
Post-Injury and Resolution Response to Repetitive Inhalation Exposure to Agricultural Organic Dust in Mice

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
2.1. Organic Dust Extract
2.2. Animals
2.3. Animal Exposure Model
2.4. Bronchoalveolar Lavage Fluid Cell Analysis
2.5. Histopathology
2.6. Flow Cytometry Phenotyping of Whole Lung Cells
2.7. Cell Sorting and Leukocyte Ex Vivo Stimulation
2.8. Apoptosis Immunohistochemistry Assay
2.9. Serum
2.10. Statistical Methods
3. Results
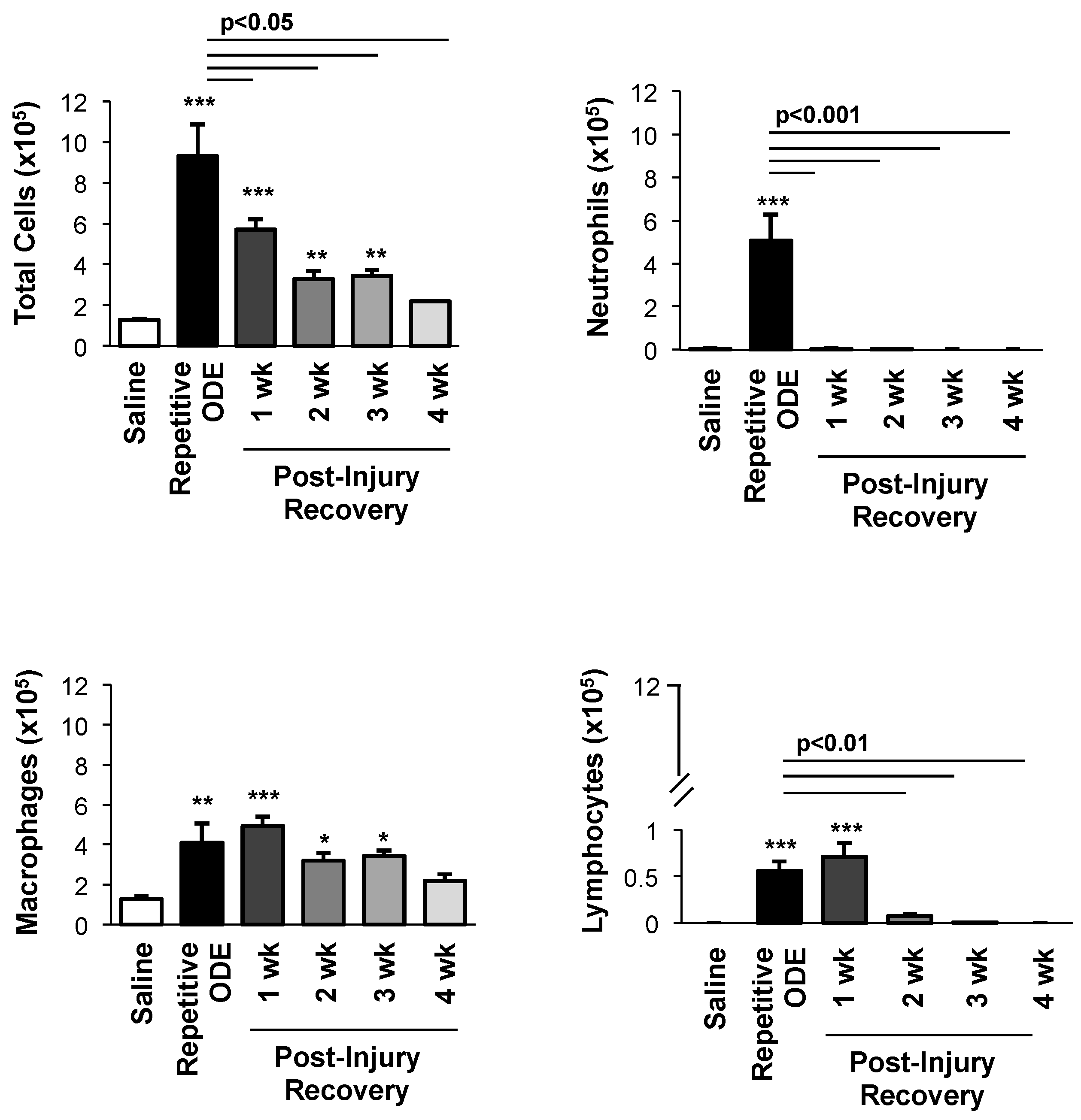
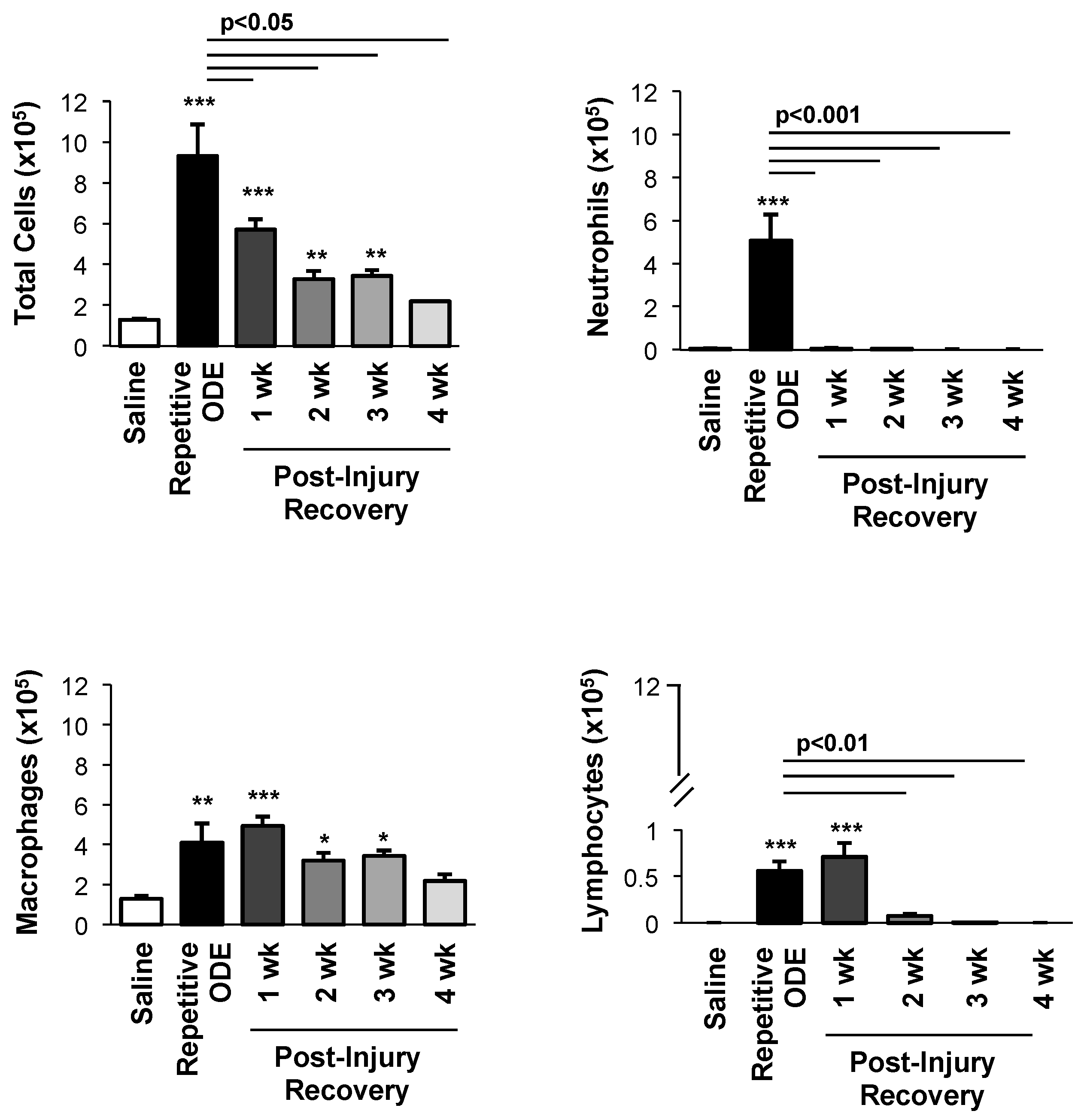
3.1. Airway Cellular Influx in BALF after Repetitive ODE Treatments and Following the 1–4 Week Post-Injury Recovery Periods
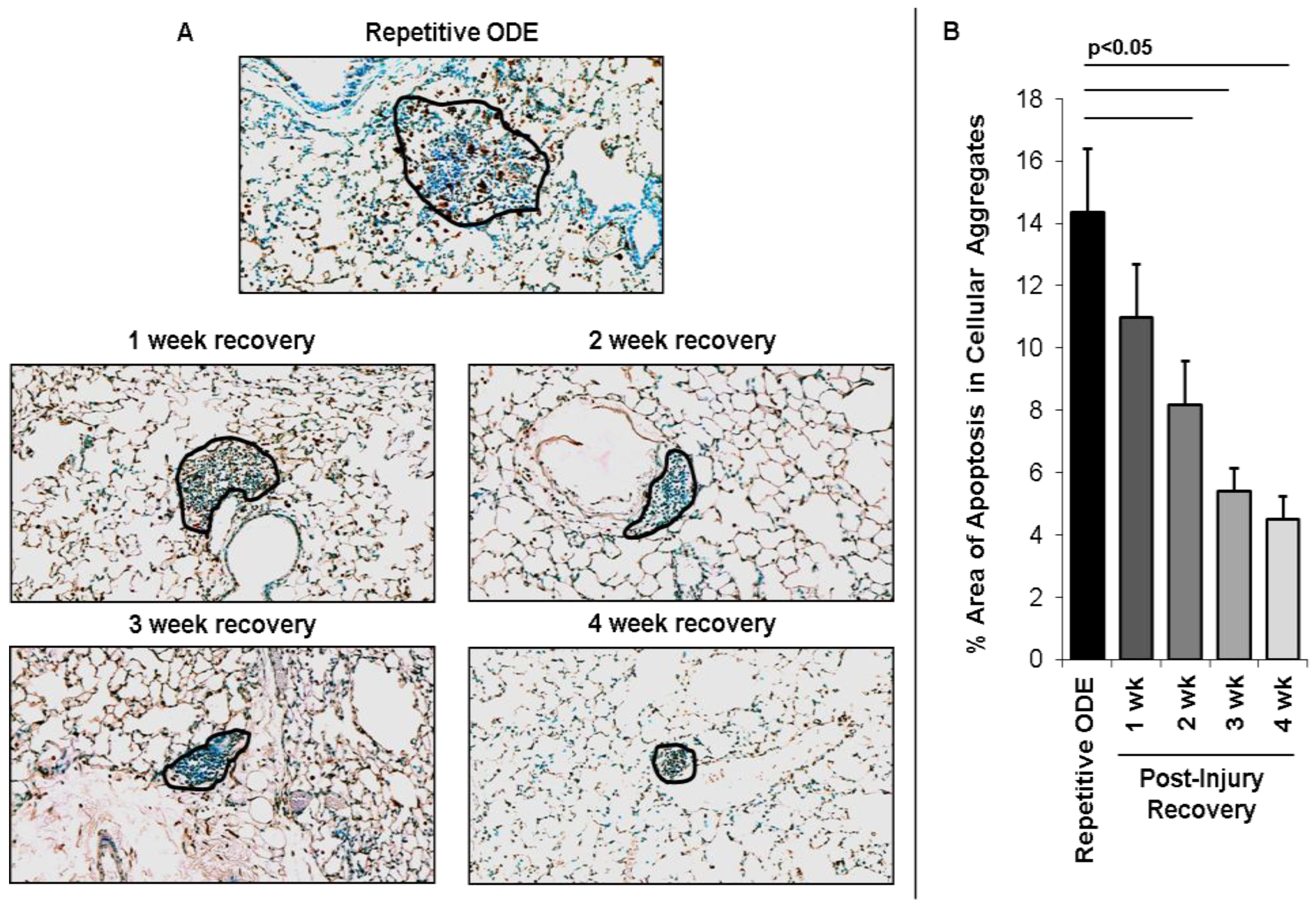
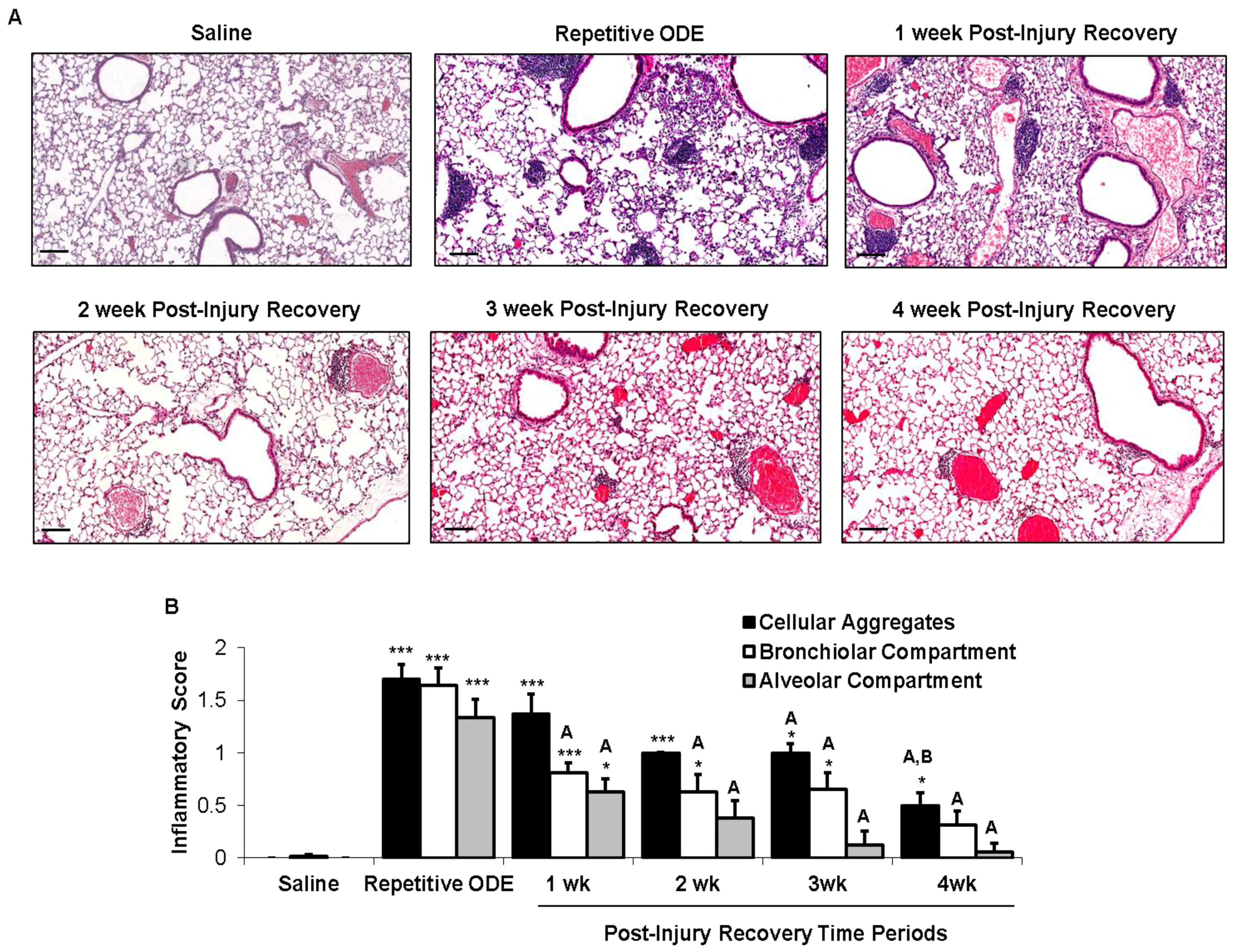
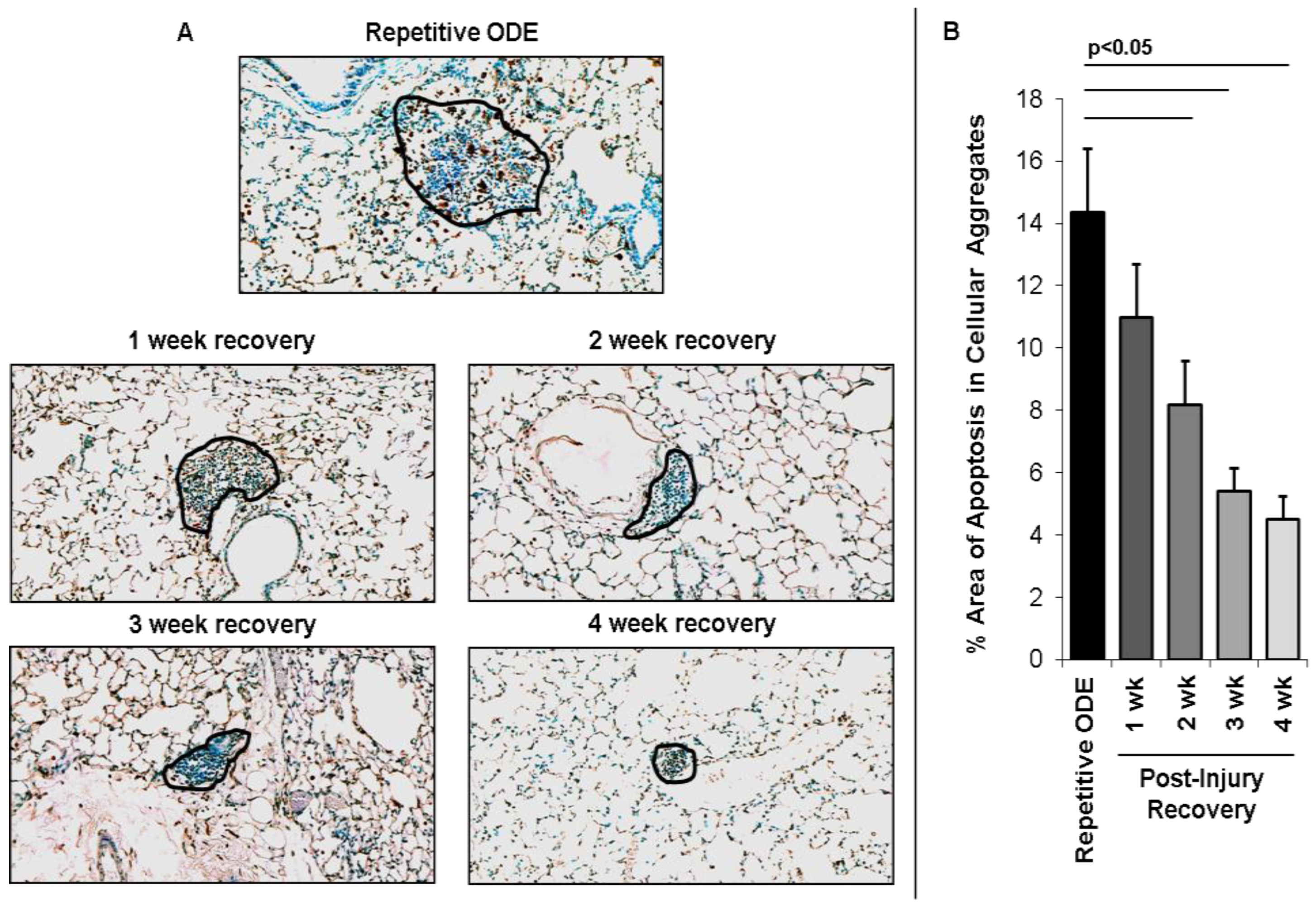
3.2. Recovery from Repetitive ODE Exposure Is Marked by Persistence of Cellular Aggregates for up to 4 Weeks in the Lung Tissue
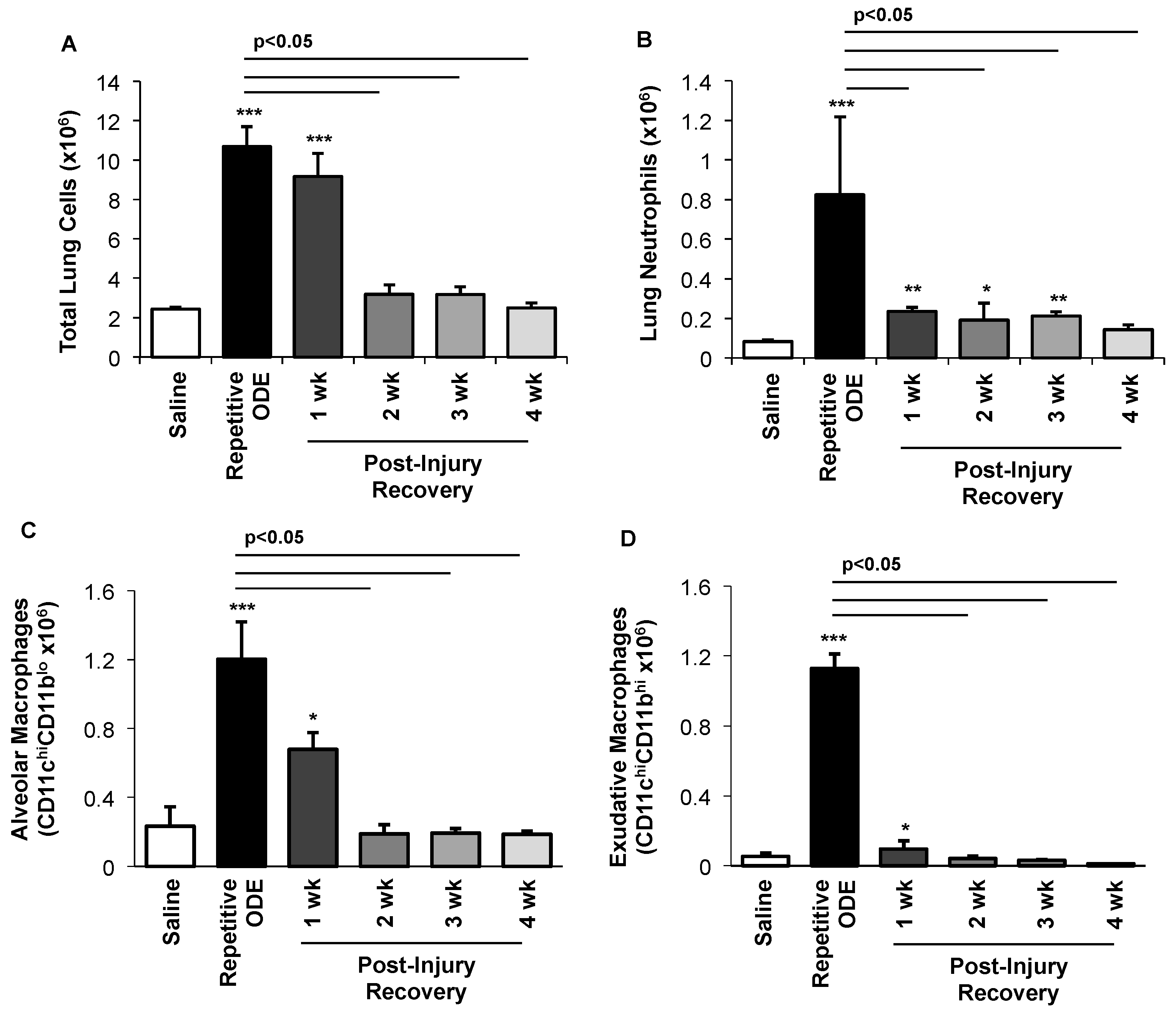
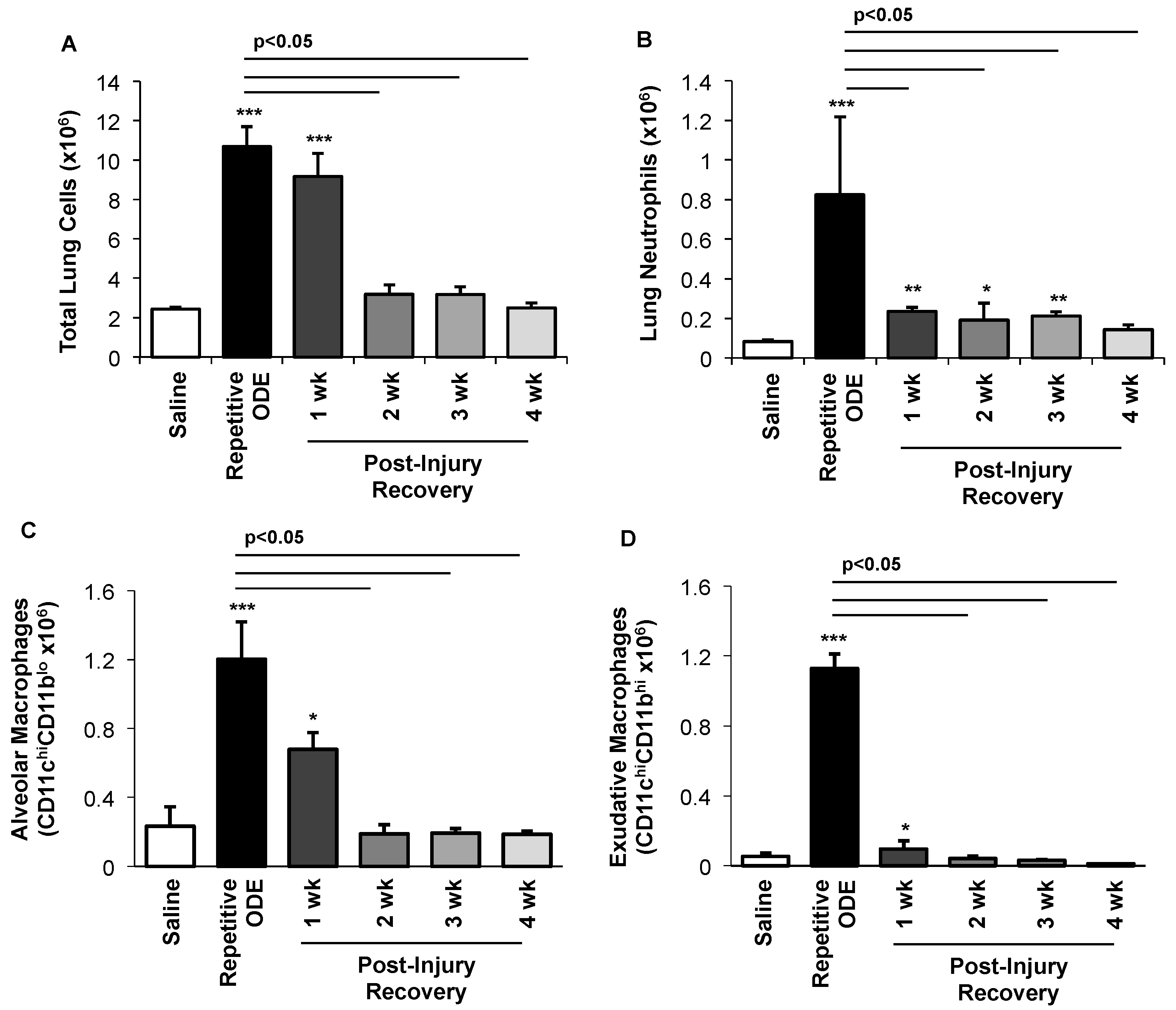
3.3. Clearance of Repetitive ODE Exposure-Induced Lung Neutrophils, Alveolar Macrophages, and Exudative Macrophages Is Time-Dependent
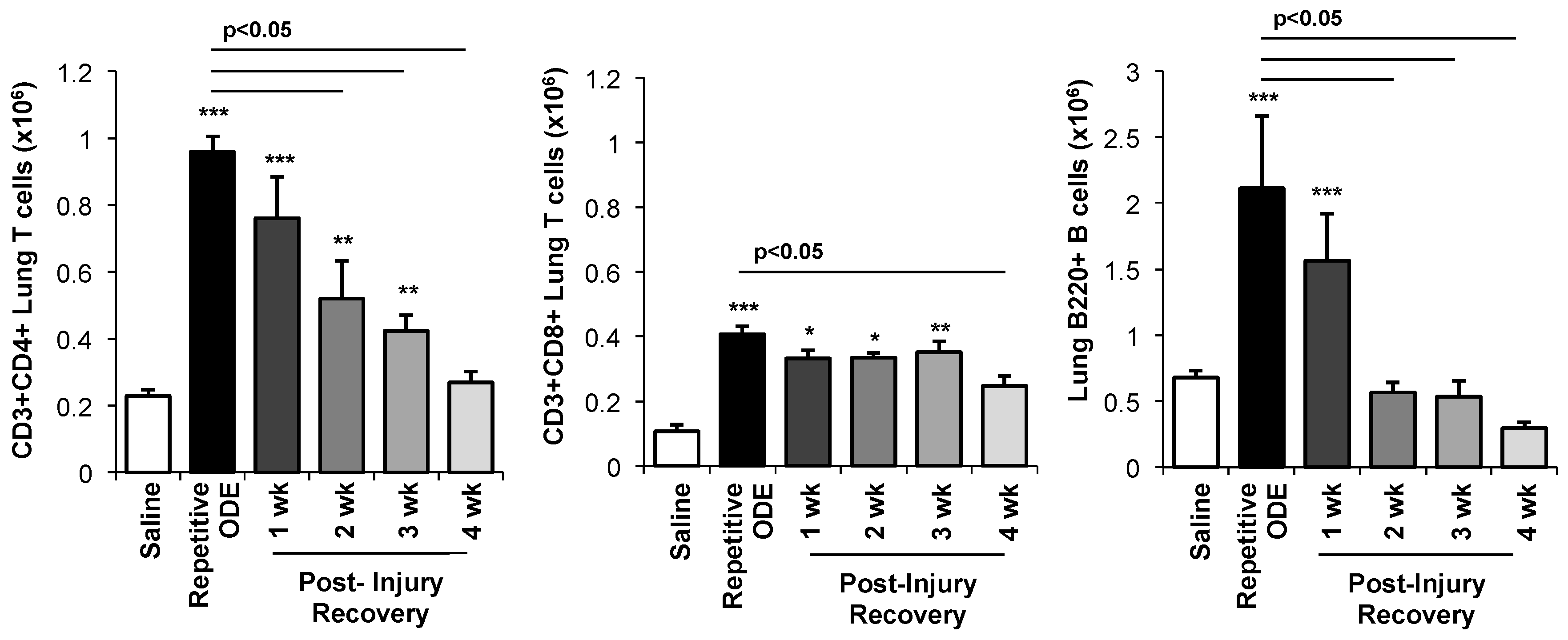
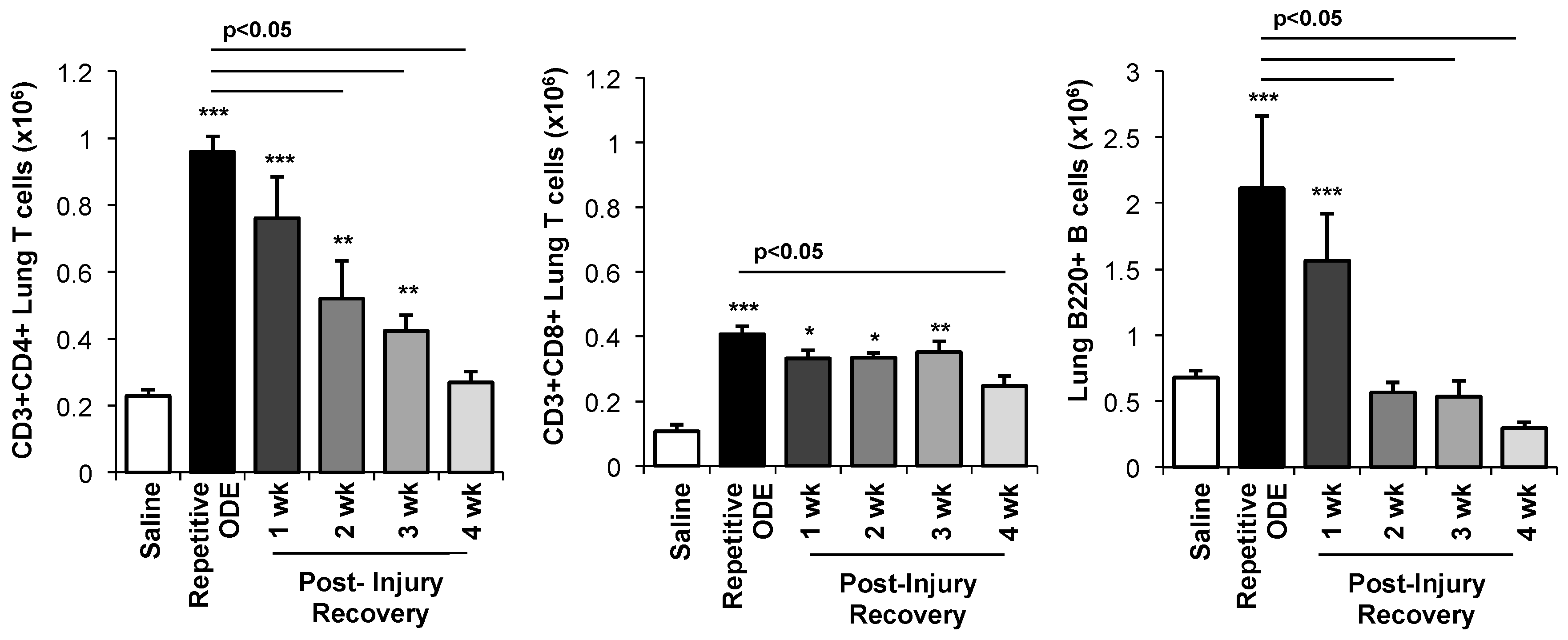
3.4. Lymphocytes, Particularly CD4+ and CD8+ T Cells, Demonstrated Prolonged Persistence in the Lung Following Final ODE Exposure
3.5. Apoptotic Cells within Cellular Aggregates Diminished over Time Following Final Repetitive ODE Exposure
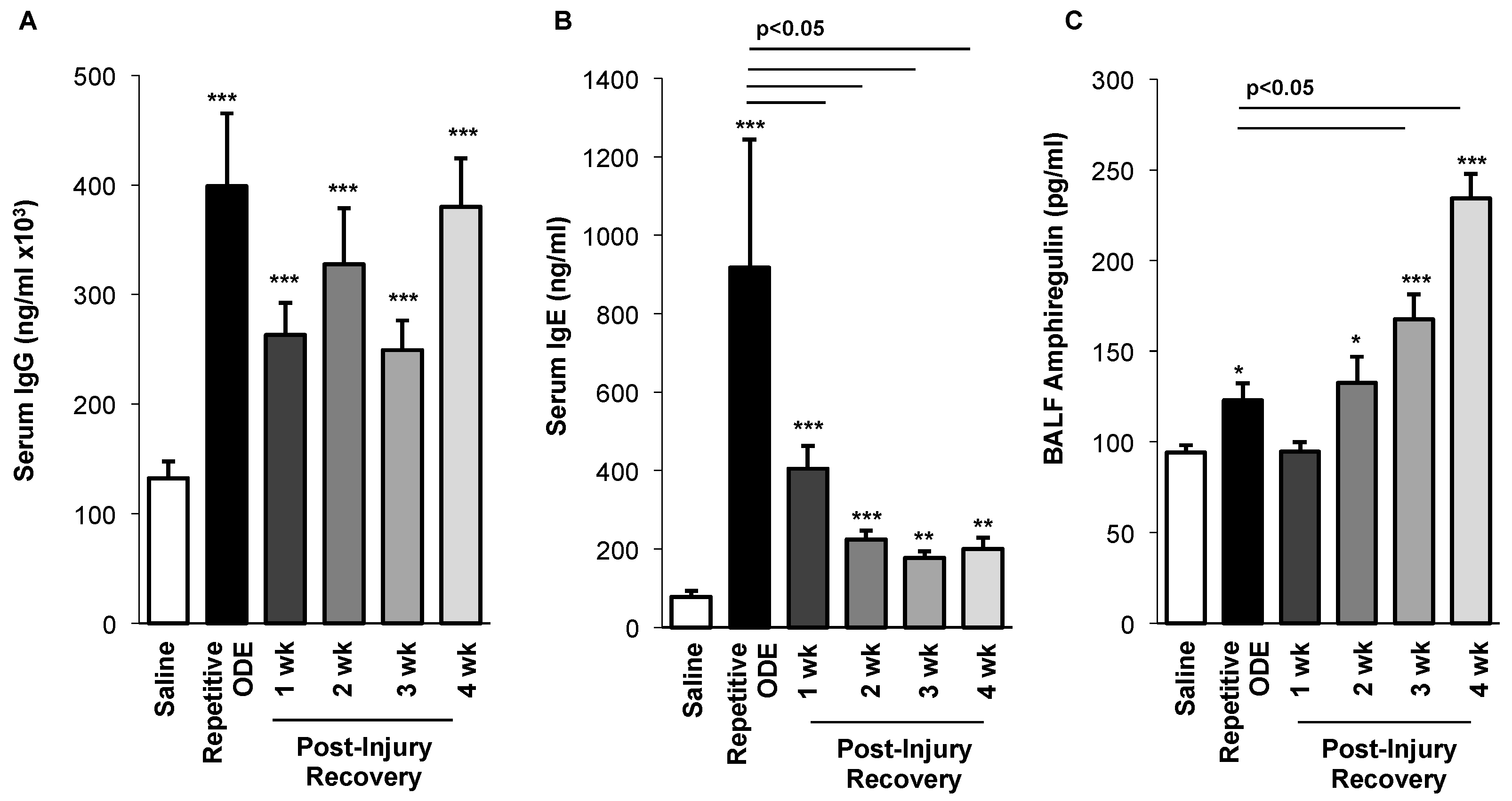
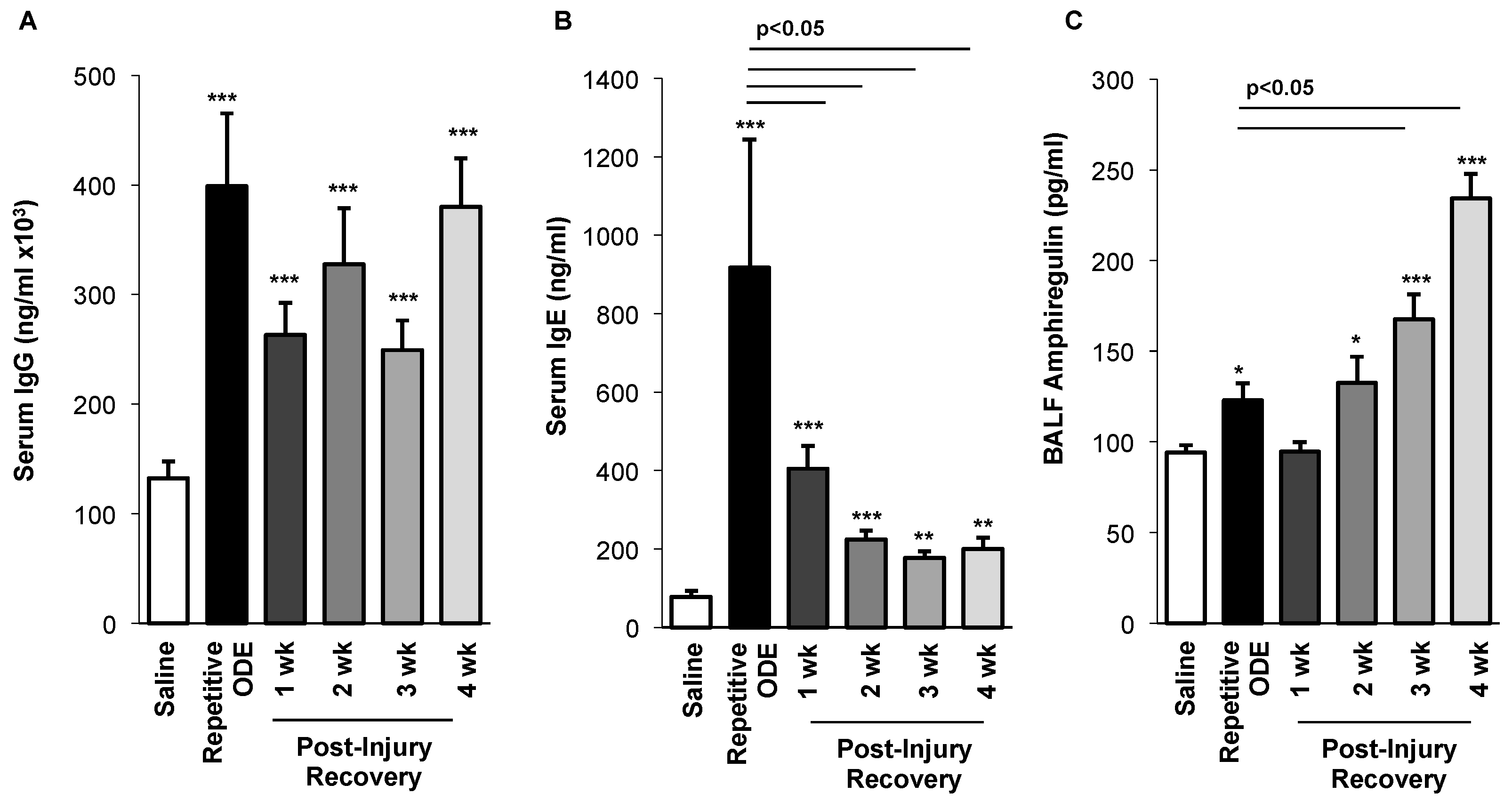
3.6. Levels of Serum Immunoglobulins Persisted and BALF Amphiregulin Levels Increased over Time Following Repetitive ODE Exposure
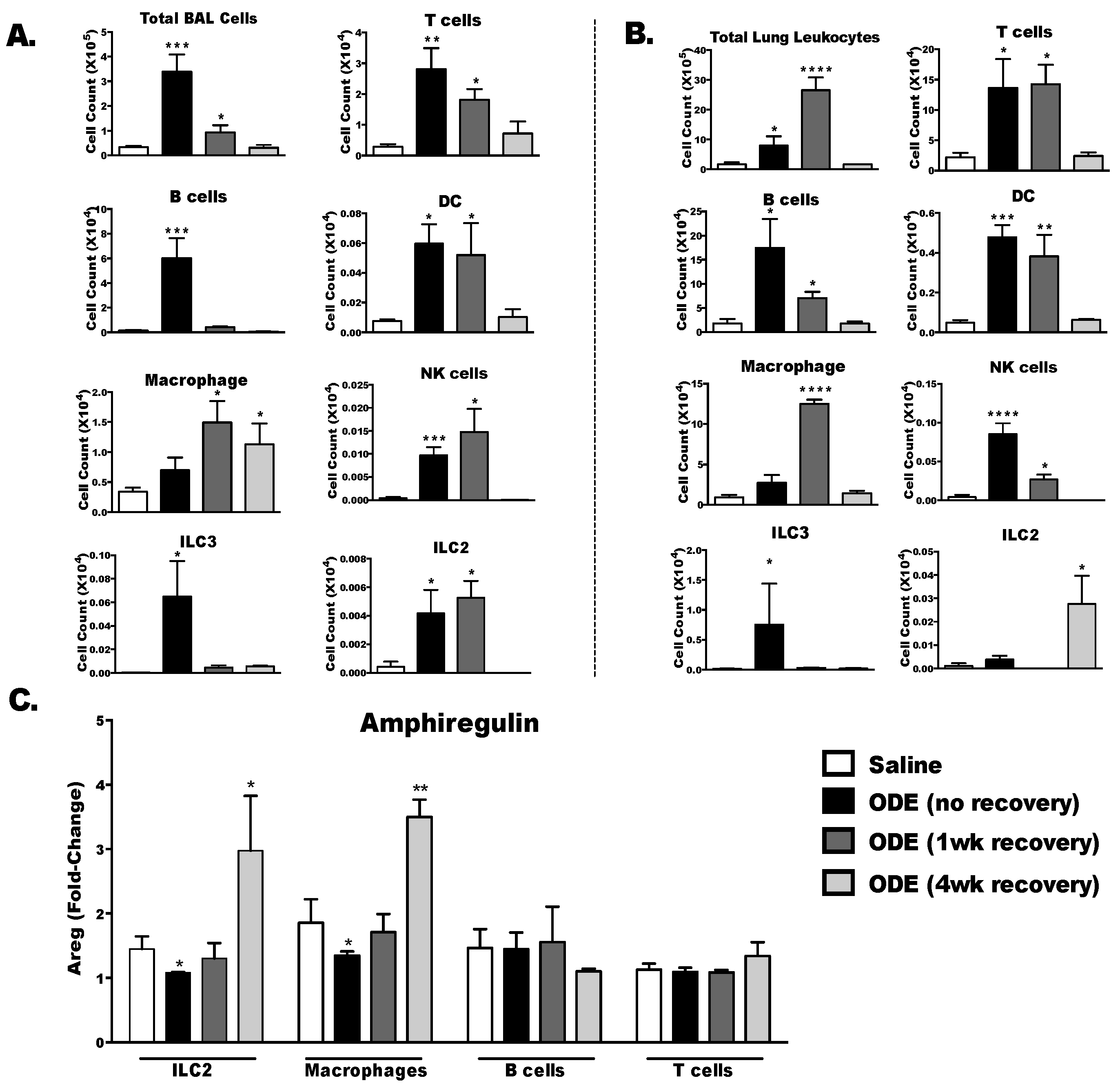
3.7. Amphiregulin is Up-Regulated in Type 2 Innate Lymphoid Cells and Macrophage Four Weeks Following the Final ODE Exposure
