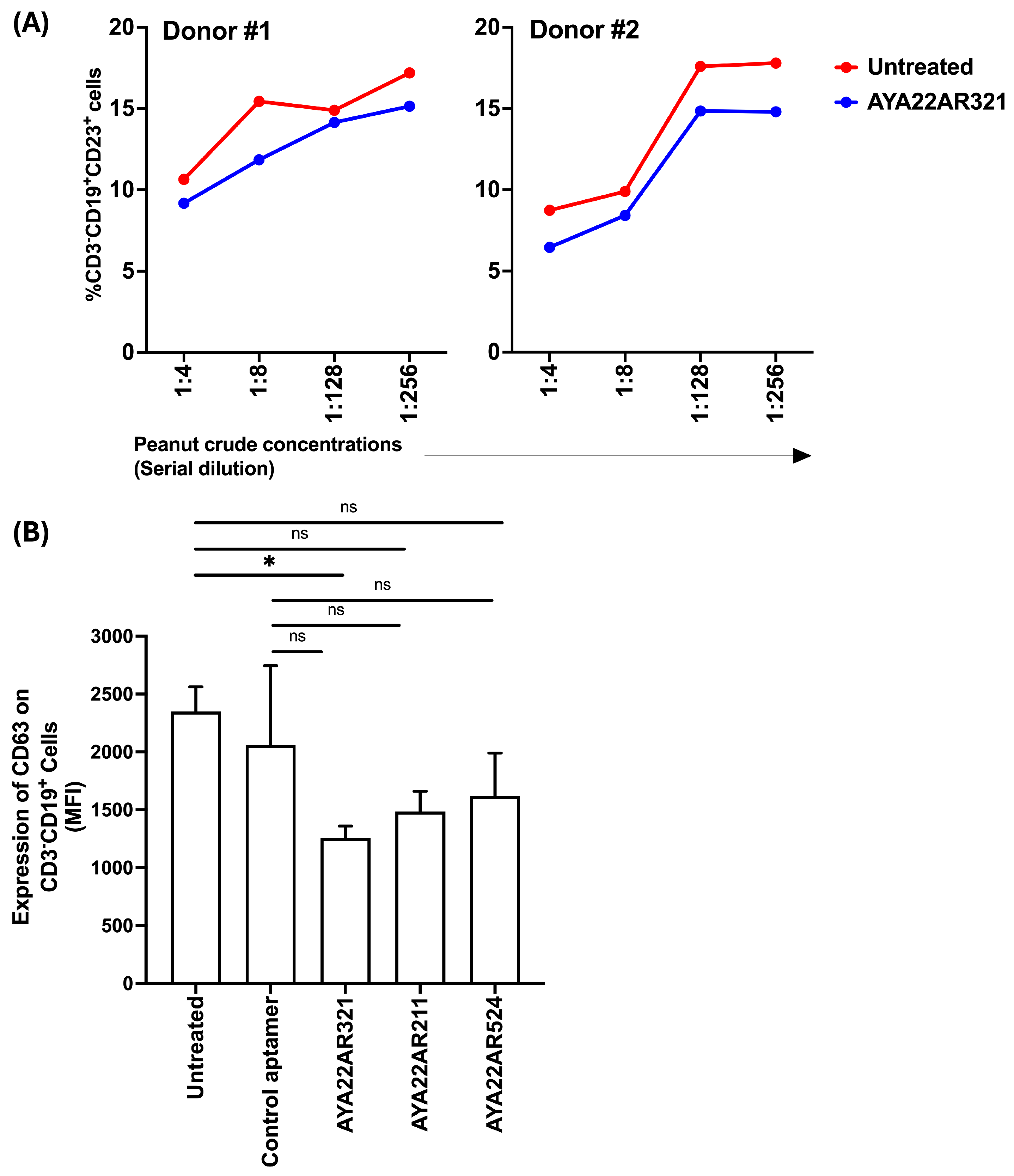
3.10. AYA22A Aptamers Neutralize the Effect of CD23 Expression on B Cells and Diminish CD63 Expression
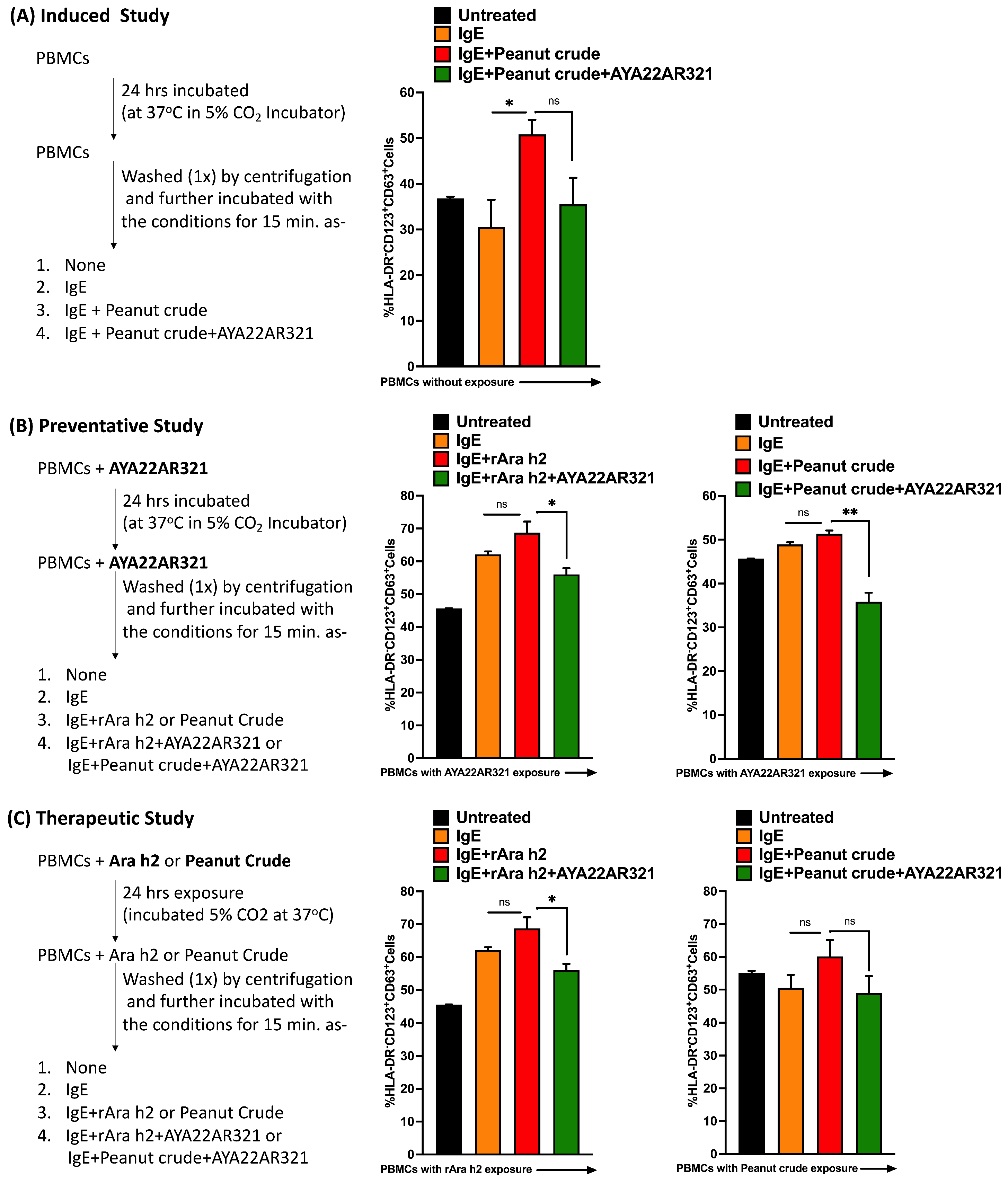
3.11. AYA22A Aptamer Therapeutically Decreases CD63 Expression in Human Basophil Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| PA | peanut allergy |
| pOIT | peanut oral immunotherapy |
| Ara h1 | Arachis hypoga 1 |
| Ara h2 | Arachis hypoga 2 |
| Ara h3 | Arachis hypoga 3 |
| SELEX | systematic evoluation of ligands by exponential enrichment |
| FBS | fetal bovin serum |
| GM-CSF | granulocyte–macrophage colony-stimulating factor |
| EPIT | epicutaneous immunotherapy |
| SLIT | sublingual immunotherapy |
References
- Jones, S.; Burks, A. Food allergy. N. Engl. J. Med. 2017, 377, 1168–1176. [Google Scholar] [CrossRef]
- Mahr, T.; Lieberman, J.; Haselkorn, T.; DamLe, V.; Ali, Y.; Chidambaram, A.; Griffin, N.; Sublett, J. Characteristics of peanut allergy diagnosis in a US health care claims database (2011–2017). J. Allergy Clin. Immunol. Pract. 2021, 9, 1683–1694. [Google Scholar] [CrossRef]
- Warren, C.; Lei, D.; Sicherer, S.; Schleimer, R.; Gupta, R. Prevalence and characteristics of peanut allergy in US adults. J. Allergy Clin. Immunol. 2021, 147, 2263–2270. [Google Scholar] [CrossRef]
- McCann, W.; Hass, S.; Norrett, K.; Cameron, A.; Etschmaier, M.; Duhig, A.; Yu, S. The Peanut Allergy Burden Study: Real-world impact of peanut allergy on resource utilization and productivity. World Allergy Organ. J. 2021, 14, 100525. [Google Scholar] [CrossRef]
- Al-Muhsen, S.; Clarke, A.; Kagan, R. Peanut allergy: An overview. CMAJ 2003, 168, 1279–1285. [Google Scholar]
- Allen, K.J.; Turner, P.J.; Pawankar, R.; Taylor, S.; Sicherer, S.; Lack, G.; Rosario, N.; Ebisawa, M.; Wong, G.; Mills, E.N.C.; et al. Precautionary labelling of foods for allergen content: Are we ready for a global framework? World Allergy Organ. J. 2014, 7, 10. [Google Scholar] [CrossRef]
- Sampson, H. Food allergy. JAMA 1997, 278, 1888–1894. [Google Scholar] [CrossRef]
- Hemmings, O.; Du Toit, G.; Radulovic, S.; Lack, G.; Santos, A. Ara h 2 is the dominant peanut allergen despite similarities with Ara h 6. J. Allergy Clin. Immunol. 2020, 146, 621–630. [Google Scholar] [CrossRef]
