

In Silico Apple Genome-Encoded MicroRNA Target Binding Sites Targeting Apple Chlorotic Leaf Spot Virus



Abstract
1. Introduction
2. Materials and Methods
2.1. Apple Mature MicroRNAs and ACLSV Genomic Data Source
2.2. Analysis of Multiple mdm-miRNA Target-Pairs in ACLSV Genome
2.3. miRanda
2.4. RNA22
2.5. TAPIR
2.6. psRNATarget
2.7. Discovering Apple mdm-miRNA-Target Interactions
2.8. RNAfold
2.9. RNAcofold
2.10. Statistical Analysis
3. Results
3.1. Apple Genome-Encoded mdm-miRNAs Targeting ACLSV Genome
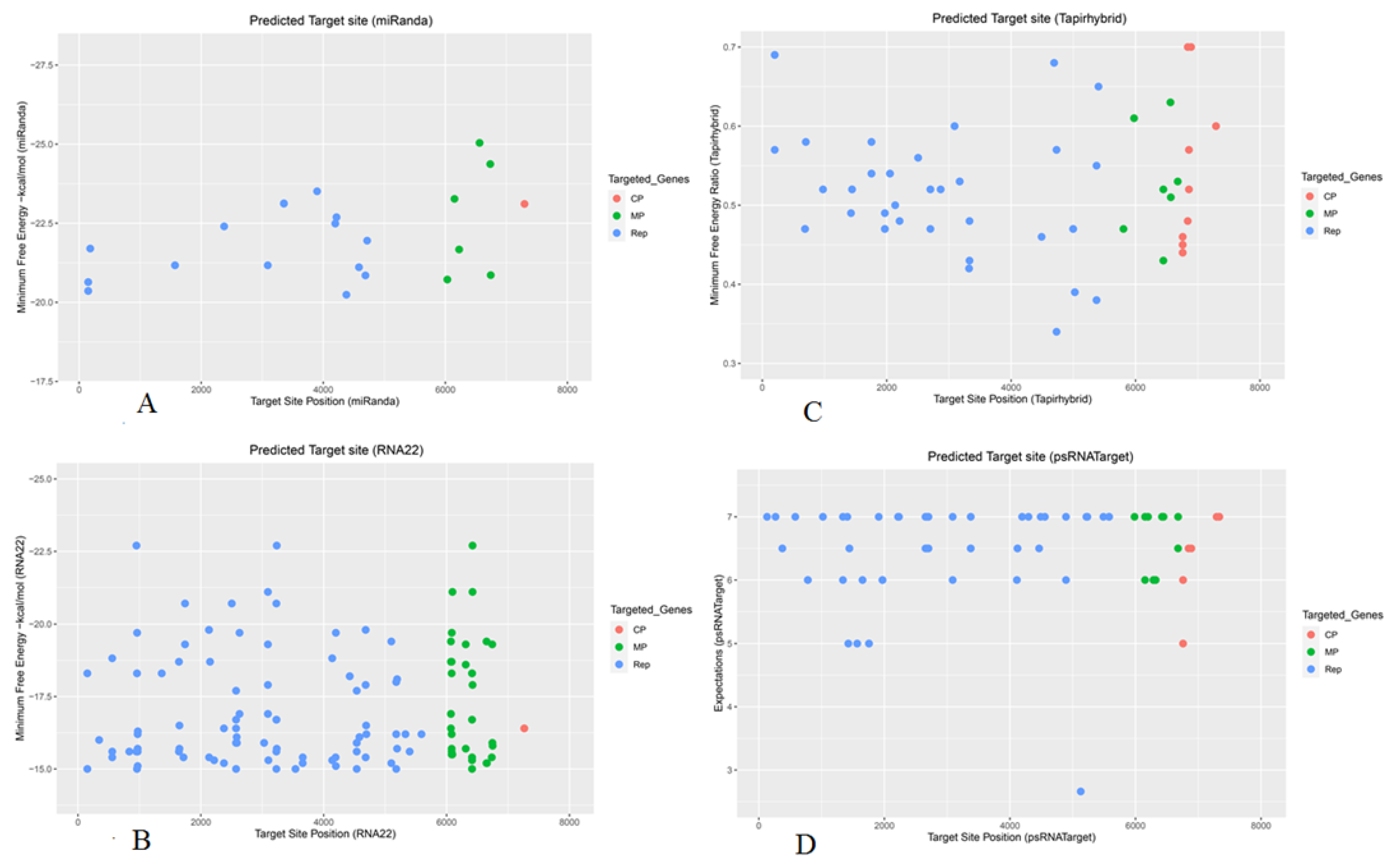
3.2. Apple mdm-miRNAs Targeting ORF1 That Ecodes Replication-Asscoiated Protein
3.3. Apple mdm-miRNAs Targeting ORF2 That Encodes Movement Protein
3.4. Apple mdm-miRNAs Targeting ORF3 That Encodes Coat Protein
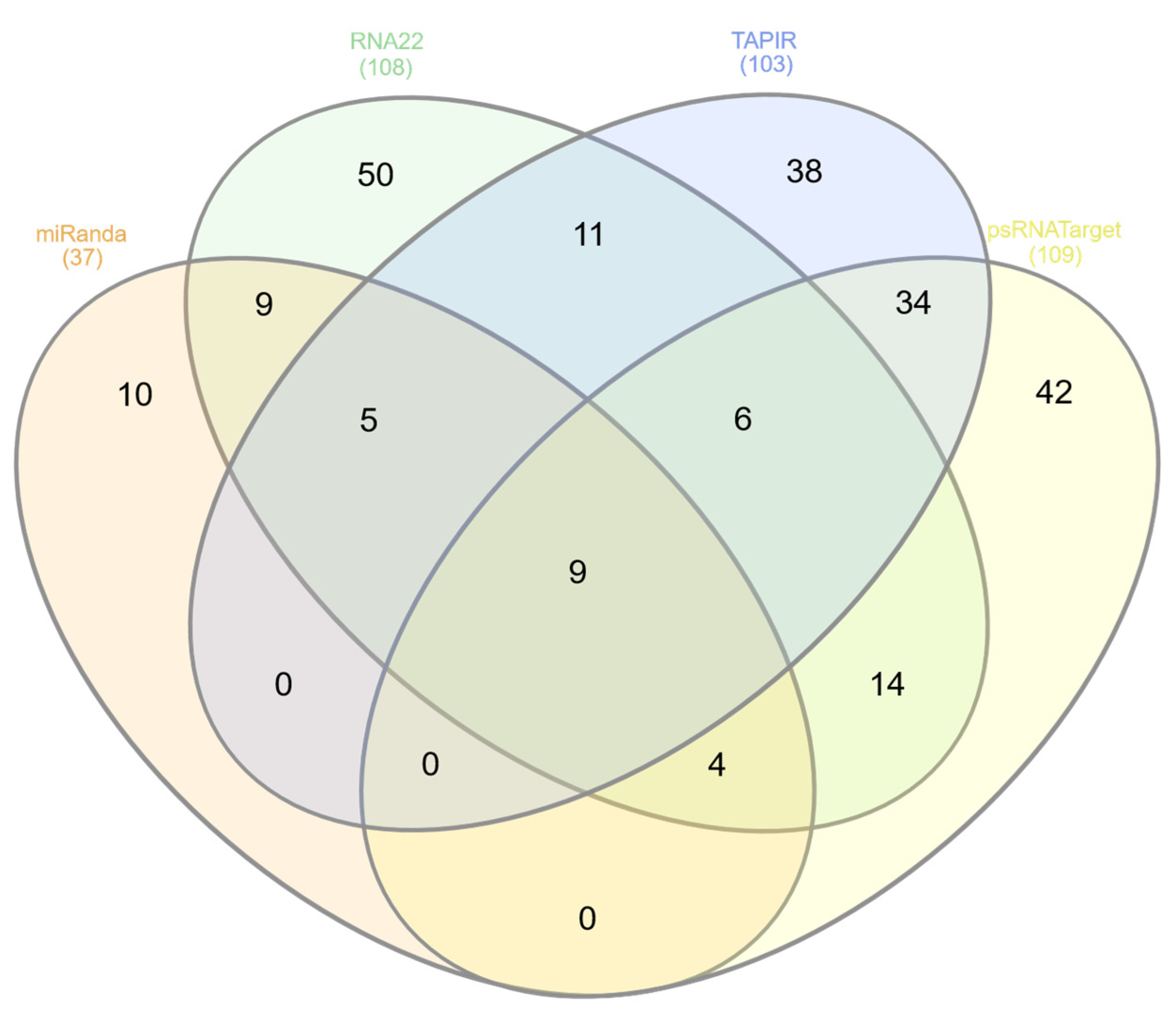
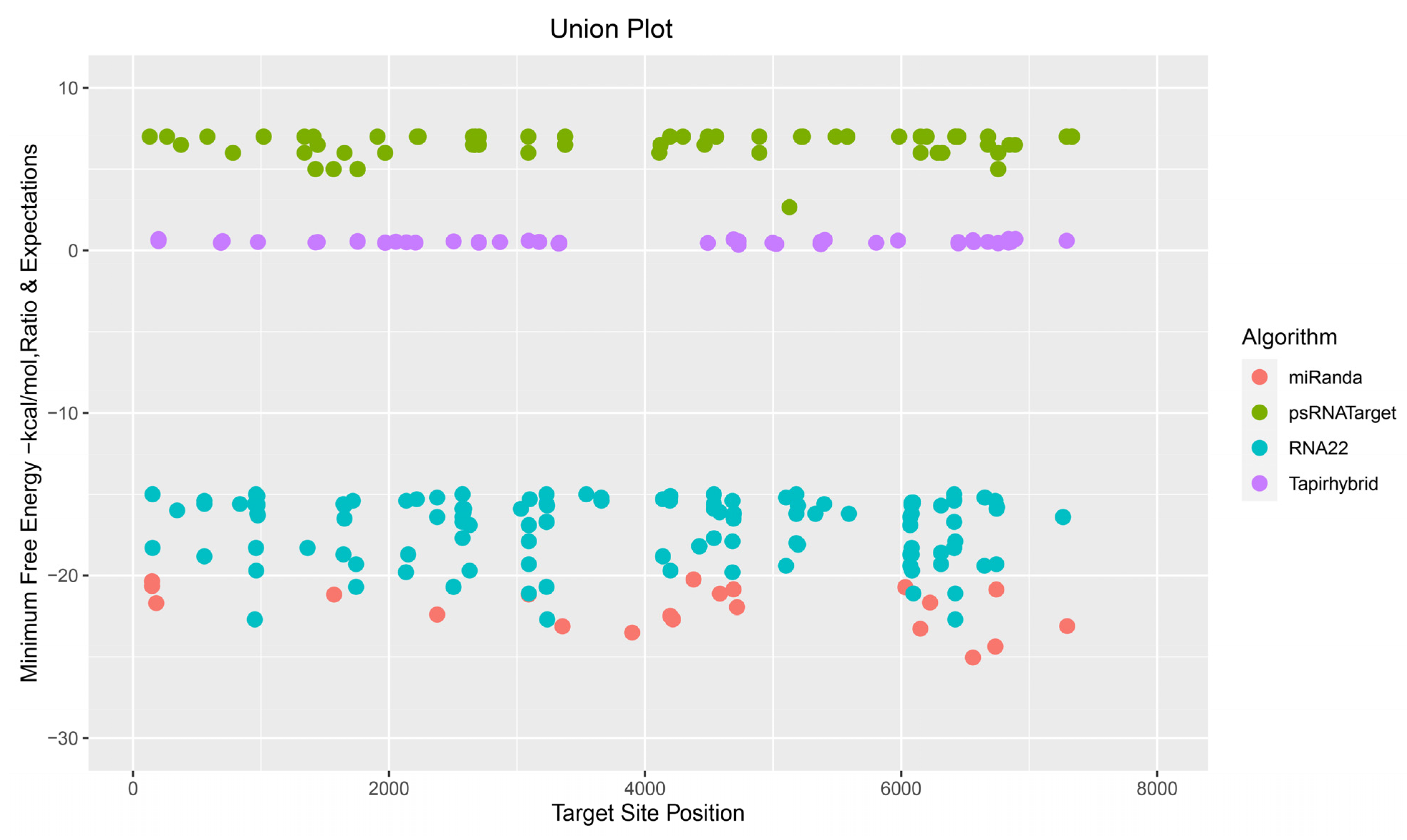
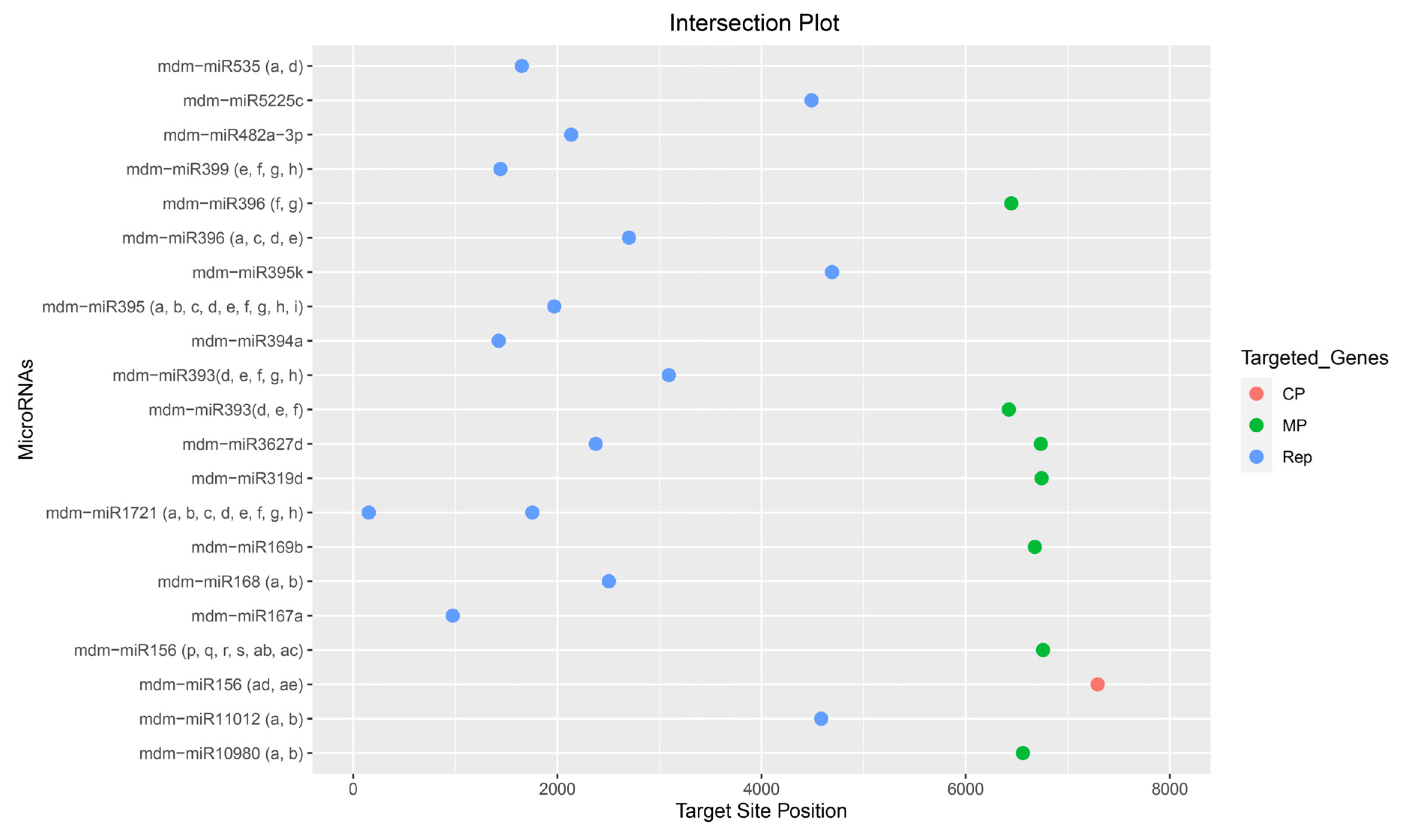
3.5. Evaluation of Common Apple MicroRNAs
3.6. Evaluation and Identification of Consensual Apple MicroRNAs for ACLSV Silencing
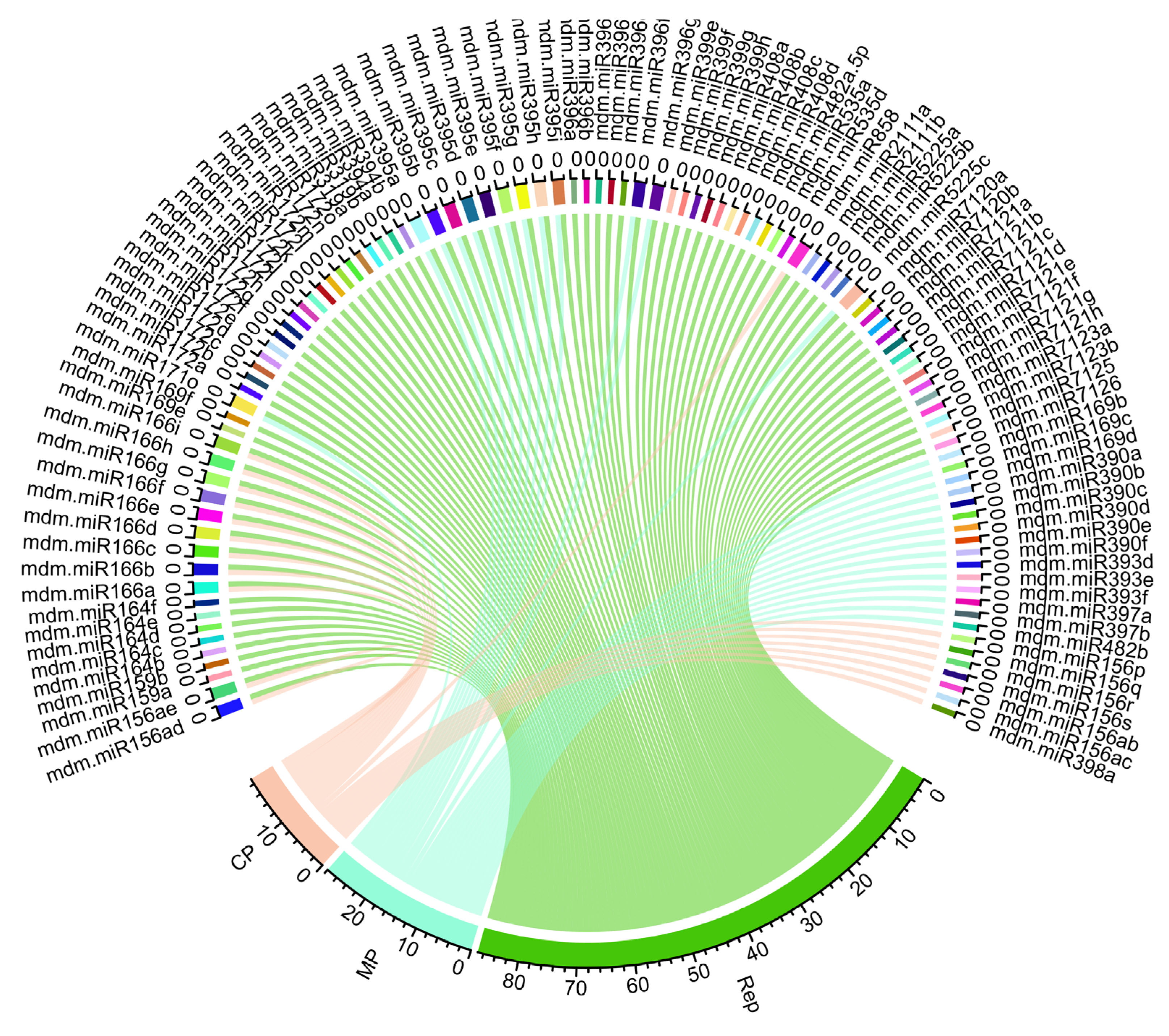
3.7. Construction of Apple mdm-miRNAs-mRNA Regulatory Network
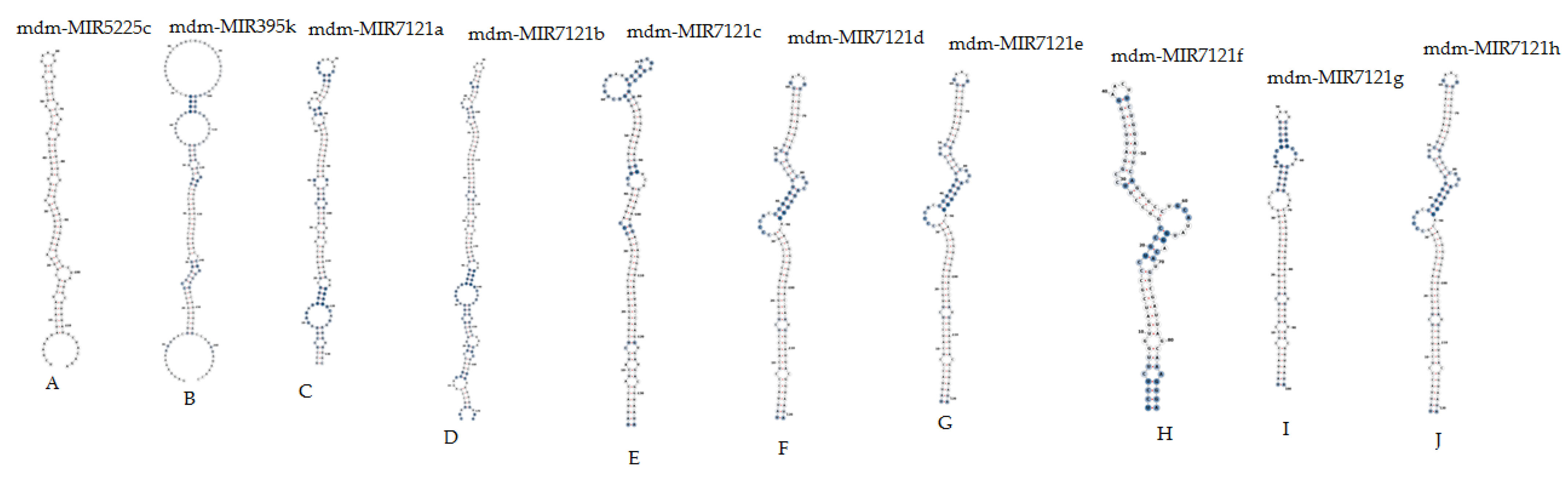
3.8. Secondary Structures of the Consensual RNA
3.9. Assessment of Free Energy
4. Discussion
5. Conclusions and Future Directions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Na, W.; Wolf, J.; Zhang, F.-S. Towards sustainable intensification of apple production in China—Yield gaps and nutrient use efficiency in apple farming systems. J. Integr. Agric. 2016, 15, 716–725. [Google Scholar]
- Shah, Z.A.; Dar, M.A.; Dar, E.A.; Obianefo, C.A.; Bhat, A.H.; Ali, M.T.; El-Sharnouby, M.; Shukry, M.; Kesba, H.; Sayed, S. Sustainable Fruit Growing: An Analysis of Differences in Apple Productivity in the Indian State of Jammu and Kashmir. Sustainability 2022, 14, 14544. [Google Scholar] [CrossRef]
- Chen, Z.; Yu, L.; Liu, W.; Zhang, J.; Wang, N.; Chen, X. Research progress of fruit color development in apple (Malus domestica Borkh.). Plant Physiol. Biochem. 2021, 162, 267–279. [Google Scholar] [CrossRef] [PubMed]
- Velasco, R.; Zharkikh, A.; Affourtit, J.; Dhingra, A.; Cestaro, A.; Kalyanaraman, A.; Fontana, P.; Bhatnagar, S.K.; Troggio, M.; Pruss, D. The genome of the domesticated apple (Malus× domestica Borkh.). Nat. Genet. 2010, 42, 833–839. [Google Scholar] [CrossRef]
- Martelli, G.; Candresse, T.; Namba, S. Trichovirus, a New Genus of Plant Viruses; Springer: Berlin/Heidelberg, Germany, 1994. [Google Scholar]
- Rwahnih, M.A.; Turturo, C.; Minafra, A.; Saldarelli, P.; Myrta, A.; Pallás, V.; Savino, V. Molecular variability of apple chlorotic leaf spot virusin different hosts and geographical regions. J. Plant Pathol. 2004, 86, 117–122. [Google Scholar]
- Abtahi, F.; Shams-Bakhsh, M.; Safaie, N.; Azizi, A.; Autonell, C.R.; Ratti, C. Incidence and genetic diversity of apple chlorotic leaf spot virus in Iran. J. Plant Pathol. 2019, 101, 513–519. [Google Scholar] [CrossRef]
- Katsiani, A.; Maliogka, V.; Candresse, T.; Katis, N. Host-range studies, genetic diversity and evolutionary relationships of ACLSV isolates from ornamental, wild and cultivated R osaceous species. Plant Pathol. 2014, 63, 63–71. [Google Scholar] [CrossRef]