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Sundblad, B.M.; von Scheele, I.; Palmberg, L.; Olsson, M.; Larsson, K. Repeated exposure to organic material alters inflammatory and physiological airway responses. Eur. Respir. J. 2009, 34, 80–88. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, S.J.; Donham, K.J.; Whitten, P.; Merchant, J.A.; Burmeister, L.F.; Popendorf, W.J. Longitudinal evaluation of dose-response relationships for environmental exposures and pulmonary function in swine production workers. Am. J. Ind. Med. 1996, 29, 33–40. [Google Scholar] [CrossRef]
- Eduard, W.; Pearce, N.; Douwes, J. Chronic bronchitis, COPD, and lung function farmers: The role of biological agents. Chest 2009, 136, 716–725. [Google Scholar] [CrossRef] [PubMed]
- Eduard, W.; Douwes, J.; Omenaas, E.; Heederik, D. Do farming exposures cause or prevent asthma? Results from a study of adult Norwegian farmers. Thorax 2004, 59, 381–386. [Google Scholar] [CrossRef] [PubMed]
- Douwes, J.; Gibson, P.; Pekkanen, J.; Pearce, N. Non-eosinophilic asthma: Importance and possible mechanisms. Thorax 2002, 57, 643–648. [Google Scholar] [CrossRef] [PubMed]
- Poole, J.A.; Wyatt, T.A.; Oldenburg, P.J.; Elliott, M.K.; West, W.W.; Sisson, J.H.; Von Essen, S.G.; Romberger, D.J. Intranasal organic dust exposure-induced airway adaptation response marked by persistent lung inflammation and pathology in mice. Am. J. Physiol. Lung Cell. Mol. Physiol. 2009, 296, L1085–L1095. [Google Scholar] [CrossRef] [PubMed]
- Romberger, D.J.; Heires, A.J.; Nordgren, T.M.; Souder, C.P.; West, W.; Liu, X.D.; Poole, J.A.; Toews, M.L.; Wyatt, T.A. Proteases in agricultural dust induce lung inflammation through PAR-1 and PAR-2 activation. Am. J. Physiol. Lung Cell. Mol. Physiol. 2015, 309, L388–L399. [Google Scholar] [CrossRef] [PubMed]
- Boissy, R.J.; Romberger, D.J.; Roughead, W.A.; Weissenburger-Moser, L.; Poole, J.A.; LeVan, T.D. Shotgun pyrosequencing metagenomic analyses of dusts from swine confinement and grain facilities. PLoS ONE 2014, 9, e95578. [Google Scholar] [CrossRef] [PubMed]