- Flinterman, A.; Knol, E.; Lencer, D.; Bardina, L.; Hartog Jager, C.; Lin, J.; Pasmans, S.; Bruijnzeel-Koomen, C.; Sampson, H.; Hoffen, E.; et al. Peanut epitopes for IgE and IgG4 in peanut-sensitized children in relation to severity of peanut allergy. J. Allergy Clin. Immunol. 2008, 121, 737–743. [Google Scholar] [CrossRef]
- Rambasek, T.; Kavuru, M. Omalizumab dosing via the recommended card versus use of the published formula. J. Allergy Clin. Immunol. 2006, 117, 708–709. [Google Scholar] [CrossRef]
- Fu, T.; Abbott, U.; Hatzos, C. Digestibility of food allergens and nonallergenic proteins in simulated gastric fluid and simulated intestinal fluid a comparative study. J. Agric. Food Chem. 2002, 50, 7154–7160. [Google Scholar] [CrossRef]
- Klemans, R.; Broekman, H.; Knol, E.; Bruijnzeel-Koomen, C.; Otten, H.; Pasmans, S.; Knulst, A. Ara h 2 is the best predictor for peanut allergy in adults. J. Allergy Clin. Immunol. Pract. 2013, 1, 632–638. [Google Scholar] [CrossRef]
- Sen, M.; Kopper, R.; Pons, L.; Abraham, E.; Burks, A.; Bannon, G. Protein structure plays a critical role in peanut allergen stability and may determine immunodominant IgE-binding epitopes. J. Immunol. 2002, 169, 882–887. [Google Scholar] [CrossRef]
- Schworer, S.; Kim, E. Sublingual immunotherapy for food allergy and its future directions. Immunotherapy 2020, 12, 921–931. [Google Scholar] [CrossRef]
- Kim, E.; Yang, L.; Ye, P.; Guo, R.; Li, Q.; Kulis, M.; Burks, A. Long-term sublingual immunotherapy for peanut allergy in children: Clinical and immunologic evidence of desensitization. J. Allergy Clin. Immunol. 2019, 144, 1320–1326. [Google Scholar] [CrossRef]
- Wang, J.; Li, X. Chinese herbal therapy for the treatment of food allergy. Curr. Allergy Asthma Rep. 2012, 12, 332338. [Google Scholar] [CrossRef]
- Hu, J.; Chen, J.; Ye, L.; Cai, Z.; Sun, J.; Ji, K. Anti-IgE therapy for IgE-mediated allergic diseases: From neutralizing IgE antibodies to eliminating IgE+ B cells. Clin. Transl. Allergy 2018, 8, 27. [Google Scholar] [CrossRef]
- Fowler, J.; Lieberman, J. Update on clinical research for food allergy treatment. Front. Allergy 2023, 4, 1154541. [Google Scholar] [CrossRef]
- Keefe, A.; Pai, S.; Ellington, A. Aptamers as therapeutics. Nat. Rev. Drug Discov. 2010, 9, 537–550. [Google Scholar] [CrossRef]
