
Detection by Sensitive Real-Time Reverse Transcription Loop-Mediated Isothermal Amplification of Olive Leaf Yellowing Associated Virus and Its Incidence in Italy and Spain

 , , , , , and
, , , , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Real-Time RT-LAMP Assay Development for Fast OLYaV Detection
2.1.1. Source of Viral Material
2.1.2. Primer Design and OLYaV Detection Using Canonical RT-PCR
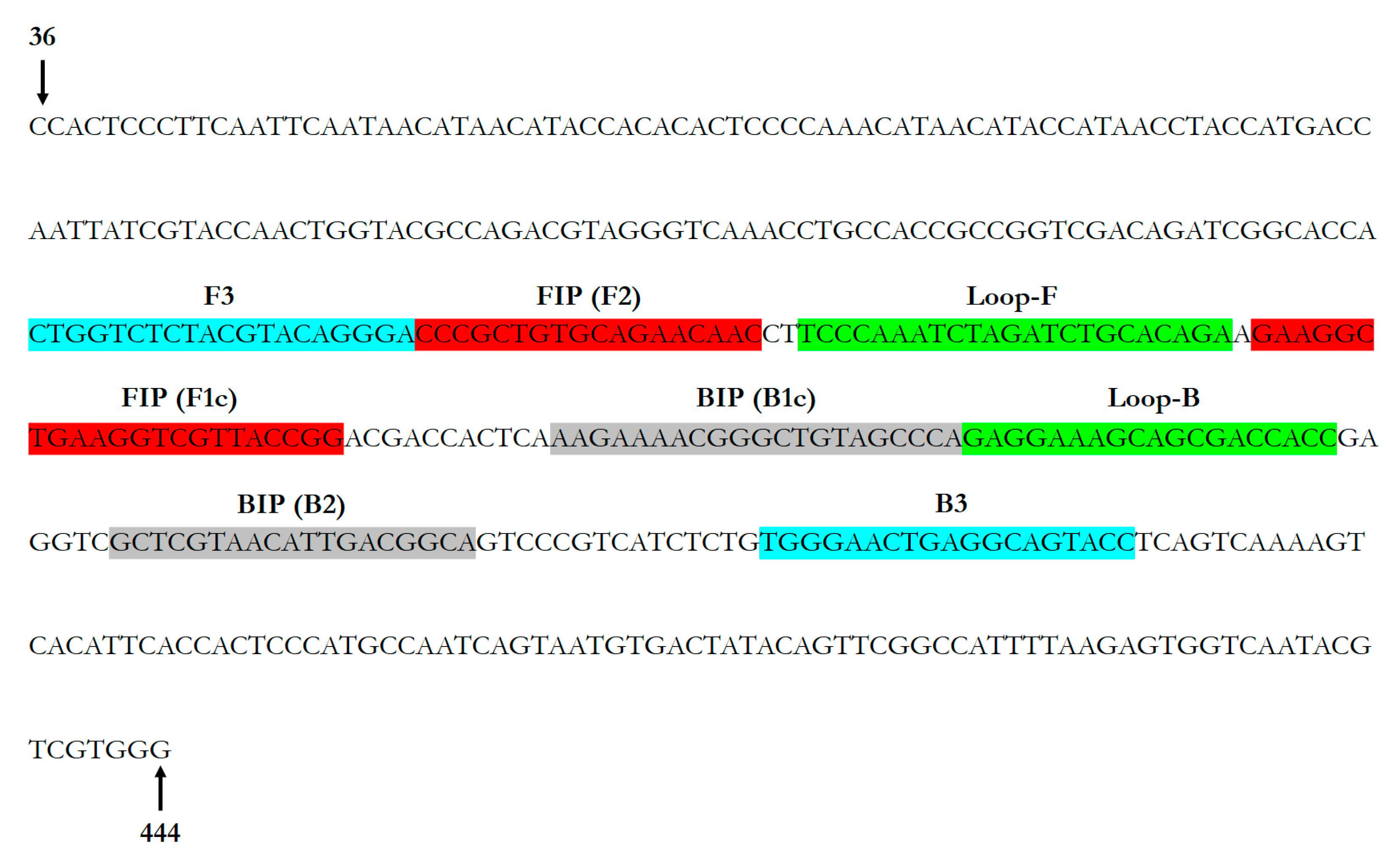
2.1.3. OLYaV Real-Time RT-LAMP Primer Design
2.1.4. OLYaV Real-Time RT-LAMP Assay Optimization
2.1.5. Sensitivity and Comparison of OLYaV Real-Time RT-LAMP Assay to Conventional RT-PCR
2.1.6. Specificity of OLYaV Real-Time RT-LAMP Assay
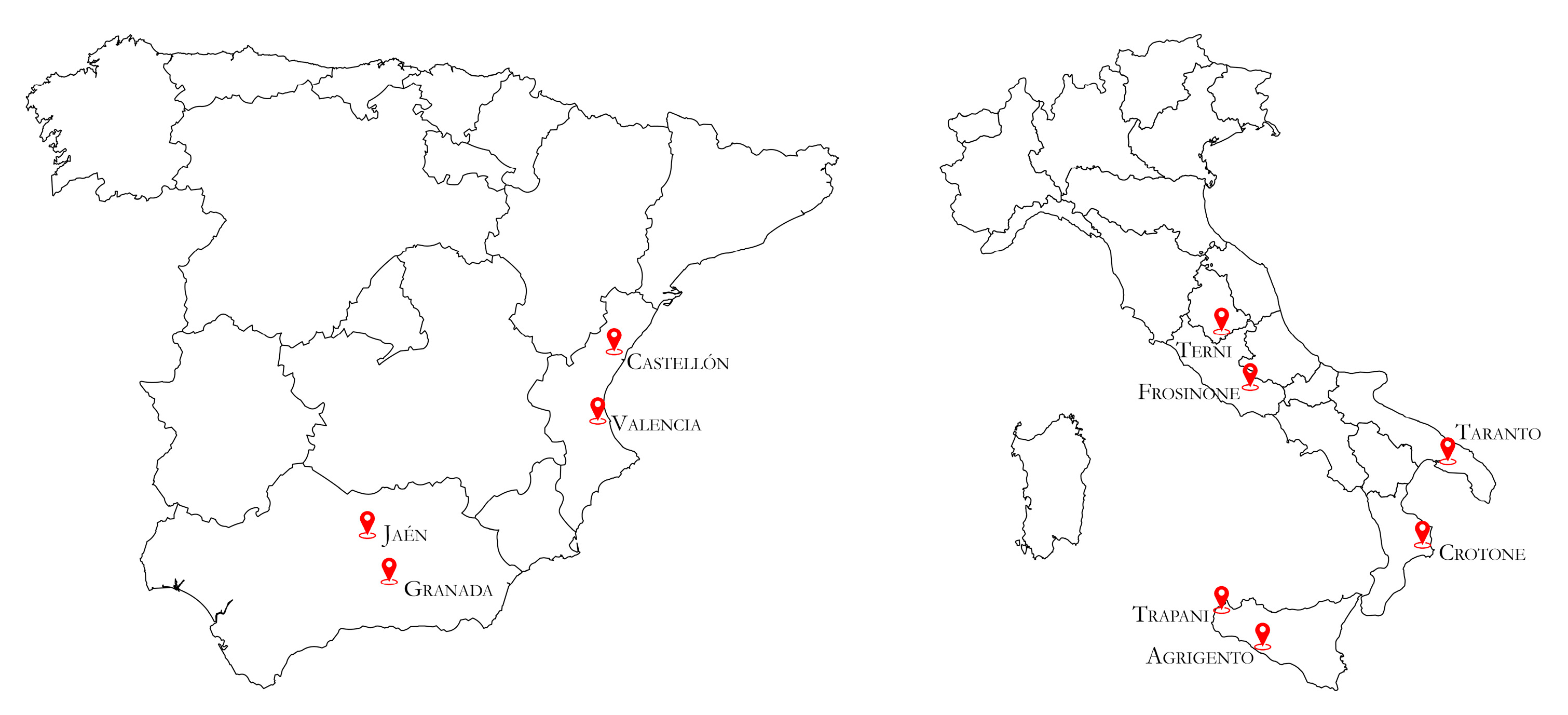