- Yaegashi, H.; Yoshikawa, N.; Candresse, T. Apple chlorotic leaf spot virus in pome fruits. In Virus and Virus-Like Diseases of Pome and Stone Fruits; Hadidi, A., Barba, M., Candresse, T., Jelkmann, W., Eds.; APS Press/American Phytopathological Society: St. Paul, MN, USA, 2011; pp. 17–21. [Google Scholar]
- Guo, W.; Zheng, W.; Wang, M.; Li, X.; Ma, Y.; Dai, H. Genome sequences of three apple chlorotic leaf spot virus isolates from hawthorns in China. PLoS ONE 2016, 11, e0161099. [Google Scholar] [CrossRef]
- Canales, C.; Morán, F.; Olmos, A.; Ruiz-García, A.B. First detection and molecular characterization of Apple stem grooving virus, apple chlorotic leaf spot virus, and apple hammerhead viroid in loquat in Spain. Plants 2021, 10, 2293. [Google Scholar] [CrossRef]
- Dhir, S.; Zaidi, A.A.; Hallan, V. Molecular Characterization and Recombination Analysis of the Complete Genome of Apple Chlorotic Leaf Spot Virus. J. Phytopathol. 2013, 161, 704–712. [Google Scholar] [CrossRef]
- Reinhart, B.J.; Weinstein, E.G.; Rhoades, M.W.; Bartel, B.; Bartel, D.P. MicroRNAs in plants. Genes Dev. 2002, 16, 1616–1626. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Meng, J.; Cui, J.; Luan, Y. Characterization and function of microRNA∗ s in plants. Front. Plant Sci. 2017, 8, 2200. [Google Scholar] [CrossRef]
- Kim, Y.J.; Zheng, B.; Yu, Y.; Won, S.Y.; Mo, B.; Chen, X. The role of Mediator in small and long noncoding RNA production in Arabidopsis thaliana. EMBO J. 2011, 30, 814–822. [Google Scholar] [CrossRef] [PubMed]
- Fang, X.; Cui, Y.; Li, Y.; Qi, Y. Transcription and processing of primary microRNAs are coupled by Elongator complex in Arabidopsis. Nat. Plants 2015, 1, 15075. [Google Scholar] [CrossRef]
- Fang, Y.; Spector, D.L. Identification of nuclear dicing bodies containing proteins for microRNA biogenesis in living Arabidopsis plants. Curr. Biol. 2007, 17, 818–823. [Google Scholar] [CrossRef] [PubMed]
- Manavella, P.A.; Koenig, D.; Weigel, D. Plant secondary siRNA production determined by microRNA-duplex structure. Proc. Natl. Acad. Sci. USA 2012, 109, 2461–2466. [Google Scholar] [CrossRef]
- Trobaugh, D.W.; Klimstra, W.B. MicroRNA regulation of RNA virus replication and pathogenesis. Trends Mol. Med. 2017, 23, 80–93. [Google Scholar] [CrossRef]
- Deng, Z.; Ma, L.; Zhang, P.; Zhu, H. Small RNAs Participate in Plant–Virus Interaction and Their Application in Plant Viral Defense. Int. J. Mol. Sci. 2022, 23, 696. [Google Scholar] [CrossRef]
- Mengistu, A.A.; Tenkegna, T.A. The role of miRNA in plant–virus interaction: A review. Mol. Biol. Rep. 2021, 48, 2853–2861. [Google Scholar] [CrossRef]
- Zhou, L.; Yuan, Q.; Ai, X.; Chen, J.; Lu, Y.; Yan, F. Transgenic Rice Plants Expressing Artificial miRNA Targeting the Rice Stripe Virus MP Gene Are Highly Resistant to the Virus. Biology 2022, 11, 332. [Google Scholar] [CrossRef]