- Amigoni, M.; Bellani, G.; Scanziani, M.; Masson, S.; Bertoli, E.; Radaelli, E.; Patroniti, N.; Di Lelio, A.; Pesenti, A.; Latini, R. Lung injury and recovery in a murine model of unilateral acid aspiration: Functional, biochemical, and morphologic characterization. Anesthesiology 2008, 108, 1037–1046. [Google Scholar] [CrossRef] [PubMed]
- Bauer, C.; Kielian, T.; Wyatt, T.A.; Romberger, D.J.; West, W.W.; Romberger, D.J. Myeloid differentiation factor 88-dependent signaling is critical for acute organic dust-induced airway inflammation in mice. Am. J. Respir. Cell Mol. Biol. 2013, 48, 781–789. [Google Scholar] [CrossRef] [PubMed]
- Poole, J.A.; Wyatt, T.A.; Kielian, T.; Oldenburg, P.; Gleason, A.M.; Bauer, A.; Golden, G.; West, W.W.; Sisson, J.H.; Romberger, D.J. Toll-like receptor 2 regulates organic dust-induced airway inflammation. Am. J. Respir. Cell Mol. Biol. 2011, 45, 711–719. [Google Scholar] [CrossRef] [PubMed]
- Poole, J.A.; Anderson, L.; Gleason, A.M.; West, W.W.; Romberger, D.J.; Wyatt, T.A. Pattern recognition scavenger receptor A/CD204 regulates airway inflammatory homeostasis following organic dust extract exposures. J. Immunotoxicol. 2015, 12, 64–73. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Charavaryamath, C.; Juneau, V.; Suri, S.S.; Janardhan, K.S.; Townsend, H.; Singh, B. Role of Toll-like receptor 4 in lung inflammation following exposure to swine barn air. Exp. Lung Res. 2008, 34, 19–35. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.; Dosman, J.A.; Rennie, D.C.; Schwartz, D.A.; Yang, I.V.; Beach, J.; Senthiselvan, A. Association of Toll-like receptor 2 gene polymorphisms with lung function in workers in swine operations. Ann. Allergy Asthma Immunol. 2013, 110, 44–50 e41. [Google Scholar] [CrossRef] [PubMed]
- Senthilselvan, A.; Dosman, J.A.; Chenard, L.; Burch, L.H.; Predicala, B.Z.; Sorowski, R.; Schneberger, D.; Hurst, T.; Kirychuk, S.; Gerdts, V.; et al. Toll-like receptor 4 variants reduce airway response in human subjects at high endotoxin levels in a swine facility. J. Allergy Clin. Immunol. 2009, 123, 1034–1040. [Google Scholar] [CrossRef] [PubMed]