2.2. Evaluation of OLYaV Incidence in Italy and Spain by Real Time RT-LAMP Assay on Symptomatic and Asymptomatic Olive Trees
3. Results
3.1. Real-Time RT-LAMP Assay Development for Fast OLYaV Detection
3.1.1. Primer Design and OLYaV Detection Using Canonical RT-PCR
3.1.2. OLYaV Real-Time RT-LAMP Primer Design
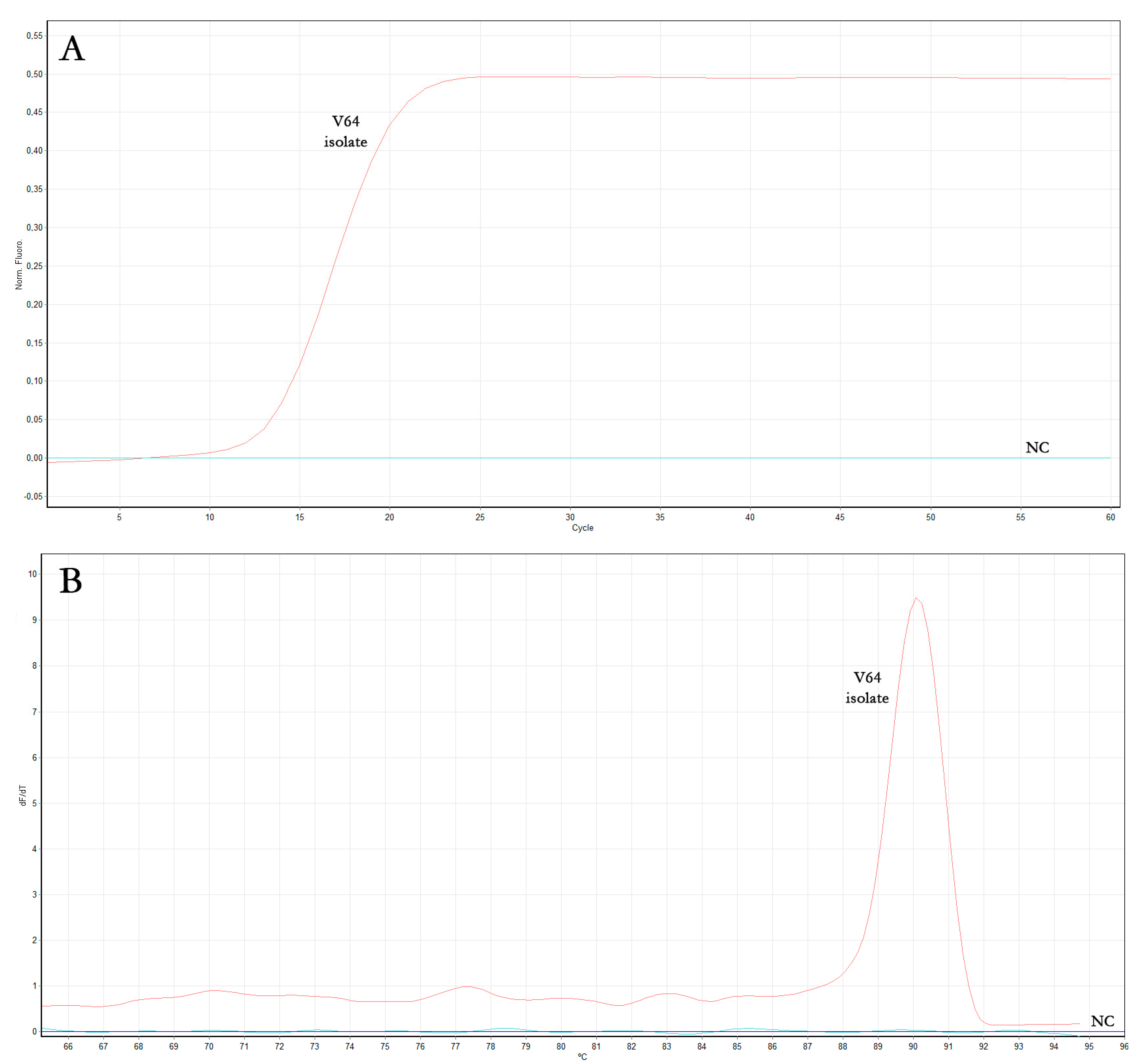
