
3D Clumped Cell Segmentation Using Curvature Based Seeded Watershed

Abstract
:1. Introduction
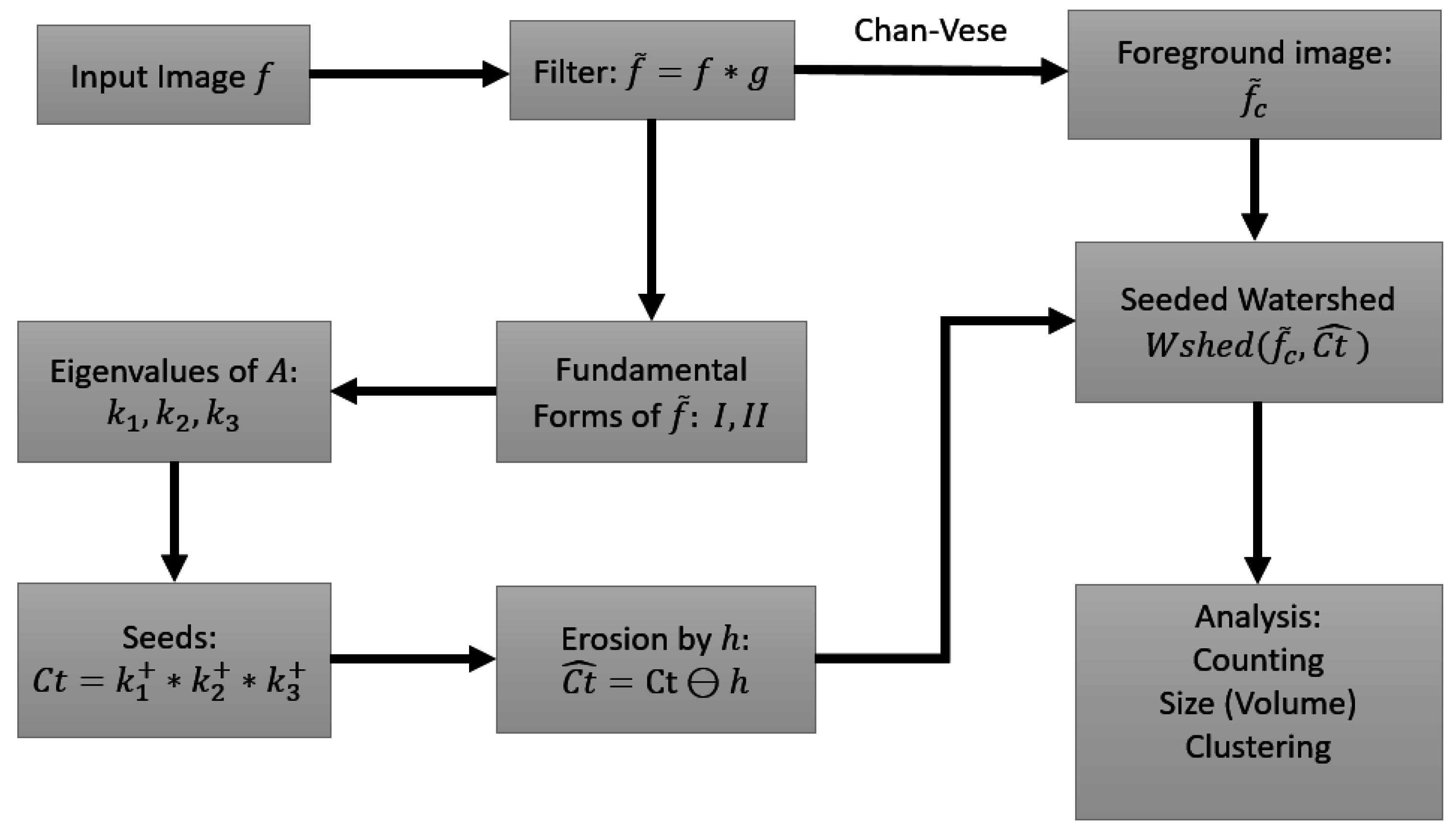
2. The Watershed Transform
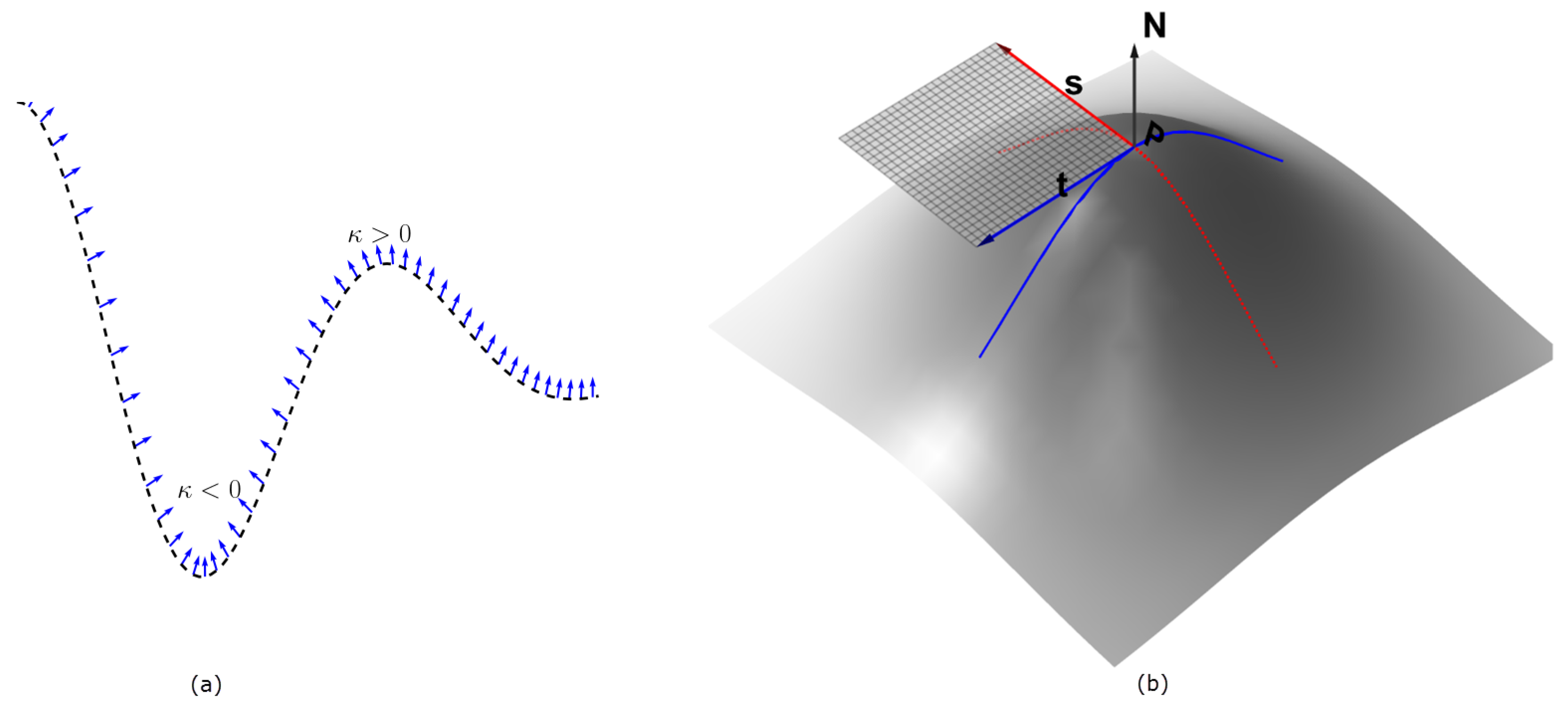
3. The Shape of the Manifold