- Darmostuk, M.; Rimpelova, S.; Gbelcova, H.; Ruml, T. Current approaches in SELEX: An update to aptamer selection technology. Biotechnol. Adv. 2015, 33, 1141–1161. [Google Scholar] [CrossRef]
- Thiel, K.; Giangrande, P. Therapeutic applications of DNA and RNA aptamers. Oligonucleotides 2009, 19, 209–222. [Google Scholar] [CrossRef]
- Jayasena, S. Aptamers: An emerging class of molecules that rival antibodies in diagnostics. Clin. Chem. 1999, 45, 1628–1650. [Google Scholar] [CrossRef]
- Wieg, T.; Williams, P.; Dreskin, S.; Jouvin, M.; Kinet, J.; Tasset, D. High-affinity oligonucleotide ligands to human IgE inhibit binding to Fc epsilon receptor I. J. Immunol. 1996, 157, 221–230. [Google Scholar]
- Poongavanam, M.; Kisley, L.; Kourentzi, K.; Landes, C.; Willson, R. Ensemble and single-molecule biophysical characterization of D17. 4 DNA aptamer–IgE interactions. Biochim. Biophys. Acta-(Bba)-Proteins Proteom. 2016, 1864, 154–164. [Google Scholar] [CrossRef]
- Ayass, M.; Tripathi, T.; Griko, N.; Okyay, T.; Ramankutty Nair, R.; Zhang, J.; Zhu, K.; Melendez, K.; Pashkov, V.; Abi-Mosleh, L. Dual Checkpoint Aptamer Immunotherapy: Unveiling Tailored Cancer Treatment Targeting CTLA-4 and NKG2A. Cancers 2024, 16, 1041. [Google Scholar] [CrossRef]
- Passante, E.; Ehrhardt, C.; Sheridan, H.; Frankish, N. RBL-2H3 cells are an imprecise model for mast cell mediator release. Inflamm. Res. 2009, 58, 611–618. [Google Scholar] [CrossRef]
- Kulczycki, A., Jr.; Isersky, C.; Metzger, H. The interaction of IgE with rat basophilic leukemia cells: I. Evidence for specific binding of IgE. J. Exp. Med. 1974, 139, 600–616. [Google Scholar] [CrossRef]
- Barsumian, E.; Isersky, C.; Petrino, M.; Siraganian, R. IgE-induced histamine release from rat basophilic leukemia cell lines: Isolation of releasing and nonreleasing clones. Eur. J. Immunol. 1981, 11, 317–323. [Google Scholar] [CrossRef]
- Zuker, M. Mfold web server for nucleic acid folding and hybridization prediction. Nucleic Acids Res. 2003, 31, 3406–3415. [Google Scholar] [CrossRef]
- Antczak, M.; Popenda, M.; Zok, T.; Sarzynska, J.; Ratajczak, T.; Tomczyk, K.; Adamiak, R.; Szachniuk, M. New functionality of RNAComposer: An application to shape the axis of miR160 precursor structure. Acta Biochim. Pol. 2016, 63, 737–744. [Google Scholar] [CrossRef]