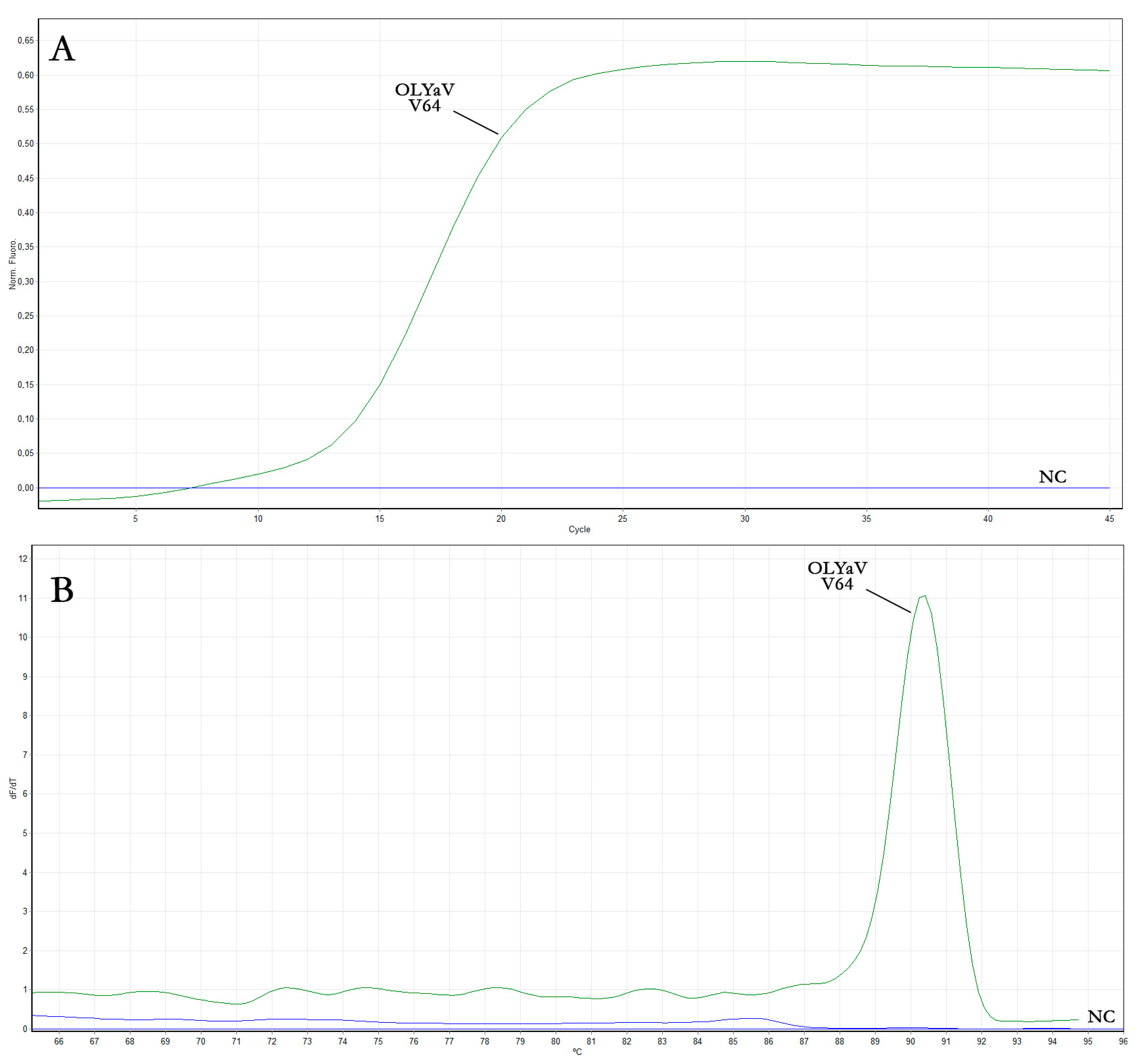
3.1.3. OLYaV Real-Time RT-LAMP Assay Optimization
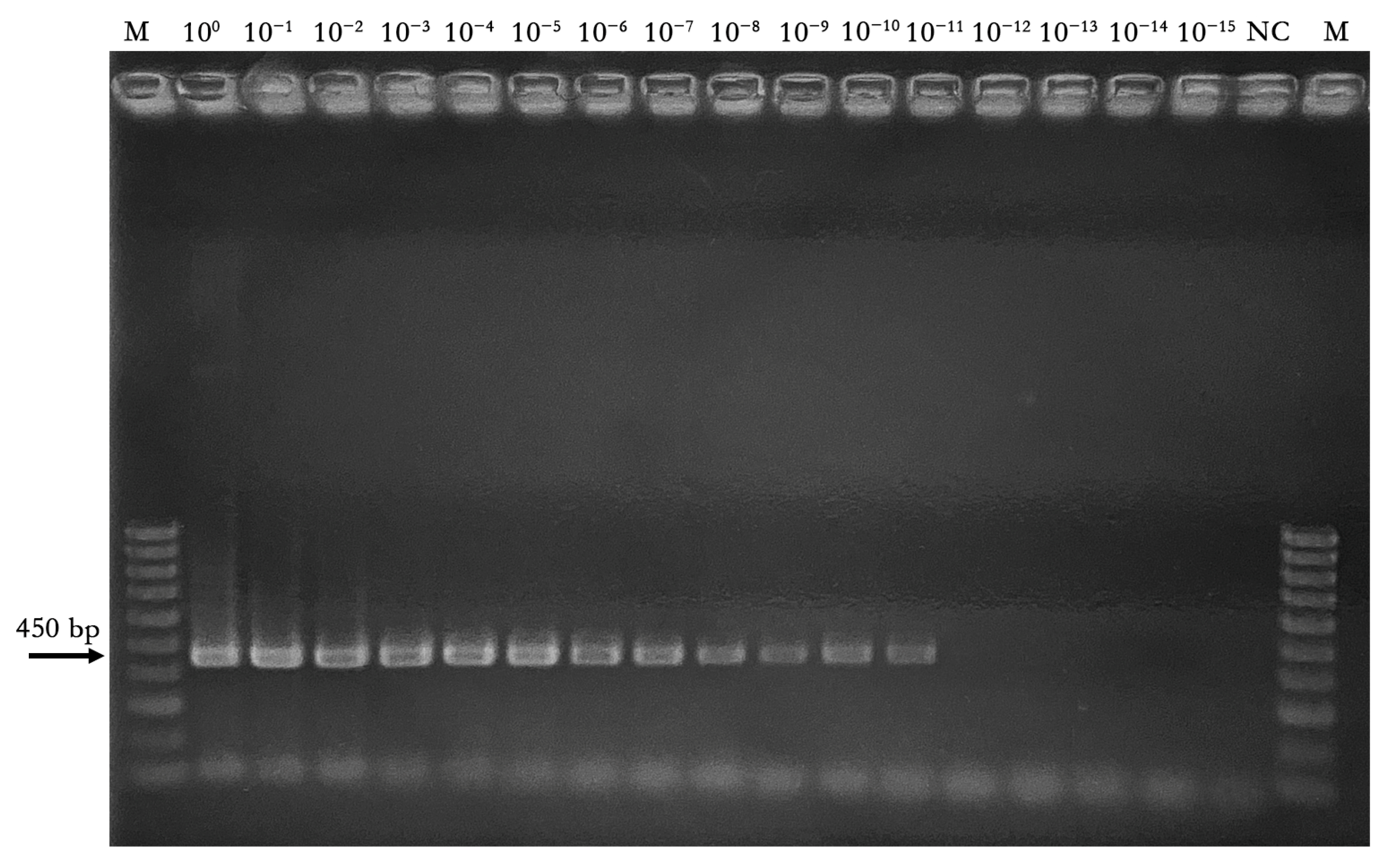
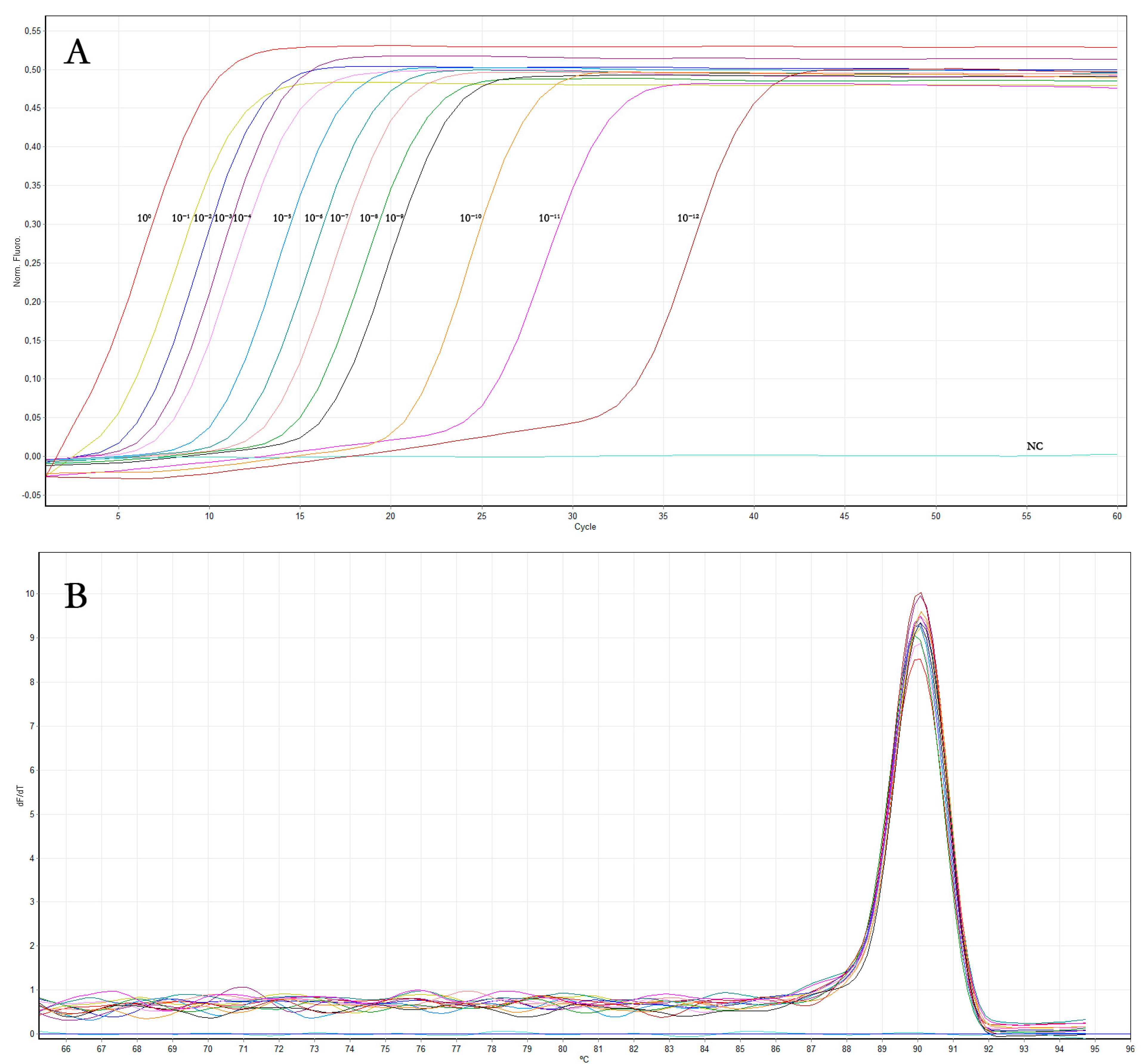
3.1.4. Sensitivity and Comparison of OLYaV Real-Time RT-LAMP Assay to Conventional End-Point RT-PCR
3.1.5. Specificity of OLYaV Real-Time LAMP Assay
3.2. Evaluation of OLYaV Incidence in Italy and Spain by Real Time RT-LAMP Assay on Symptomatic and Asymptomatic Olive Trees
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rapoport, H.F.; Fabbri, A.; Sebastiani, L. Olive biology. In The Olive Tree Genome; Springer: Cham, Switzerland, 2016; pp. 13–25. [Google Scholar] [CrossRef]
- Fraga, H.; Moriondo, M.; Leolini, L.; Santos, J.A. Mediterranean olive orchards under climate change: A review of future impacts and adaptation strategies. Agronomy 2020, 11, 56. [Google Scholar] [CrossRef]
- FAO (Food and Agriculture Organization). Available online: http://www.fao.org/faostat/en (accessed on 1 February 2023).
- European Commission. Available online: https://commission.europa.eu (accessed on 1 February 2023).