3.1. The First and Second Fundamental Forms
3.2. Curvature of Implicit Surfaces
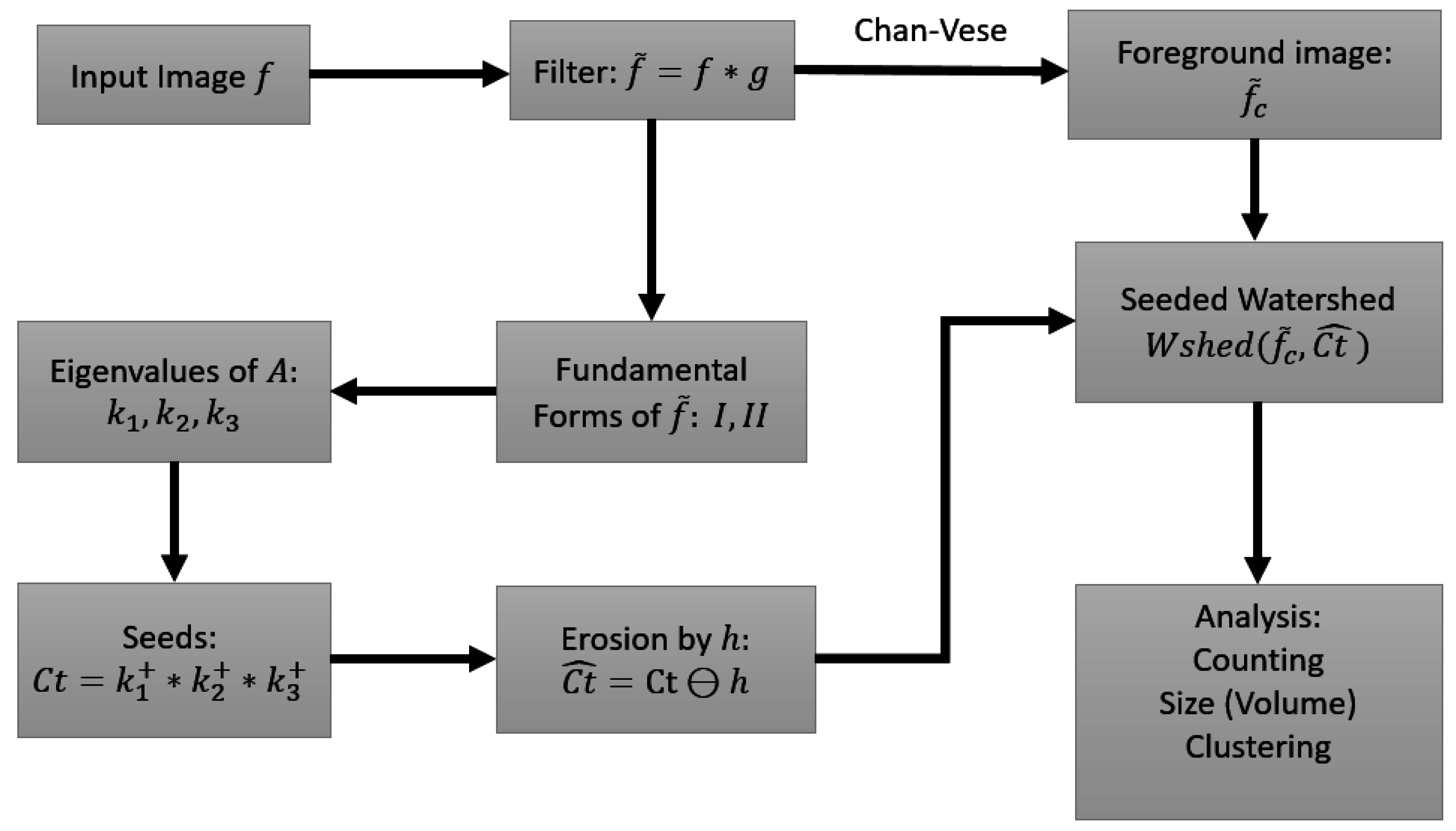
4. Materials and Methods
4.1. Dataset
4.2. Evaluation Methodology
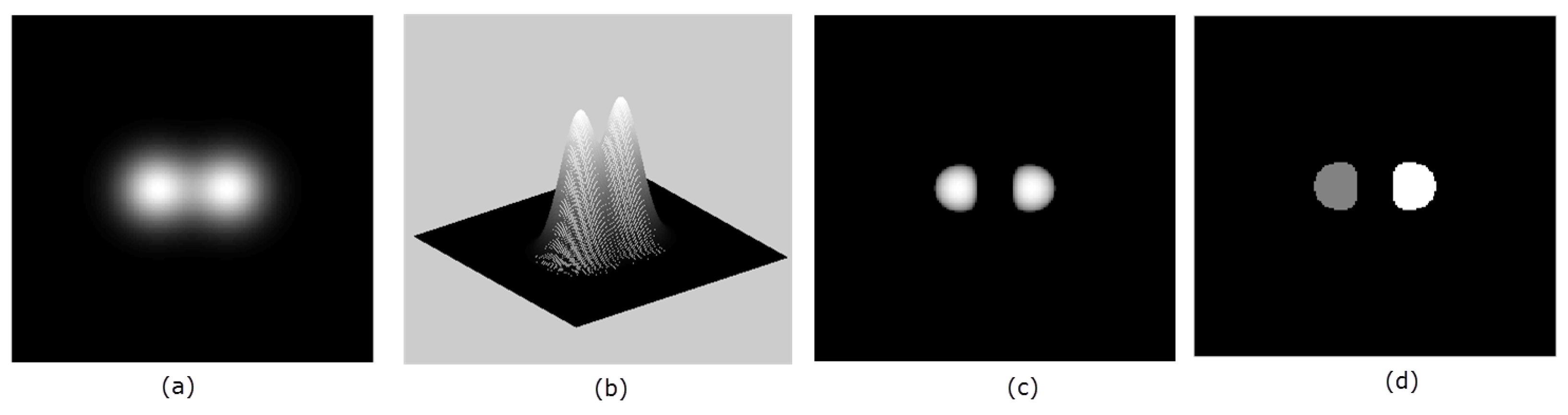