- Popenda, M.; Szachniuk, M.; Antczak, M.; Purzycka, K.; Lukasiak, P.; Bartol, N.; Blazewicz, J.; Adamiak, R. Automated 3D structure composition for large RNAs. Nucleic Acids Res. 2012, 40, e112. [Google Scholar] [CrossRef]
- Honorato, R.; Koukos, P.; Jiménez-Garcia, B.; Tsaregorodtsev, A.; Verlato, M.; Giachetti, A.; Rosato, A.; Bonvin, A. Structural biology in the clouds: The WeNMR-EOSC ecosystem. Front. Mol. Biosci. 2021, 8, 729513. [Google Scholar] [CrossRef]
- Van Zundert, G.; Rodrigues, J.; Trellet, M.; Schmitz, C.; Kastritis, P.; Karaca, E.; Melquiond, A.; Dijk, M.; De Vries, S.; Bonvin, A. The HADDOCK2. 2 web server: User-friendly integrative modeling of biomolecular complexes. J. Mol. Biol. 2016, 428, 720–725. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Židek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Morita, Y.; Siraganian, R. Inhibition of IgE-mediated histamine release from rat basophilic leukemia cells and rat mast cells by inhibitors of transmethylation. J. Immunol. 1981, 127, 1339–1344. [Google Scholar] [CrossRef]
- Koppelman, S.; Smits, M.; Tomassen, M.; De Jong, G.; Baumert, J.; Taylor, S.; Witkamp, R.; Veldman, R.; Pieters, R.; Wichers, H. Release of major peanut allergens from their matrix under various pH and simulated saliva conditions—Ara h2 and ara h6 are readily bio-accessible. Nutrients 2018, 10, 1281. [Google Scholar] [CrossRef]
- Maleki, S.; Viquez, O.; Jacks, T.; Dodo, H.; Champagne, E.; Chung, S.; Landry, S. The major peanut allergen, Ara h 2, functions as a trypsin inhibitor, and roasting enhances this function. J. Allergy Clin. Immunol. 2003, 112, 190–195. [Google Scholar] [CrossRef]
- Yoo, J.; Kim, N.; Seo, J.; Kim, S.; Lee, S.; Kim, S.; Kim, H.; Lee, S.; Kim, M. Inhibitory effects of mulberry fruit extract in combination with naringinase on the allergic response in IgE-activated RBL-2H3 cells. Int. J. Mol. Med. 2014, 33, 469–477. [Google Scholar] [CrossRef]
- Staats, H.; Kirwan, S.; Choi, H.; Shelburne, C.; Abraham, S.; Leung, G.; Chen, D. A mast cell degranulation screening assay for the identification of novel mast cell activating agents. Medchemcomm 2013, 4, 88–94. [Google Scholar] [CrossRef]