- Lodolini, E.M.; Polverigiani, S.; Ali, S.; Mutawea, M.; Qutub, M.; Pierini, F.; Neri, D. Effect of complementary irrigation on yield components and alternate bearing of a traditional olive orchard in semi-arid conditions. Spanish J. Agric. Res. 2016, 14, 1203. [Google Scholar] [CrossRef]
- Marra, F.P.; Marino, G.; Marchese, A.; Caruso, T. Effects of different irrigation regimes on a super-high-density olive grove cv. “Arbequina”: Vegetative growth, productivity and polyphenol content of the oil. Irrig. Sci. 2016, 34, 313–325. [Google Scholar] [CrossRef]
- Beyá-Marshall, V.; Herrera, J.; Fichet, T.; Trentacoste, E.R.; Kremer, C. The effect of water status on productive and flowering variables in young ‘Arbequina’olive trees under limited irrigation water availability in a semiarid region of Chile. Hortic. Environ. Biotechnol. 2018, 59, 815–826. [Google Scholar] [CrossRef]
- Rallo, L.; Barranco, D.; Castro-García, S.; Connor, D.J.; Gómez del Campo, M.; Rallo, P. High-density olive plantations. Hortic. Rev. 2013, 41, 303–384. [Google Scholar] [CrossRef]
- Connor, D.J.; Gómez-del-Campo, M.; Rousseaux, M.C.; Searles, P.S. Structure, management and productivity of hedgerow olive orchards: A review. Sci. Hortic. 2014, 169, 71–93. [Google Scholar] [CrossRef]
- Visioli, F.; Poli, A.; Gall, C. Antioxidant and other biological activities of phenols from olives and olive oil. Med. Res. Rev. 2001, 22, 65–75. [Google Scholar] [CrossRef]
- Kiritsakis, A. Olive Oil, Table Olive, Olive Paste; Printing Akritidi Sindos: Thessaloniki, Greece, 2007. [Google Scholar]
- Kiritsakis, A.; Shahidi, F. Olives and Olive Oil as Functional Foods: Bioactivity, Chemistry and Processing; John Wiley & Sons: Oxford, UK, 2017. [Google Scholar]
- Tous, J.; Romero, A.; Hermoso, J.F. New trends in olive orchard design for continuous mechanical harvesting. Adv. Hortic. Sci. 2010, 24, 43–52. Available online: http://digital.casalini.it/10.1400/132342 (accessed on 1 February 2023).
- Montes-Osuna, N.; Mercado-Blanco, J. Verticillium wilt of olive and its control: What did we learn during the last decade? Plants 2020, 9, 735. [Google Scholar] [CrossRef]
- Saponari, M.; Boscia, D.; Nigro, F.; Martelli, G.P. Identification of DNA sequences related to Xylella fastidiosa in oleander, almond and olive trees exhibiting leaf scorch symptoms in Apulia (Southern Italy). J. Plant Pathol. 2013, 95, 668. [Google Scholar] [CrossRef]
- Martelli, G.P. Infectious diseases and certification of olive: An overview. EPPO Bull. 1999, 29, 127–133. [Google Scholar] [CrossRef]
- Bertacca, S.; Caruso, A.G.; Trippa, D.; Marchese, A.; Giovino, A.; Matic, S.; Noris, E.; Font San Ambrosio, M.I.; Alfaro, A.; Panno, S.; et al. Development of a Real-Time Loop-Mediated Isothermal Amplification Assay for the Rapid Detection of Olea Europaea Geminivirus. Plants 2022, 11, 660. [Google Scholar] [CrossRef]
- Alabi, O.J.; Diaz-Lara, A.; Erickson, T.M.; Al Rwahnih, M. Olea europaea geminivirus is present in a germplasm repository and in California and Texas olive (Olea europaea L.) groves. Arch. Virol. 2021, 166, 3399–3404. [Google Scholar] [CrossRef]
- Felix, M.R.F.; Clara, M.I.E. Two Necrovirus isolates with properties of Olive latent virus 1 and of Tobacco necrosis virus from olive in Portugal. In IV International Symposium on Olive Growing; Vitagliano, C., Martelli, G.P., Eds.; ISHS Acta Horticulturae: Valenzano, Italy, 2000; pp. 725–728. [Google Scholar] [CrossRef]
- Al Abdullah, A.; Minafra, A.; Elbeaino, T.; Saponari, M.; De Stradis, A.; Savino, V.; Martelli, G.P. Complete nucleotide sequence of Olive latent virus 3, representing a new genome structure in the genus Marafivirus. Virus Res. 2010, 152, 10–17. [Google Scholar] [CrossRef]