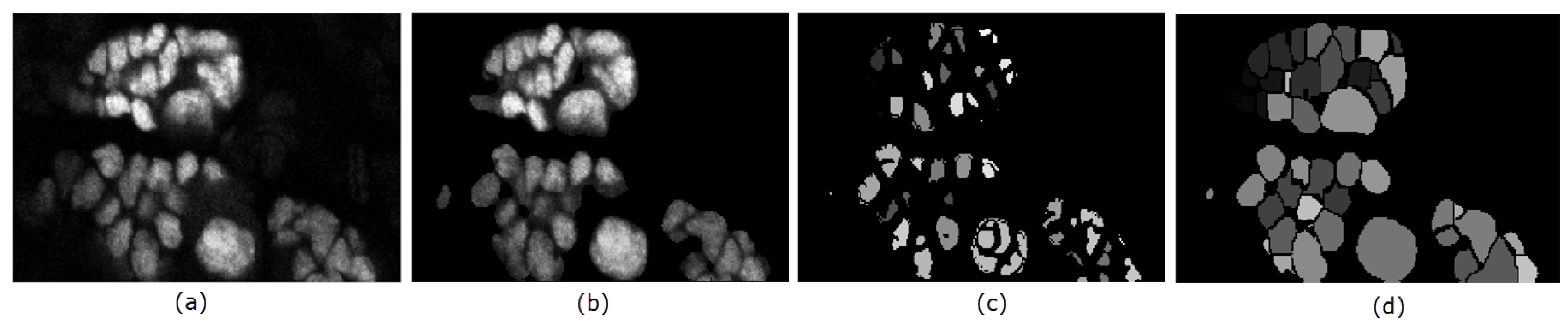
4.3. Data Pre-processing
4.4. Seed Detection and Watershed
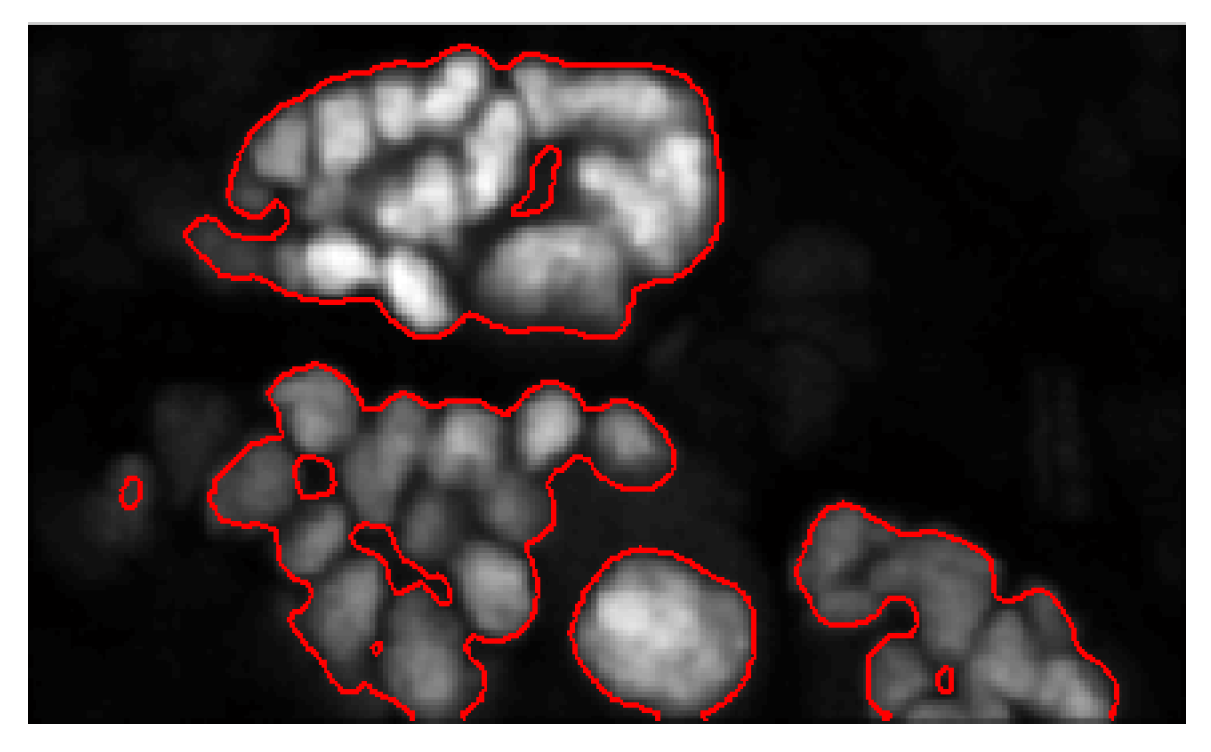
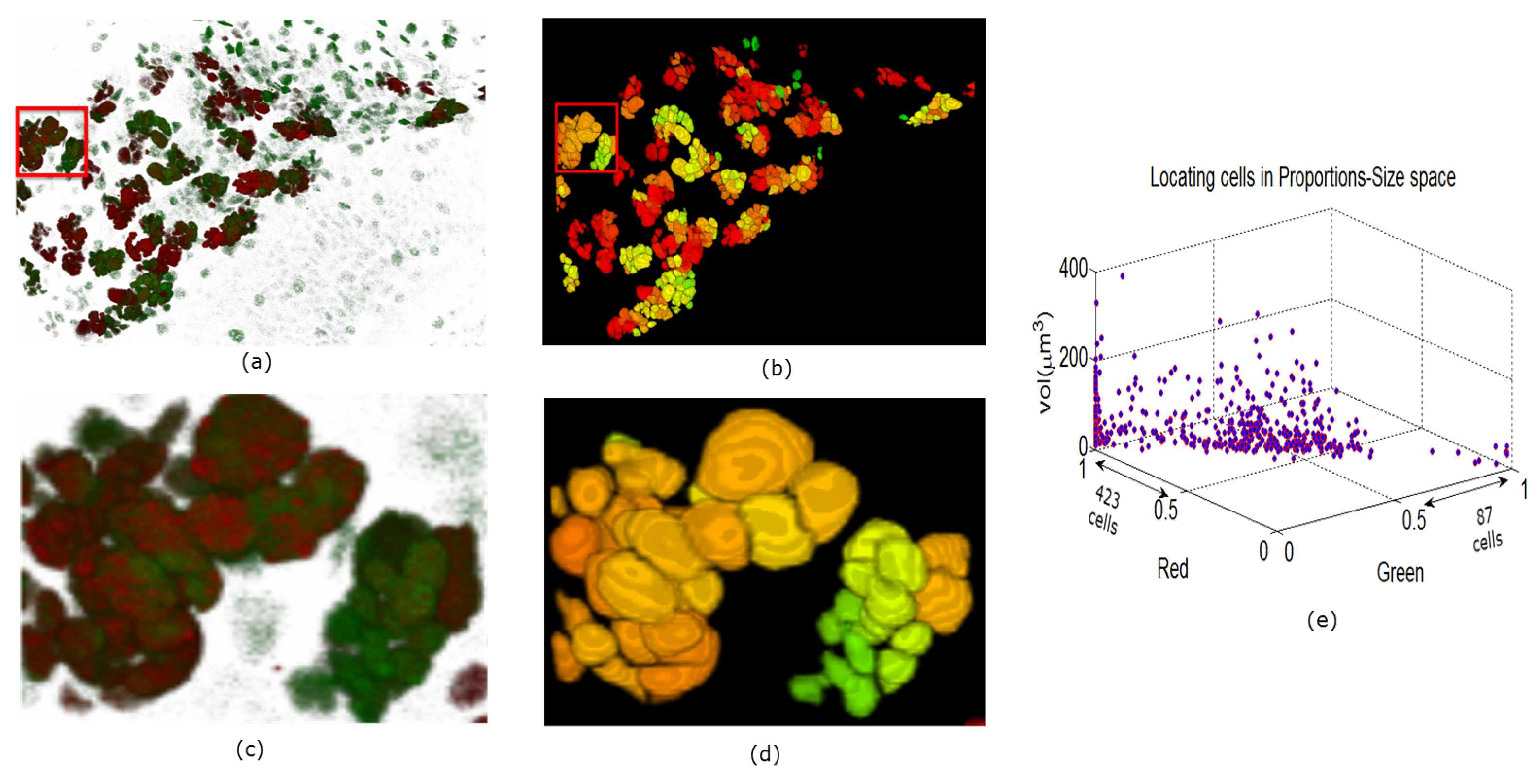
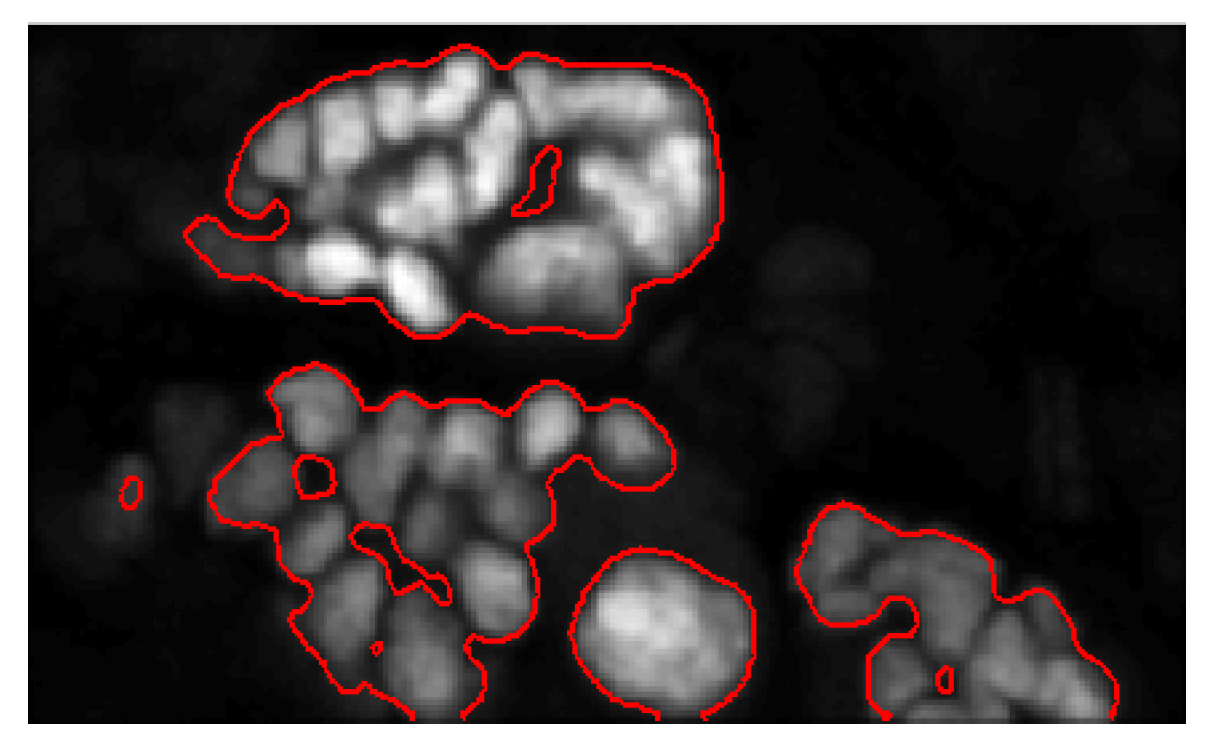
5. Results and Discussion
5.1. Evaluation on Sample Dataset
5.2. Computational Issues and Discussions
6. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Guan, B.X.; Bhanu, B.; Thakoor, N.; Talbot, P.; Lin, S. Automatic cell region detection by k-means with weighted entropy. In Proceedings of the 2013 IEEE 10th International Symposium on Biomedical Imaging (ISBI), San Francisco, CA, USA, 7–11 April 2013; pp. 418–421.