- Miao, S.; Liang, C.; Li, J.; Baker, B.; Luo, L. Polycistronic artificial microRNA-mediated resistance to cucumber green mottle mosaic virus in cucumber. Int. J. Mol. Sci. 2021, 22, 12237. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Li, Y.; Zhang, Y.; Wu, C.; Wang, S.; Hao, L.; Wang, S.; Li, T. Md-miR156ab and Md-miR395 target WRKY transcription factors to influence apple resistance to leaf spot disease. Front. Plant Sci. 2017, 8, 526. [Google Scholar] [CrossRef]
- Qu, D.; Yan, F.; Meng, R.; Jiang, X.; Yang, H.; Gao, Z.; Dong, Y.; Yang, Y.; Zhao, Z. Identification of microRNAs and their targets associated with fruit-bagging and subsequent sunlight re-exposure in the “Granny Smith” apple exocarp using high-throughput sequencing. Front. Plant Sci. 2016, 7, 27. [Google Scholar] [CrossRef] [PubMed]
- Niu, C.; Li, H.; Jiang, L.; Yan, M.; Li, C.; Geng, D.; Xie, Y.; Yan, Y.; Shen, X.; Chen, P. Genome-wide identification of drought-responsive microRNAs in two sets of Malus from interspecific hybrid progenies. Hortic. Res. 2019, 6, 75. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Feng, C.; Zhai, Z.; Peng, X.; Wang, Y.; Sun, Y.; Li, J.; Shen, X.; Xiao, Y.; Zhu, S. The apple microR171i-SCARECROW-LIKE PROTEINS26. 1 module enhances drought stress tolerance by integrating ascorbic acid metabolism. Plant Physiol. 2020, 184, 194–211. [Google Scholar] [CrossRef] [PubMed]
- Tahir, M.M.; Li, S.; Liu, Z.; Fan, L.; Tang, T.; Zhang, X.; Mao, J.; Li, K.; Khan, A.; Shao, Y. Different miRNAs and hormones are involved in PEG-induced inhibition of adventitious root formation in apple. Sci. Hortic. 2022, 303, 111206. [Google Scholar] [CrossRef]
- Kozomara, A.; Birgaoanu, M.; Griffiths-Jones, S. miRBase: From microRNA sequences to function. Nucleic Acids Res. 2019, 47, D155–D162. [Google Scholar] [CrossRef]
- Sayers, E.W.; Beck, J.; Bolton, E.E.; Bourexis, D.; Brister, J.R.; Canese, K.; Comeau, D.C.; Funk, K.; Kim, S.; Klimke, W. Database resources of the national center for biotechnology information. Nucleic Acids Res. 2021, 49, D10. [Google Scholar] [CrossRef]
- Enright, A.; John, B.; Gaul, U.; Tuschl, T.; Sander, C.; Marks, D. MicroRNA targets in Drosophila. Genome Biol. 2003, 4, 1–27. [Google Scholar] [CrossRef]
- John, B.; Enright, A.J.; Aravin, A.; Tuschl, T.; Sander, C.; Marks, D.S. Human microRNA targets. PLoS Biol. 2004, 2, e363. [Google Scholar] [CrossRef]
- Miranda, K.C.; Huynh, T.; Tay, Y.; Ang, Y.-S.; Tam, W.-L.; Thomson, A.M.; Lim, B.; Rigoutsos, I. A pattern-based method for the identification of MicroRNA binding sites and their corresponding heteroduplexes. Cell 2006, 126, 1203–1217. [Google Scholar] [CrossRef]
- Loher, P.; Rigoutsos, I. Interactive exploration of RNA22 microRNA target predictions. Bioinformatics 2012, 28, 3322–3323. [Google Scholar] [CrossRef] [PubMed]
- Bonnet, E.; He, Y.; Billiau, K.; Van de Peer, Y. TAPIR, a web server for the prediction of plant microRNA targets, including target mimics. Bioinformatics 2010, 26, 1566–1568. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Zhao, P.X. psRNATarget: A plant small RNA target analysis server. Nucleic Acids Res. 2011, 39, W155–W159. [Google Scholar] [CrossRef]