- Monso, E.; Riu, E.; Radon, K.; Magarolas, R.; Danuser, B.; Iversen, M.; Morera, J.; Nowak, D. Chronic obstructive pulmonary disease in never-smoking animal farmers working inside confinement buildings. Am. J. Ind. Med. 2004, 46, 357–362. [Google Scholar] [CrossRef] [PubMed]
- Dakhama, A.; Israel-Assayag, E.; Cormier, Y. Role of interleukin-2 in the development and persistence of lymphocytic alveolitis in farmer’s lung. Eur. Respir. J. 1998, 11, 1281–1286. [Google Scholar] [CrossRef] [PubMed]
- Bailey, K.L.; Meza, J.L.; Smith, L.M.; Von Essen, S.G.; Romberger, D.J. Agricultural exposures in patients with COPD in health systems serving rural areas. J. Agromed. 2007, 12, 71–76. [Google Scholar] [CrossRef]
- Szczyrek, M.; Krawczyk, P.; Milanowski, J.; Jastrzebska, I.; Zwolak, A.; Daniluk, J. Chronic obstructive pulmonary disease in farmers and agricultural workers—An overview. Ann. Agric. Environ. Med. 2011, 18, 310–313. [Google Scholar] [PubMed]
- Fernandez, I.E.; Amarie, O.V.; Mutze, K.; Konigshoff, M.; Yildirim, A.O.; Eickelberg, O. Systematic phenotyping and correlation of biomarkers with lung function and histology in lung fibrosis. Am. J. Physiol. Lung Cell. Mol. Physiol. 2016, 310, L919–L927. [Google Scholar] [CrossRef] [PubMed]
- Robb, C.T.; Regan, K.H.; Dorward, D.A.; Rossi, A.G. Key mechanisms governing resolution of lung inflammation. Semin. Immunopathol. 2016, 38, 425–448. [Google Scholar] [CrossRef] [PubMed]
- Poole, J.A.; Gleason, A.M.; Bauer, C.; West, W.W.; Alexis, N.; van Rooijen, N.; Reynolds, S.J.; Romberger, D.J.; Kielian, T.L. CD11c(+)/CD11b(+) cells are critical for organic dust-elicited murine lung inflammation. Am. J. Respir. Cell Mol. Biol. 2012, 47, 652–659. [Google Scholar] [CrossRef] [PubMed]