- Pal, S.K.; Ghosh, A.; Shankar, B.U. Segmentation of remotely sensed images with fuzzy thresholding, and quantitative evaluation. Int. J. Remote Sens. 2000, 21, 2269–2300. [Google Scholar] [CrossRef]
- Chan, T.F.; Vese, L.A. Active Contours without Edge. IEEE Trans. Image Process. 2001, 10, 266–277. [Google Scholar] [CrossRef] [PubMed]
- Zimmer, C.; Labruyere, E.; Meas-Yedid, V.; Guillen, N.; Olivo-Marin, J.C. Improving Active Contours for Segmentation and Tracking of Motile Cells in Videomicroscopy. In Proceedings of the 16th International Conference on Pattern Recognition (ICPR), Quebec City, QC, Canada, 11–15 August 2002; pp. 286–289.
- Hodneland, E.; Kögel, T.; Frei, D.M.; Gerdes, H.H.; Lundervold, A. CellSegm—A MATLAB toolbox for high-throughput 3D cell segmentation. Source Code Biol. Med. 2013, 8, 16. [Google Scholar] [CrossRef] [PubMed]
- Lou, X.; Kang, M.; Xenopoulos, P.; Muñoz-Descalzo, S.; Hadjantonakis, A.K. A Rapid and Efficient 2D/3D Nuclear Segmentation Method for Analysis of Early Mouse Embryo and Stem Cell Image Data. Stem Cell Rep. 2014, 2, 382–397. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Rajapakse, J.C. Segmentation of Clustered Nuclei With Shape Markers and Marking Function. IEEE Trans. Biomed. Eng. 2009, 56, 741–748. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Sun, C.; Su, R.; Pham, T.D. Segmentation of clustered nuclei based on curvature weighting. In IVCNZ; McCane, B., Mills, S., Deng, J.D., Eds.; ACM: New York, NY, USA, 2012; pp. 49–54. [Google Scholar]
- Yang, X.; Li, H.; Zhou, X. Nuclei Segmentation Using Marker-Controlled Watershed, Tracking Using Mean-Shift, and Kalman Filter in Time-Lapse Microscopy. IEEE Trans. Circ. Syst. 2006, 53-I, 2405–2414. [Google Scholar] [CrossRef]
- Rodríguez, R.M.; Alarcón, T.E.; Pacheco, O. A new strategy to obtain robust markers for blood vessels segmentation by using the watersheds method. Comput. Biol. Med. 2005, 35, 665–686. [Google Scholar] [CrossRef] [PubMed]
- Roerdink, J.; Meijster, A. The Watershed Transform: Definitions, Algorithms and Parallization Strategies. Fundam. Inform. 2001, 41, 187–228. [Google Scholar]
- Beucher, S.; Meyer, F. The Morphological Approach to Segmentation: The Watershed Transformation. In Mathematical Morphology in Image Processing; CRC Press: Boca Raton, FL, USA, 1992; pp. 433–481. [Google Scholar]
- Beucher, S. Watershed, hierarchical segmentation and waterfall algorithm. Math. Morphol. Its Appl. Image Process. 1994, II, 69–76. [Google Scholar]
- Lin, G.; Adiga, U.; Olson, K.; Guzowski, J.; Barnes, C.; Roysam, B. A Hybrid 3D Watershed Algorithms Incorporating Gradient cues and Object Models for Automatic Segmentation of Nuclei in Confocal Image Stacks. Cytom. Part A 2003, 56, 23–36. [Google Scholar] [CrossRef] [PubMed]
- Wählby, C.; Bengtsson, E. Segmentation of Cell Nuclei in Tissue by Combining Seeded Watersheds with Gradient Information. In SCIA; Bigün, J., Gustavsson, T., Eds.; Springer: Berlin, Germany, 2003; Volume 2749, pp. 408–414. [Google Scholar]
- Deng, H.; Zhang, W.; Mortensen, E.N.; Dietterich, T.G.; Shapiro, L.G. Principal Curvature-Based Region Detector for Object Recognition. In Proceedings of the IEEE Conference on Computer Vision and Pattern Recognition, Minneapolis, MN, USA, 17–22 June 2007.
- Pulla, S.; Razdan, A.; Farin, G. Improved curvature estimation for watershed segmentation of 3-dimensional meshes. IEEE Trans. Vis. Comput. Graph. 2001, 5, 308–321. [Google Scholar]
- Digabel, H.; Lantuéjoul, C. Iterative Algorithms. In Proceedings of the European Symposium on Quantitative Analysis of Microstructures in Materials Sciences, Biology and Medicine, Caen, France, 4–7 October 1977; pp. 85–99.