- Dai, X.; Zhuang, Z.; Zhao, P.X. psRNATarget: A plant small RNA target analysis server (2017 release). Nucleic Acids Res. 2018, 46, W49–W54. [Google Scholar] [CrossRef]
- Krzywinski, M.; Schein, J.; Birol, I.; Connors, J.; Gascoyne, R.; Horsman, D.; Jones, S.J.; Marra, M.A. Circos: An information aesthetic for comparative genomics. Genome Res. 2009, 19, 1639–1645. [Google Scholar] [CrossRef]
- Lorenz, R.; Bernhart, S.; Siederdissen, C.; Tafer, H.; Flamm, C.; Stadler, P.; Hofacker, I. ViennaRNA package 2.0. Algorithms Mol. Biol. 2011, 6, 26. [Google Scholar] [CrossRef]
- Bernhart, S.H.; Tafer, H.; Mückstein, U.; Flamm, C.; Stadler, P.F.; Hofacker, I.L. Partition function and base pairing probabilities of RNA heterodimers. Algorithms Mol. Biol. 2006, 1, 3. [Google Scholar] [CrossRef] [PubMed]
- Gandrud, C. Reproducible Research with R and RStudio; Chapman and Hall/CRC: Boca Raton, FL, USA, 2018. [Google Scholar]
- German, S.; Candresse, T.; Lanneau, M.; Huet, J.; Pernollet, J.; Dunez, J. Nucleotide sequence and genomic organization of apple chlorotic leaf spot closterovirus. Virology 1990, 179, 104–112. [Google Scholar] [CrossRef] [PubMed]
- Sato, K.; Yoshikawa, N.; Takahashi, T. Complete nucleotide sequence of the genome of an apple isolate of apple chlorotic leaf spot virus. J. Gen. Virol. 1993, 74, 1927–1931. [Google Scholar] [CrossRef]
- Singh, R.M.; Singh, D.; Hallan, V. Movement protein of Apple chlorotic leaf spot virus is genetically unstable and negatively regulated by Ribonuclease E in E. coli. Sci. Rep. 2017, 7, 2133. [Google Scholar] [CrossRef] [PubMed]
- Satoh, H.; Matsuda, H.; Kawamura, T.; Isogai, M.; Yoshikawa, N.; Takahashi, T. Intracellular distribution, cell-to-cell trafficking and tubule-inducing activity of the 50 kDa movement protein of Apple chlorotic leaf spot virus fused to green fluorescent protein. J. Gen. Virol. 2000, 81, 2085–2093. [Google Scholar] [CrossRef] [PubMed]
- Yaegashi, H.; Takahashi, T.; Isogai, M.; Kobori, T.; Ohki, S.; Yoshikawa, N. Apple chlorotic leaf spot virus 50 kDa movement protein acts as a suppressor of systemic silencing without interfering with local silencing in Nicotiana benthamiana. J. Gen. Virol. 2007, 88, 316–324. [Google Scholar] [CrossRef] [PubMed]
- Isogai, M.; Yoshikawa, N. Mapping the RNA-binding domain on the Apple chlorotic leaf spot virus movement protein. J. Gen. Virol. 2005, 86, 225–229. [Google Scholar] [CrossRef]
- Yaegashi, H.; Isogai, M.; Tajima, H.; Sano, T.; Yoshikawa, N. Combinations of two amino acids (Ala40 and Phe75 or Ser40 and Tyr75) in the coat protein of apple chlorotic leaf spot virus are crucial for infectivity. J. Gen. Virol. 2007, 88, 2611–2618. [Google Scholar] [CrossRef]
- Mazeikiene, I.; Siksnianiene, J.B.; Gelvonauskiene, D.; Bendokas, V.; Stanys, V. Prevalence and molecular variability of Apple chlorotic leaf spot virus capsid protein genes in Lithuania. J. Plant Dis. Prot. 2018, 125, 389–396. [Google Scholar] [CrossRef]
- Zhang, B.; Pan, X.; Cox, S.; Cobb, G.; Anderson, T. Evidence that miRNAs are different from other RNAs. Cell. Mol. Life Sci. CMLS 2006, 63, 246–254. [Google Scholar] [CrossRef]