- Poole, J.A.; Gleason, A.M.; Bauer, C.; West, W.W.; Alexis, N.; Reynolds, S.J.; Romberger, D.J.; Kielian, T. alphabeta T cells and a mixed Th1/Th17 response are important in organic dust-induced airway disease. Ann. Allergy Asthma Immunol. 2012, 109, 266–273. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Krishnamoorthy, N.; Burkett, P.R.; Dalli, J.; Abdulnour, R.E.; Colas, R.; Rampn, S.; Phipps, R.P.; Petasis, N.A.; Kuchroo, V.K.; Serhan, C.N.; et al. Cutting edge: Maresin-1 engages regulatory T cells to limit type 2 innate lymphoid cell activation and promote resolution of lung inflammation. J. Immunol. 2015, 194, 863–867. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.Y.; Umetsu, D.T.; Dekruyff, R.H. Innate lymphoid cells in asthma: Will they take your breath away? Eur. J. Immunol. 2016, 46, 795–806. [Google Scholar] [CrossRef] [PubMed]
- Zaiss, D.M.; Gause, W.C.; Osborne, L.C.; Artis, D. Emerging functions of amphiregulin in orchestrating immunity, inflammation, and tissue repair. Immunity 2015, 42, 216–226. [Google Scholar] [CrossRef] [PubMed]
- Monticelli, L.A.; Sonnenberg, G.F.; Abt, M.C.; Alenghat, T.; Ziegler, C.G.; Doering, T.A.; Angelosanto, J.M.; Laidlaw, B.J.; Yank, C.Y.; Sathaliyawala, T.; et al. Innate lymphoid cells promote lung-tissue homeostasis after infection with influenza virus. Nat. Immunol. 2011, 12, 1045–1054. [Google Scholar] [CrossRef] [PubMed]
- Colgan, S.P. Neutrophils and inflammatory resolution in the mucosa. Semin. Immunol. 2015, 27, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Sharma, A. Neutrophils: Cinderella of innate immune system. Int. Immunopharmacol. 2010, 10, 1325–1334. [Google Scholar] [CrossRef] [PubMed]
- Jie, H.; He, Y.; Huang, X.; Zhou, Q.; Han, Y.; Liu, X.; Bai, Y.; Sun, E. Necrostatin-1 enhances the resolution of inflammation by specifically inducing neutrophil apoptosis. Oncotarget 2016, 12, 19367–19381. [Google Scholar] [CrossRef] [PubMed]
- Kawano, H.; Kayama, H.; Nakama, T.; Hashimoto, T.; Umemoto, E.; Takeda, K. IL-10-producing lung interstitial macrophages prevent neutrophilic asthma. Int. Immunol. 2016, 28, 489–501. [Google Scholar] [CrossRef] [PubMed]
- Campbell, E.L.; Colgan, S.P. Neutrophils and inflammatory metabolism in antimicrobial functions of the mucosa. J. Leukoc. Biol. 2015, 98, 517–522. [Google Scholar] [CrossRef] [PubMed]