- Beucher, S.; Lantuejoul, C. Use of Watersheds in Contour Detection. In Proceedings of the International Workshop on Image Processing: Real-time Edge and Motion Detection/Estimation, Rennes, France, 17–21 September 1979.
- Audigier, R.; Lotufo, R. Seed-Relative Segmentation Robustness of Watershed and Fuzzy Connectedness Approaches. IEEE Comput. Graph. Image Process. 2007, SIBGRAPI, 61–70. [Google Scholar]
- Meyer, F. Topographic distance and watershed lines. Signal Process. 1994, 38, 113–125. [Google Scholar] [CrossRef]
- Lee, M.J. Riemannian Manifolds: An Introduction to Curvature; Springer: Berlin, Germany, 1997. [Google Scholar]
- Do Carmo, M. Differential Geometry of Curves and Surfaces; Prentice-Ha lnc.: Englewood Cliffs, NJ, USA, 1976. [Google Scholar]
- Svoboda, D.; Kozubek, M.; Stejskal, S. Generation of Digital Phantoms of Cell Nuclei and Simulation of Image Formation in 3D Image Cytometry. Cytom. Part A 2009, 75A, 494–509. [Google Scholar] [CrossRef] [PubMed]
- Rand, M.W. Objective Criteria for the Evaluation of Clustering Methods. Am. Stat. Assoc. 1971, 66, 846–850. [Google Scholar] [CrossRef]
- Jaccard, P. The Distribution of the Flora in the Alpine Zone. New Phytol. 1912, 11, 37–50. [Google Scholar] [CrossRef]
- Van den Boomgaard, R.; van Balen, R. Methods for fast morphological image transforms using bitmapped binary images. CVGIP 1992, 54, 252–258. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