- Ayass, M.; Tripathi, T.; Griko, N.; Pashkov, V.; Dai, J.; Zhang, J.; Herbert, F.; Ramankutty Nair, R.; Okyay, T.; Zhu, K.; et al. Highly efficacious and safe neutralizing DNA aptamer of SARS-CoV-2 as an emerging therapy for COVID-19 disease. Virol. J. 2022, 19, 227. [Google Scholar] [CrossRef]
- Hiasa, M.; Abe, M.; Nakano, A.; Oda, A.; Amou, H.; Kido, S.; Takeuchi, K.; Kagawa, K.; Yata, K.; Hashimoto, T.; et al. GM-CSF and IL-4 induce dendritic cell differentiation and disrupt osteoclastogenesis through M-CSF receptor shedding by up-regulation of TNF-ɑconverting enzyme (TACE). Blood J. Am. Soc. Hematol. 2009, 114, 4517–4526. [Google Scholar]
- Tripathi, T.; Yin, W.; Xue, Y.; Zurawski, S.; Fujita, H.; Hanabuchi, S.; Liu, Y.; Oh, S.; Joo, H. Central Roles of OX40L–OX40 Interaction in the Induction and Progression of Human T Cell–Driven Acute Graft-versus-Host Disease. Immunohorizons 2019, 3, 110–120. [Google Scholar] [CrossRef] [PubMed]
- Selb, R.; Eckl-Dorna, J.; Neunkirchner, A.; Schmetterer, K.; Marth, K.; Gamper, J.; Jahn-Schmid, B.; Pickl, W.; Valenta, R.; Niederberger, V. CD23 surface density on B cells is associated with IgE levels and determines IgE-facilitated allergen uptake, as well as activation of allergen-specific T cells. J. Allergy Clin. Immunol. 2017, 139, 290–299. [Google Scholar] [CrossRef] [PubMed]
- Engeroff, P.; Caviezel, F.; Mueller, D.; Thoms, F.; Bachmann, M.; Vogel, M. CD23 provides a noninflammatory pathway for IgE-allergen complexes. J. Allergy Clin. Immunol. 2020, 145, 301–311. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Liu, J.; Truong, T.; Zukin, E.; Chen, W.; Saxon, A. Blocking allergic reaction through targeting surface-bound IgE with low-affinity anti-IgE antibodies. J. Immunol. 2017, 198, 3823–3834. [Google Scholar] [CrossRef] [PubMed]
- Kamath, S.; Bublin, M.; Kitamura, K.; Matsui, T.; Ito, K.; Lopata, A. Cross-reactive epitopes and their role in food allergy. J. Allergy Clin. Immunol. 2023, 151, 1178–1190. [Google Scholar] [CrossRef] [PubMed]
- Paungfoo-Lonhienne, C.; Lonhienne, T.; Mudge, S.; Schenk, P.; Christie, M.; Carroll, B.; Schmidt, S. DNA is taken up by root hairs and pollen, and stimulates root and pollen tube growth. Plant Physiol. 2010, 153, 799–805. [Google Scholar] [CrossRef] [PubMed]
- Purnamasari, R.; Sudadi, U.; Santosa, D. A Review: Is Cinderella’s story of self-DNA extracellular effect towards plant growth real? IOP Conf. Ser. Earth Environ. Sci. 2021, 824, 012026. [Google Scholar] [CrossRef]