- Das, A.; Sinha, M.; Datta, S.; Abas, M.; Chaffee, S.; Sen, C.K.; Roy, S. Monocyte and macrophage plasticity in tissue repair and regeneration. Am. J. Pathol. 2015, 185, 2596–2606. [Google Scholar] [CrossRef] [PubMed]
- Mosser, D.M.; Edwards, J.P. Exploring the full spectrum of macrophage activation. Nat. Rev. Immunol. 2008, 8, 958–969. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, J.; Westphalen, K. Macrophage-epithelial interactions in pulmonary alveoli. Semin. Immunopathol. 2016, 38, 461–469. [Google Scholar] [CrossRef] [PubMed]
- Lai, J.F.; Zindl, C.L.; Duffy, L.B.; Atkinson, T.P.; Jung, Y.W.; van Rooijen, N.; Waites, K.B.; Krause, D.C.; Chaplin, D.D. Critical role of macrophages and their activation via MyD88-NFkappaB signaling in lung innate immunity to Mycoplasma pneumoniae. PLoS ONE 2010, 5, e14417. [Google Scholar] [CrossRef] [PubMed]
- Robbe, P.; Draijer, C.; Borg, T.R.; Luinge, M.; Timens, W.; Wouters, I.M.; Melgert, B.N.; Hylkema, M.N. Distinct macrophage phenotypes in allergic and nonallergic lung inflammation. Am. J. Physiol. Lung Cell. Mol. Physiol. 2015, 308, L358–L367. [Google Scholar] [CrossRef] [PubMed]
- Croasdell, A.; Duffney, P.F.; Kim, N.; Lacy, S.H.; Sime, P.J.; Phipps, R.P. PPARgamma and the Innate Immune System Mediate the Resolution of Inflammation. PPAR Res. 2015, 2015, 549691. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Meng, C.; Liu, G.; Yang, D.; Fu, L.; Zhang, M.; Zhang, Z.; Xia, H.; Yao, S.; Zhang, S. Classically activated macrophages protect against lipopolysaccharide-induced acute lung injury by expressing amphiregulin in mice. Anesthesiology 2016, 124, 1086–1099. [Google Scholar] [CrossRef] [PubMed]
- Randall, T.D. Bronchus-associated lymphoid tissue (BALT) structure and function. Adv. Immunol. 2010, 107, 187–241. [Google Scholar] [PubMed]
- Foo, S.Y.; Phipps, S. Regulation of inducible BALT formation and contribution to immunity and pathology. Mucosal Immunol. 2010, 3, 537–544. [Google Scholar] [CrossRef] [PubMed]