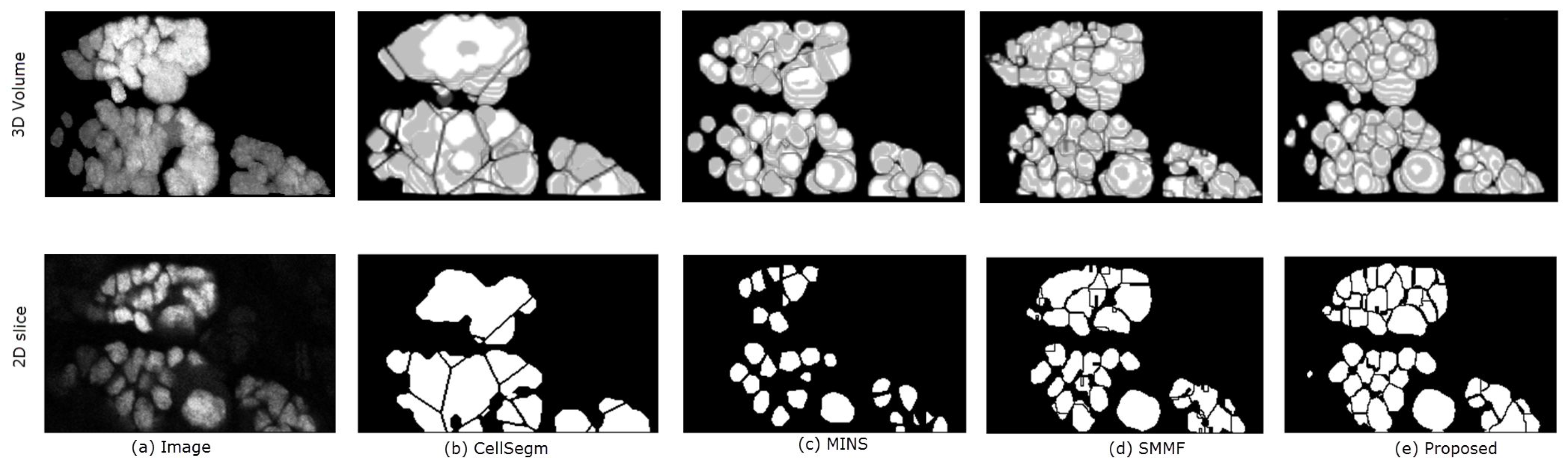{kind=link}
{kind=link}
{kind=link}
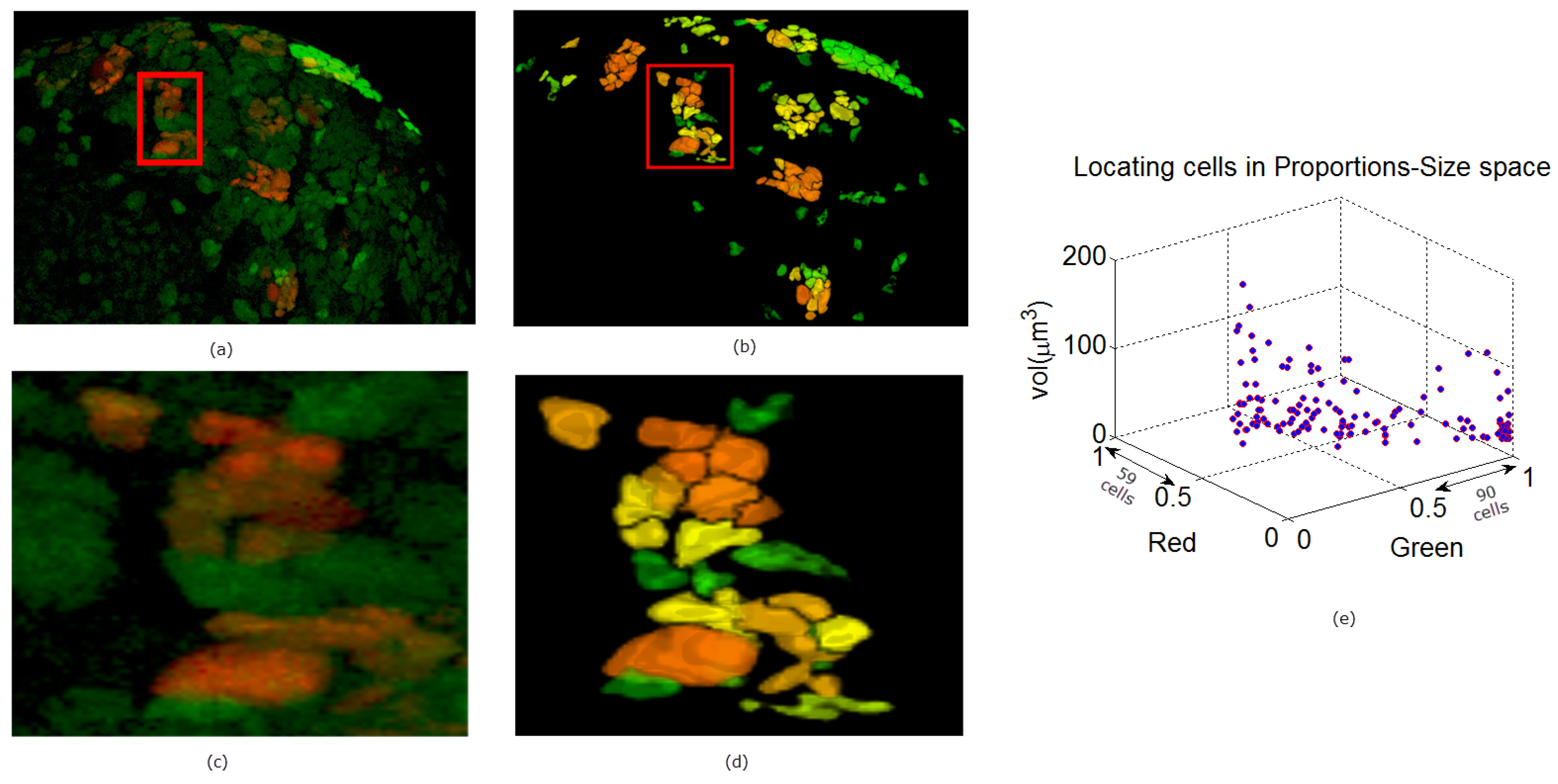{kind=link}
| Algorithm | RI | JI | Oversegmented | Undersegmented | Missing |
|---|---|---|---|---|---|
| CellSegm | 92.24% | 0.659 | 0.005 | 0.00 | 0.00 |
| MINS | 95.86% | 0.713 | 0.003 | 0.00 | 0.025 |
| SMMF | 96.57% | 0.804 | 0.00 | ||
| Proposed | 97.05% | 0.812 | 0.00 | 0.015 | 0.00 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Atta-Fosu, T.; Guo, W.; Jeter, D.; Mizutani, C.M.; Stopczynski, N.; Sousa-Neves, R. 3D Clumped Cell Segmentation Using Curvature Based Seeded Watershed. J. Imaging 2016, 2, 31. https://doi.org/10.3390/jimaging2040031
Atta-Fosu T, Guo W, Jeter D, Mizutani CM, Stopczynski N, Sousa-Neves R. 3D Clumped Cell Segmentation Using Curvature Based Seeded Watershed. Journal of Imaging. 2016; 2(4):31. https://doi.org/10.3390/jimaging2040031
Chicago/Turabian StyleAtta-Fosu, Thomas, Weihong Guo, Dana Jeter, Claudia M. Mizutani, Nathan Stopczynski, and Rui Sousa-Neves. 2016. "3D Clumped Cell Segmentation Using Curvature Based Seeded Watershed" Journal of Imaging 2, no. 4: 31. https://doi.org/10.3390/jimaging2040031
APA StyleAtta-Fosu, T., Guo, W., Jeter, D., Mizutani, C. M., Stopczynski, N., & Sousa-Neves, R. (2016). 3D Clumped Cell Segmentation Using Curvature Based Seeded Watershed. Journal of Imaging, 2(4), 31. https://doi.org/10.3390/jimaging2040031