- Humeniuk, P.; Dubiela, P.; Hoffmann-Sommergruber, K. Dendritic cells and their role in allergy: Uptake, proteolytic processing and presentation of allergens. Int. J. Mol. Sci. 2017, 18, 1491. [Google Scholar] [CrossRef]
- Castenmiller, C.; Nagy, N.; Kroon, P.; Auger, L.; Desgagnés, R.; Martel, C.; Mire, L.; Morel, B.; Roberge, J.; Stordeur, V.; et al. A novel peanut allergy immunotherapy: Plant-based enveloped Ara h 2 Bioparticles activate dendritic cells and polarize T cell responses to Th1. World Allergy Organ. J. 2023, 16, 100839. [Google Scholar] [CrossRef]
- Peyron, I.; Hartholt, R.; Pedró-Cos, L.; Alphen, F.; Ten Brinke, A.; Lardy, N.; Meijer, A.; Voorberg, J. Comparative profiling of HLA-DR and HLA-DQ associated factor VIII peptides presented by monocyte-derived dendritic cells. Haematologica 2018, 103, 172. [Google Scholar] [CrossRef] [PubMed]
- Ashjaei, K.; Bublin, M.; Smole, U.; Lengger, N.; Hafner, C.; Breiteneder, H.; Wagner, S.; Hoffmann-Sommergruber, K. Differential T-helper cell polarization after allergen-specific stimulation of autologous dendritic cells in polysensitized allergic patients. Int. Arch. Allergy Immunol. 2015, 166, 97–106. [Google Scholar] [CrossRef] [PubMed]
- López-Garcia, L.; Castro-Manrreza, M. TNF-ɑ and IFN-ɣ participate in improving the immunoregulatory capacity of mesenchymal stem/stromal cells: Importance of cell-cell contact and extracellular vesicles. Int. J. Mol. Sci. 2021, 22, 9531. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Li, Y.; Zeng, J.; Chang, N.; Cheng, Y.; Zhen, X.; Zhong, D.; Chen, R.; Ma, G.; Wang, Y. Mesenchymal Stem/Stromal Cells in Asthma Therapy: Mechanisms and Strategies for Enhancement. Cell Transplant. 2023, 32, 09636897231180128. [Google Scholar] [CrossRef] [PubMed]
- Borne, G.; Daniel, C.; Wagner, M.; Plaisance, C.; Nolen, A.; Kelkar, R.; Ahmadzadeh, S.; Myrcik, D.; Shekoohi, S.; Kaye, A.; et al. Palforzia for Peanut Allergy: A Narrative Review and Update on a Novel Immunotherapy. Cureus 2023, 15, e50485. [Google Scholar] [PubMed]
- From the Medical Letter on Drugs and Therapeutics. Peanut Allergen Powder (Palforzia). Available online: https://jamanetwork.com/journals/jama/article-abstract/2768131 (accessed on 15 March 2023).