- Prabu, G.; Mandal, A.K.A. Computational identification of miRNAs and their target genes from expressed sequence tags of tea (Camellia sinensis). Genom. Proteom. Bioinform. 2010, 8, 113–121. [Google Scholar] [CrossRef]
- Ossowski, S.; Schwab, R.; Weigel, D. Gene silencing in plants using artificial microRNAs and other small RNAs. Plant J. 2008, 53, 674–690. [Google Scholar] [CrossRef]
- Mathews, D.H.; Sabina, J.; Zuker, M.; Turner, D.H. Expanded sequence dependence of thermodynamic parameters improves prediction of RNA secondary structure. J. Mol. Biol. 1999, 288, 911–940. [Google Scholar] [CrossRef]
- Niu, Q.-W.; Lin, S.-S.; Reyes, J.L.; Chen, K.-C.; Wu, H.-W.; Yeh, S.-D.; Chua, N.-H. Expression of artificial microRNAs in transgenic Arabidopsis thaliana confers virus resistance. Nat. Biotechnol. 2006, 24, 1420–1428. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, M.A.; Ali, B.; Brown, J.K.; Shahid, I.; Yu, N. In Silico Identification of Cassava Genome-Encoded MicroRNAs with Predicted Potential for Targeting the ICMV-Kerala Begomoviral Pathogen of Cassava. Viruses 2023, 15, 486. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, N.A.; Ngah, N.M.F.N.C.; Abas, A.; Talip, N.; Sarian, M.N.; Hamezah, H.S.; Harun, S.; Bunawan, H. Candidate miRNAs from Oryza sativa for Silencing the Rice Tungro Viruses. Agriculture 2023, 13, 651. [Google Scholar] [CrossRef]
- Ashraf, M.A.; Tariq, H.K.; Hu, X.-W.; Khan, J.; Zou, Z. Computational Biology and Machine Learning Approaches Identify Rubber Tree (Hevea brasiliensis Muell. Arg.) Genome Encoded MicroRNAs Targeting Rubber Tree Virus 1. Appl. Sci. 2022, 12, 12908. [Google Scholar] [CrossRef]
- Ashraf, M.A.; Feng, X.; Hu, X.; Ashraf, F.; Shen, L.; Iqbal, M.S.; Zhang, S. In silico identification of sugarcane (Saccharum officinarum L.) genome encoded microRNAs targeting sugarcane bacilliform virus. PLoS ONE 2022, 17, e0261807. [Google Scholar] [CrossRef]
- Ashraf, M.A.; Ashraf, F.; Feng, X.; Hu, X.; Shen, L.; Khan, J.; Zhang, S. Potential targets for evaluation of sugarcane yellow leaf virus resistance in sugarcane cultivars: In silico sugarcane miRNA and target network prediction. Biotechnol. Biotechnol. Equip. 2021, 35, 1980–1991. [Google Scholar] [CrossRef]
- Ashraf, F.; Ashraf, M.A.; Hu, X.; Zhang, S. A novel computational approach to the silencing of Sugarcane Bacilliform Guadeloupe A Virus determines potential host-derived MicroRNAs in sugarcane (Saccharum officinarum L.). PeerJ 2020, 8, e8359. [Google Scholar] [CrossRef]
- Gaafar, Y.Z.A.; Ziebell, H. Novel targets for engineering Physostegia chlorotic mottle and tomato brown rugose fruit virus-resistant tomatoes: In silico prediction of tomato microRNA targets. PeerJ 2020, 8, e10096. [Google Scholar] [CrossRef]
- Petchthai, U.; Yee, C.S.L.; Wong, S.-M. Resistance to CymMV and ORSV in artificial microRNA transgenic Nicotiana benthamiana plants. Sci. Rep. 2018, 8, 9958. [Google Scholar] [CrossRef]
- Zhang, D.; Zhang, N.; Shen, W.; Li, J.-F. Engineered artificial microRNA precursors facilitate cloning and gene silencing in arabidopsis and rice. Int. J. Mol. Sci. 2019, 20, 5620. [Google Scholar] [CrossRef]