- Kudo, D.; Toyama, M.; Aoyagi, T.; Akahori, Y.; Yamamoto, H.; Ishii, K.; Kanno, E.; Maruyama, R.; Kaku, M.; Kushimoto, S.; et al. Involvement of high mobility group box 1 and the therapeutic effect of recombinant thrombomodulin in a mouse model of severe acute respiratory distress syndrome. Clin. Exp. Immunol. 2013, 173, 276–287. [Google Scholar] [CrossRef] [PubMed]
- Mock, J.R.; Garibaldi, B.T.; Aggarwal, N.R.; Jenkins, J.; Limjunyawong, N.; Singer, B.D.; Chau, E.; Rabold, R.; Files, D.C.; Sidhaye, V.; et al. Foxp3+ regulatory T cells promote lung epithelial proliferation. Mucosal Immunol. 2014, 7, 1440–1451. [Google Scholar] [CrossRef] [PubMed]
- Garibaldi, B.T.; D’Alessio, F.R.; Mock, J.R.; Files, D.C.; Chau, E.; Eto, Y.; Drummond, M.B.; Aggarwal, N.R.; Sidhaye, V.; King, L.S. Regulatory T cells reduce acute lung injury fibroproliferation by decreasing fibrocyte recruitment. Am. J. Respir. Cell Mol. Biol. 2013, 48, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Erkinjuntti-Pekkanen, R.; Reiman, M.; Kokkarinen, J.I.; Tukiainen, H.O.; Terho, E.O. IgG antibodies, chronic bronchitis, and pulmonary function values in farmer’s lung patients and matched controls. Allergy 1999, 54, 1181–1187. [Google Scholar] [CrossRef] [PubMed]
- Kaukonen, K.; Savolainen, J.; Nermes, M.; Viander, M.; Terho, E.O. IgE antibody response against Aspergillus umbrosus in farmer’s lung disease. Int. Arch. Allergy Immunol. 1997, 112, 313–316. [Google Scholar] [CrossRef] [PubMed]
- Manzo, N.D.; Foster, W.M.; Stripp, B.R. Amphiregulin-dependent mucous cell metaplasia in a model of nonallergic lung injury. Am. J. Respir. Cell Mol. Biol. 2012, 47, 349–357. [Google Scholar] [CrossRef] [PubMed]
- Kelly, F.L.; Sun, J.; Fischer, B.M.; Voynow, J.A.; Kummarapurugu, A.B.; Zhang, H.L.; Nugent, J.L.; Beasley, R.F.; Martinu, T.; Gwinn, W.M.; et al. Diacetyl induces amphiregulin shedding in pulmonary epithelial cells and in experimental bronchiolitis obliterans. Am. J. Respir. Cell Mol. Biol. 2014, 51, 568–574. [Google Scholar] [CrossRef] [PubMed]