- Busse, W.; Corren, J.; Lanier, B.; McAlary, M.; Fowler-Taylor, A.; Della Cioppa, G.; As, A.; Gupta, N. Omalizumab, anti-IgE recombinant humanized monoclonal antibody, for the treatment of severe allergic asthma. J. Allergy Clin. Immunol. 2001, 108, 184–190. [Google Scholar] [CrossRef]
- Belliveau, P. Omalizumab: A monoclonal anti-IgE antibody. Medscape Gen. Med. 2005, 7, 27. [Google Scholar]
- Massanari, M.; Holgate, S.; Busse, W.; Jimenez, P.; Kianifard, F.; Zeldin, R. Effect of omalizumab on peripheral blood eosinophilia in allergic asthma. Respir. Med. 2010, 104, 188–196. [Google Scholar] [CrossRef] [PubMed]
- Azzano, P.; Paquin, M.; Langlois, A.; Morin, C.; Parizeault, G.; Lacombe-Barrios, J.; Samaan, K.; Graham, F.; Paradis, L.; Des Roches, A.; et al. Determinants of omalizumab dose–related efficacy in oral immunotherapy: Evidence from a cohort of 181 patients. J. Allergy Clin. Immunol. 2021, 147, 233–243. [Google Scholar] [CrossRef]
- Tirumalasetty, J.; Barshow, S.; Kost, L.; Morales, L.; Sharma, R.; Lazarte, C.; Nadeau, K. Peanut allergy: Risk factors, immune mechanisms, and best practices for oral immunotherapy success. Expert Rev. Clin. Immunol. 2023, 19, 785–795. [Google Scholar]
- Zhou, Y.; Wang, J.; Yang, X.; Lin, D.; Gao, Y.; Su, Y.; Yang, S.; Zhang, Y.; Zheng, J. Peanut allergy, allergen composition, and methods of reducing allergenicity: A review. Int. J. Food Sci. 2013, 2013, 909140. [Google Scholar] [CrossRef] [PubMed]
- Palladino, C.; Breiteneder, H. Peanut allergens. Mol. Immunol. 2018, 100, 58–70. [Google Scholar] [CrossRef] [PubMed]
- Maleki, S.; Chung, S.; Champagne, E.; Raufman, J. The effects of roasting on the allergenic properties of peanut proteins. J. Allergy Clin. Immunol. 2000, 106, 763–768. [Google Scholar] [CrossRef] [PubMed]
- Moghaddam, A.; Hillson, W.; Noti, M.; Gartlan, K.; Johnson, S.; Thomas, B.; Artis, D.; Sattentau, Q. Dry roasting enhances peanut-induced allergic sensitization across mucosal and cutaneous routes in mice. J. Allergy Clin. Immunol. 2014, 134, 1453–1456. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Wang, S.; Sun, X.; Deng, Z.; Niu, B.; Chen, Q. Study on mechanism of increased allergenicity induced by Ara h 3 from roasted peanut using bone marrow-derived dendritic cells. Food Sci. Hum. Wellness 2023, 12, 755–764. [Google Scholar] [CrossRef]
- Li, X.; Huang, C.; Schofield, B.; Burks, A.; Bannon, G.; Kim, K.; Huang, S.; Sampson, H. Strain-dependent induction of allergic sensitization caused by peanut allergen DNA immunization in mice. J. Immunol. 1999, 162, 3045–3052. [Google Scholar] [CrossRef] [PubMed]
- National Library of Medicine. Clinical Trials: A Study to Evaluate Safety, Tolerability and Immune Response in Adults Allergic to Peanut after Receiving Intradermal or Intramuscular Administration of ASP0892 (ARA-LAMP-vax), a Single Multivalent Peanut (Ara h1, h2, h3) Lysosomal Associated Membrane Protein DNA Plasmid Vaccine. Available online: https://clinicaltrials.gov/study/NCT02851277 (accessed on 15 March 2023).