- Zhang, N.; Zhang, D.; Chen, S.L.; Gong, B.-Q.; Guo, Y.; Xu, L.; Zhang, X.-N.; Li, J.-F. Engineering artificial microRNAs for multiplex gene silencing and simplified transgenic screen. Plant Physiol. 2018, 178, 989–1001. [Google Scholar] [CrossRef]
- Yasir, M.; Motawaa, M.; Wang, Q.; Zhang, X.; Khalid, A.; Cai, X.; Li, F. Simple webserver-facilitated method to design and synthesize artificial miRNA gene and its application in engineering viral resistance. Plants 2022, 11, 2125. [Google Scholar] [CrossRef]
- Ali, I.; Amin, I.; Briddon, R.W.; Mansoor, S. Artificial microRNA-mediated resistance against the monopartite begomovirus Cotton leaf curl Burewala virus. Virol. J. 2013, 10, 231. [Google Scholar] [CrossRef] [PubMed]
- Duan, C.-G.; Wang, C.-H.; Fang, R.-X.; Guo, H.-S. Artificial microRNAs highly accessible to targets confer efficient virus resistance in plants. J. Virol. 2008, 82, 11084–11095. [Google Scholar] [CrossRef] [PubMed]
- Dweep, H.; Sticht, C.; Gretz, N. In-silico algorithms for the screening of possible microRNA binding sites and their interactions. Curr. Genom. 2013, 14, 127–136. [Google Scholar] [CrossRef] [PubMed]
- Thody, J.; Moulton, V.; Mohorianu, I. PAREameters: A tool for computational inference of plant miRNA–mRNA targeting rules using small RNA and degradome sequencing data. Nucleic Acids Res. 2020, 48, 2258–2270. [Google Scholar] [CrossRef]
- Pinzón, N.; Li, B.; Martinez, L.; Sergeeva, A.; Presumey, J.; Apparailly, F.; Seitz, H. microRNA target prediction programs predict many false positives. Genome Res. 2017, 27, 234–245. [Google Scholar] [CrossRef]
- Kertesz, M.; Iovino, N.; Unnerstall, U.; Gaul, U.; Segal, E. The role of site accessibility in microRNA target recognition. Nat. Genet. 2007, 39, 1278–1284. [Google Scholar] [CrossRef]
- Golyshev, V.; Pyshnyi, D.; Lomzov, A. Calculation of Energy for RNA/RNA and DNA/RNA Duplex Formation by Molecular Dynamics Simulation. Mol. Biol. 2021, 55, 927–940. [Google Scholar] [CrossRef]
- Ghoshal, A.; Shankar, R.; Bagchi, S.; Grama, A.; Chaterji, S. MicroRNA target prediction using thermodynamic and sequence curves. BMC Genom. 2015, 16, 999. [Google Scholar] [CrossRef]
- Kaja, E.; Szcześniak, M.W.; Jensen, P.J.; Axtell, M.J.; McNellis, T.; Makałowska, I. Identification of apple miRNAs and their potential role in fire blight resistance. Tree Genet. Genomes 2015, 11, 812. [Google Scholar] [CrossRef]
- Liu, X.-X.; Luo, X.-F.; Luo, K.-X.; Liu, Y.-L.; Pan, T.; Li, Z.-Z.; Duns, G.J.; He, F.-L.; Qin, Z.-D. Small RNA sequencing reveals dynamic microRNA expression of important nutrient metabolism during development of Camellia oleifera fruit. Int. J. Biol. Sci. 2019, 15, 416. [Google Scholar] [CrossRef]
- Witkos, T.M.; Koscianska, E.; Krzyzosiak, W.J. Practical aspects of microRNA target prediction. Curr. Mol. Med. 2011, 11, 93–109. [Google Scholar] [CrossRef]