- Madsen, A.M.; Saber, A.T.; Nordly, P.; Sharma, A.K.; Wallin, H.; Vogel, U. Inflammation but no DNA (deoxyribonucleic acid) damage in mice exposed to airborne dust from a biofuel plant. Scand. J. Work Environ. Health 2008, 34, 278–287. [Google Scholar] [CrossRef] [PubMed]
- Lemieszek, M.; Chilosi, M.; Golec, M.; Skorska, C.; Huaux, F.; Yakoub, Y.; Pastena, C.; Daniele, I.; Cholewa, G.; et al. Mouse model of hypersensitivity pneumonitis after inhalation exposure to different microbial antigens associated with organic dusts. Ann. Agric. Environ. Med. 2011, 18, 159–168. [Google Scholar] [PubMed]
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Warren, K.J.; Wyatt, T.A.; Romberger, D.J.; Ailts, I.; West, W.W.; Nelson, A.J.; Nordgren, T.M.; Staab, E.; Heires, A.J.; Poole, J.A. Post-Injury and Resolution Response to Repetitive Inhalation Exposure to Agricultural Organic Dust in Mice. Safety 2017, 3, 10. https://doi.org/10.3390/safety3010010
Warren KJ, Wyatt TA, Romberger DJ, Ailts I, West WW, Nelson AJ, Nordgren TM, Staab E, Heires AJ, Poole JA. Post-Injury and Resolution Response to Repetitive Inhalation Exposure to Agricultural Organic Dust in Mice. Safety. 2017; 3(1):10. https://doi.org/10.3390/safety3010010
Chicago/Turabian StyleWarren, Kristi J., Todd A. Wyatt, Debra J. Romberger, Isaak Ailts, William W. West, Amy J. Nelson, Tara M. Nordgren, Elizabeth Staab, Art J. Heires, and Jill A. Poole. 2017. "Post-Injury and Resolution Response to Repetitive Inhalation Exposure to Agricultural Organic Dust in Mice" Safety 3, no. 1: 10. https://doi.org/10.3390/safety3010010
APA StyleWarren, K. J., Wyatt, T. A., Romberger, D. J., Ailts, I., West, W. W., Nelson, A. J., Nordgren, T. M., Staab, E., Heires, A. J., & Poole, J. A. (2017). Post-Injury and Resolution Response to Repetitive Inhalation Exposure to Agricultural Organic Dust in Mice. Safety, 3(1), 10. https://doi.org/10.3390/safety3010010