- Mayorga, C.; Palomares, F.; Cañas, J.; Pérez-Sánchez, N.; Núñez, R.; Torres, M.; Gómez, F. New insights in therapy for food allergy. Foods 2021, 10, 1037. [Google Scholar] [CrossRef] [PubMed]
- Ellenbogen, Y.; Jiménez-Saiz, R.; Spill, P.; Chu, D.; Waserman, S.; Jordana, M. The initiation of Th2 immunity towards food allergens. Int. J. Mol. Sci. 2018, 19, 1447. [Google Scholar] [CrossRef]
- Wisniewski, J.; Commins, S.; Agrawal, R.; Hulse, K.; Yu, M.; Cronin, J.; Heymann, P.; Pomes, A.; Platts-Mills, T.; Workman, L.; et al. Analysis of cytokine production by peanut-reactive T cells identifies residual Th2 effectors in highly allergic children who received peanut oral immunotherapy. Clin. Exp. Allergy 2015, 45, 1201–1213. [Google Scholar] [CrossRef]
- Czolk, R.; Klueber, J.; Sørensen, M.; Wilmes, P.; Codreanu-Morel, F.; Skov, P.; Hilger, C.; Bindslev-Jensen, C.; Ollert, M.; Kuehn, A. IgE-mediated peanut allergy: Current and novel predictive biomarkers for clinical phenotypes using multi-omics approaches. Front. Immunol. 2021, 11, 594350. [Google Scholar] [CrossRef]
- Aguilera-Insunza, R.; Venegas, L.; Iruretagoyena, M.; Rojas, L.; Borzutzky, A. Role of dendritic cells in peanut allergy. Expert Rev. Clin. Immunol. 2018, 14, 367–378. [Google Scholar] [CrossRef] [PubMed]
- Plewako, H.; Wosińska, K.; Arvidsson, M.; Björker, J.; Skov, P.; Hakansson, L.; Rak, S. Basophil interleukin 4 and interleukin 13 production is suppressed during the early phase of rush immunotherapy. Int. Arch. Allergy Immunol. 2006, 141, 346–353. [Google Scholar] [CrossRef] [PubMed]
- Sampath, V.; Nadeau, K.C. Newly identified T cell subsets in mechanistic studies of food immunotherapy. J. Clin. Investig. 2019, 129, 1431–1440. [Google Scholar] [CrossRef] [PubMed]
- Berin, M.; Agashe, C.; Burks, A.; Chiang, D.; Davidson, W.; Dawson, P.; Grishin, A.; Henning, A.; Jones, S.; Kim, E.; et al. Allergen-specific T cells and clinical features of food allergy: Lessons from CoFAR immunotherapy cohorts. J. Allergy Clin. Immunol. 2022, 149, 1373–1382. [Google Scholar] [CrossRef] [PubMed]
- Du Toit, G.; Roberts, G.; Sayre, P.; Bahnson, H.; Radulovic, S.; Santos, A.; Brough, H.; Phippard, D.; Basting, M.; Feeney, M.; et al. Randomized trial of peanut consumption in infants at risk for peanut allergy. N. Engl. J. Med. 2015, 372, 803–813. [Google Scholar] [CrossRef]
- Du Toit, G.; Huffaker, M.; Radulovic, S.; Feeney, M.; Fisher, H.; Byron, M.; Dunaway, L.; Calatroni, A.; Johnson, M.; Foong, R.; et al. Follow-up to adolescence after early peanut introduction for allergy prevention. NEJM Evid. 2024, 3, EVIDoa2300311. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ayass, M.A.; Tripathi, T.; Griko, N.; Ramankutty Nair, R.; Okyay, T.; Zhang, J.; Zhu, K.; Melendez, K.; Pashkov, V.; Abi-Mosleh, L. AYA22A Aptamers Mitigate Peanut Allergenicity: Insights from Degranulation Assays and Modulating Immune Responses. Allergies 2024, 4, 94-123. https://doi.org/10.3390/allergies4030008
Ayass MA, Tripathi T, Griko N, Ramankutty Nair R, Okyay T, Zhang J, Zhu K, Melendez K, Pashkov V, Abi-Mosleh L. AYA22A Aptamers Mitigate Peanut Allergenicity: Insights from Degranulation Assays and Modulating Immune Responses. Allergies. 2024; 4(3):94-123. https://doi.org/10.3390/allergies4030008
Chicago/Turabian StyleAyass, Mohamad Ammar, Trivendra Tripathi, Natalya Griko, Ramya Ramankutty Nair, Tutku Okyay, Jin Zhang, Kevin Zhu, Kristen Melendez, Victor Pashkov, and Lina Abi-Mosleh. 2024. "AYA22A Aptamers Mitigate Peanut Allergenicity: Insights from Degranulation Assays and Modulating Immune Responses" Allergies 4, no. 3: 94-123. https://doi.org/10.3390/allergies4030008
APA StyleAyass, M. A., Tripathi, T., Griko, N., Ramankutty Nair, R., Okyay, T., Zhang, J., Zhu, K., Melendez, K., Pashkov, V., & Abi-Mosleh, L. (2024). AYA22A Aptamers Mitigate Peanut Allergenicity: Insights from Degranulation Assays and Modulating Immune Responses. Allergies, 4(3), 94-123. https://doi.org/10.3390/allergies4030008