- Niu, F.; Pan, S.; Wu, Z.; Jiang, D.; Li, S. Complete nucleotide sequences of the genomes of two isolates of apple chlorotic leaf spot virus from peach (Prunus persica) in China. Arch. Virol. 2012, 157, 783–786. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Wang, G.; Hu, H.; Tian, R.; Hong, N. The genome sequences of three isolates of Apple chlorotic leaf spot virus from pear (Pyrus sp.) in China. Can. J. Plant Pathol. 2014, 36, 396–402. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Algorithms | Parameter | Features | Availability |
|---|---|---|---|
| miRanda | Score threshold = 140, Free energy = −20 Kcal/mol, Gap open penalty = −9.00 Gap extend penalty = −4.00 | Seed-based interaction, Target site accessibility, free energy of RNA-RNA duplex, conservation | http://www.microrna.org/ (accessed 26 January 2023) [31,32] |
| RNA22 | Folding energy = −15 Kcal/mol Number of paired-up bases = 12, Sensitivity (63%), Specificity (61%), | Non-seed-based interaction, Site complementarity, Target site multiplicity, Pattern recognition, Folding energy of heteroduplex | https://cm.jefferson.edu/rna22/Interactive/ (accessed on 22 October 2022) [33,34] |
| TAPIR | Free energy = −20 Kcal/mol, Hit per target = 1 | Seed paring, Free energy of duplex, Multiple target sites, | http://bibiserv.techfak.uni-bielefeld.de/rnahybrid (accessed on 9 November 2022) [35] |
| psRNATarget | Expectation Score = 6.5, HSP size = 19, Penalty for G:U pair = 0.5 Penalty for opening gap = 2 | Multiplicity of target site, Translation inhibition, Target accessibility, Complementarity scoring | https://www.zhaolab.org/psRNATarget/analysis?function=2 (accessed on 9 November 2022) [36,37] |
| Apple miRNAs | Position miRanda | Position RNA22 | Position TAPIR | Position psRNATarget | MFE * miRanda | MFE ** RNA22 | MFE Ratio TAPIR | Expectation psRNATarget |
|---|---|---|---|---|---|---|---|---|
| mdm-miR156 (p, q, r, s) | 6758 | 6758 | 0.44 | 6.00 | ||||
| mdm-miR156 (ab, ac) | 6758 | 6758 | 0.46 | 5.00 | ||||
| mdm-156 (ad, ae) | 7293 | 7293 | 0.60 | 7.00 | ||||
| mes-miR167a | 975 | 976 | −16.30 | 0.52 | ||||
| mdm-miR168 (a, b) | 2504 | 2505 | −20.70 | 0.56 | ||||
| mdm-miR169b | 6678 | 6678 | 0.53 | 6.50 | ||||
| mdm-miR319d | 6744 | 6744 | −20.86 | −18.10 | ||||
| mdm-miR393 (d, e, f) | 6423 | 6423 | −19.30 | 7.00 | ||||
| mdm-miR393 (d, e, f) | 3092 | 3091 | −22.70 | 0.60 | ||||
| mdm-miR393 (g, h) | 3091 | 3092 | −21.17 | −21.11 | ||||
| mdm-miR394 (a, b) | 1426 | 1426 | 0.49 | 5.00 | ||||
| mdm-miR395 (a, b, c, d, e, f, g, h, i) | 1970 | 1970 | 0.47 | 6.00 | ||||
| mdm-miR395k | 4691 | 4691 | 4691 | −20.85 | −18.00 | 0.68 | ||
| mdm-miR396 (a, c, d, e) | 2702 | 2702 | 0.52 | 6.50 | ||||
| mdm-miR396 (f, g) | 6447 | 6447 | 0.43 | 7.00 | ||||
| mdm-399 (e, f, g, h) | 1443 | 1443 | 0.52 | 6.50 | ||||
| mdm-482a-3p | 2135 | 2136 | −18.30 | 0.48 | ||||
| mdm-482b | 2207 | 2207 | 0.34 | 6.00 | ||||
| mdm-535a | 1652 | 1652 | −18.60 | 6.00 | ||||
| mdm-535b | 1652 | 1652 | −17.90 | 6.50 | ||||
| mdm-miR3627d | 2376 | 2376 | −22.40 | −19.30 | ||||
| mdm-miR3627d (1) | 6736 | 6736 | −24.37 | −19.80 | ||||
| mdm-5225c | 4490 | 4490 | 0.46 | 7.00 | ||||
| mdm-7121 (a, b, c) | 149 | 153 | 1755 | 1755 | −20.63 | −22.40 | 0.58 | 5.00 |
| mdm-miR7121 (d, e, f, g, h) | 1755 | 1755 | 0.58 | 5.00 | ||||
| mdm-miR10980 (a, b) | 6561 | 6561 | −25.04 | 0.63 | ||||
| mdm-miR11012 (a, b) | 4585 | 4582 | −21.11 | −18.82 |
| miRNA ID | Accession IDs | Length Precursor | MFE/Kcal/mol | AMFE | MFEI | (G + C)% |
|---|---|---|---|---|---|---|
| mdm-MIR5225c | MI0023156 | 119 nt | −51.30 | −43.10 | −0.85 | 50.42 |
| mdm-MIR395k | MI0035639 | 168 nt | −43.27 | −25.75 | −0.68 | 37.50 |
| mdm-MIR7121a | MI0023144 | 132 nt | −49.40 | −37.42 | −0.79 | 46.97 |
| mdm-MIR7121b | MI0023145 | 172 nt | −70.60 | −41.04 | −0.85 | 48.26 |
| mdm-MIR7121c | MI0023146 | 135 nt | −71.30 | −52.81 | −1.09 | 48.15 |
| mdm-MIR7121d | MI0023147 | 121 nt | −67.50 | −55.78 | −1.08 | 51.24 |
| mdm-MIR7121e | MI0023148 | 121 nt | −67.50 | −55.78 | −1.08 | 51.24 |
| mdm-MIR7121f | MI0023149 | 88 nt | −39.90 | −45.34 | −0.79 | 56.82 |
| mdm-MIR7121g | MI0023150 | 100 nt | −45.80 | −45.80 | −0.89 | 51.00 |
| mdm-MIR7121h | MI0023151 | 121 nt | −67.50 | −55.78 | −1.08 | 51.24 |
| Apple Mature miRNA ID | Accession ID | Mdm-miRNA-Target Sequence (5′–3′) | ΔG Duplex (Kcal/mol) |
|---|---|---|---|
| mdm-miR5225c | MIMAT0026052 | 5′ UCUGUCGUGGGUGAGAUGGUGC 3′ 5′ GAAGCAGTGTACCCAAGACATA 3′ | −15.90 |
| mdm-miR395k | MIMAT0043586 | 5′ GUUUCCUCAAACACUUCAUU 3′ 5′ AGGCAGGAGTTTGAGGAAAC 3′ | −18.30 |
| mdm-miR7121a | MIMAT0026040 | 5′ UCCUCUUGGUGAUCGCCCUGU 3′ 5′ AAAGGGAGTTCATCGAGAGAA 3′ | −22.10 |
| mdm-miR7121b | MIMAT0026041 | 5′ UCCUCUUGGUGAUCGCCCUGU 3′ 5′ AAAGGGAGTTCATCGAGAGAA 3′ | −22.10 |
| mdm-miR7121c | MIMAT0026042 | 5′ UCCUCUUGGUGAUCGCCCUGU 3′ 5′ AAAGGGAGTTCATCGAGAGAA 3′ | −22.10 |
| mdm-miR7121d | MIMAT0026043 | 5′ UCCUCUUGGUGAUCGCCCUGC 3′ 5′ AAAGGGAGTTCATCGAGAGAA 3′ | −22.10 |
| mdm-miR7121e | MIMAT0026044 | 5′ UCCUCUUGGUGAUCGCCCUGC 3′ 5′ AAAGGGAGTTCATCGAGAGAA 3′ | −22.10 |
| mdm-miR7121f | MIMAT0026045 | 5′ UCCUCUUGGUGAUCGCCCUGC 3′ 5′ AAAGGGAGTTCATCGAGAGAA 3′ | −22.10 |
| mdm-miR7121g | MIMAT0026046 | 5′ UCCUCUUGGUGAUCGCCCUGC 3′ 5′ AAAGGGAGTTCATCGAGAGAA 3′ | −22.10 |
| mdm-miR7121h | MIMAT0026047 | 5′ UCCUCUUGGUGAUCGCCCUGC 3′ 5′ AAAGGGAGTTCATCGAGAGAA 3′ | −22.10 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ashraf, M.A.; Murtaza, N.; Brown, J.K.; Yu, N. In Silico Apple Genome-Encoded MicroRNA Target Binding Sites Targeting Apple Chlorotic Leaf Spot Virus. Horticulturae 2023, 9, 808. https://doi.org/10.3390/horticulturae9070808
Ashraf MA, Murtaza N, Brown JK, Yu N. In Silico Apple Genome-Encoded MicroRNA Target Binding Sites Targeting Apple Chlorotic Leaf Spot Virus. Horticulturae. 2023; 9(7):808. https://doi.org/10.3390/horticulturae9070808
Chicago/Turabian StyleAshraf, Muhammad Aleem, Nimra Murtaza, Judith K. Brown, and Naitong Yu. 2023. "In Silico Apple Genome-Encoded MicroRNA Target Binding Sites Targeting Apple Chlorotic Leaf Spot Virus" Horticulturae 9, no. 7: 808. https://doi.org/10.3390/horticulturae9070808
APA StyleAshraf, M. A., Murtaza, N., Brown, J. K., & Yu, N. (2023). In Silico Apple Genome-Encoded MicroRNA Target Binding Sites Targeting Apple Chlorotic Leaf Spot Virus. Horticulturae, 9(7), 808. https://doi.org/10.3390/horticulturae9070808