- Loconsole, G.; Saponari, M.; Faggioli, F.; Albanese, G.; Bouyahia, H.; Elbeaino, T.; Materazzi, A.; Nuzzaci, M.; Prota, V.; Romanazzi, G.; et al. Inter-laboratory validation of PCR-based protocol for detection of olive viruses. EPPO Bull. 2010, 40, 423–428. [Google Scholar] [CrossRef]
- Chiumenti, M.; Greco, C.; De Stradis, A.; Loconsole, G.; Cavalieri, V.; Altamura, G.; Zicca, S.; Saldarelli, P.; Saponari, M. Olea Europaea Geminivirus: A Novel Bipartite Geminivirid Infecting Olive Trees. Viruses 2021, 13, 481. [Google Scholar] [CrossRef]
- Fontana, A.; Piscopo, A.; De Bruno, A.; Tiberini, A.; Muzzalupo, I.; Albanese, G. Impact of olive leaf yellowing associated virus on olive (Olea europaea L.) oil. Eur. J. Lipid Sci. Technol. 2019, 121, 1800472. [Google Scholar] [CrossRef]
- Godena, S.; Bendini, A.; Giambanelli, E.; Cerretani, L.; Ðermić, D.; Ðermić, E. Cherry leafroll virus: Impact on olive fruit and virgin olive oil quality. Eur. J. Lipid Sci. Technol. 2012, 114, 535–541. [Google Scholar] [CrossRef]
- Erilmez, S.; Erkan, S. The Investigation about the effects of Strawberry latent ring spot virus on certain fruit quality characteristics of olive tree. J. Turk. Phytopathol. 2014, 43, 7–13. [Google Scholar]
- Hull, R. Comparative Plant Virology; Elsevier Academic Press: London, UK, 2009. [Google Scholar]
- Sabanadzovic, S.; Abou-Ghanem, N.; La Notte, P.; Savino, V.; Scarito, G.; Martelli, G.P. Partial molecular characterization and RT-PCR detection of a putative closterovirus associated with olive leaf yellowing. J. Plant Pathol. 1999, 81, 37–45. Available online: https://www.jstor.org/stable/41997939 (accessed on 1 February 2023).
- Ruiz-García, A.B.; Candresse, T.; Canales, C.; Morán, F.; Machado De Oliveira, C.; Bertolini, E.; Olmos, A. Molecular Characterization of the Complete Coding Sequence of Olive Leaf Yellowing-Associated Virus. Plants 2020, 9, 1272. [Google Scholar] [CrossRef]
- International Committee on Taxonomy of Viruses (ICTV). Available online: https://talk.ictvonline.org (accessed on 1 February 2023).
- Çağlayan, K.; Faggioli, F.; Barba, M. Viruses, phytoplasmas, and diseases of unknown etiology of olive trees. In Virus and Virus-Like Diseases of Pome and Stone Fruits; APS Publications: St. Paul, MN, USA, 2011; pp. 289–297. [Google Scholar] [CrossRef]
- Faggioli, F.; Ferretti, L.; Albanese, G.; Sciarroni, R.; Pasquini, G.; Lumia, V.; Barba, M. Distribution of olive tree viruses in Italy as revealed by one-step RT-PCR. J. Plant Pathol. 2005, 87, 45–51. [Google Scholar]
- Faggioli, F.; Roschetti, A.; Luigi, M.; Leo, G.; Albanese, G. Sanitary Selection of Olive (Olea europaea L.) Cultivars for Worldwide Distribution. Asian J. Plant Pathol. 2017, 11, 28–34. [Google Scholar] [CrossRef]
- Savino, V.; Sabanadzovic, S.; Scarito, G.; Laviola, C.; Martelli, G.P. Two olive yellows of possible viral origin in Sicily. Inf. Fitopatol. 1996, 46, 55–59. [Google Scholar]
- Saponari, M.; Alkowni, R.; Grieco, F.; Driouech, N.; Hassan, M.; Di Terlizzi, B.; Digiaro, M.; Pantaleo, V.; Savino, V.; Martelli, G.P. Detection of Olive-infecting virus in the Mediterranean basin. Acta Hortic. 2002, 586, 787–790. [Google Scholar] [CrossRef]
- Albanese, G.; Faggioli, F.; Ferretti, L.; Sciarroni, R.; La Rosa, R.; Barba, M. Sanitary status evaluation of olive cultivars in Calabria and Sicily. J. Plant Pathol. 2003, 85, 304. [Google Scholar]
- Martelli, G.P. Infectious diseases of olive. In Olive Diseases and Disorders; Schena, L., Agosteo, G.E., Cacciola, S.O., Eds.; Transworld Research Network: Kerala, India, 2011; pp. 71–88. [Google Scholar]
- Rowhani, A.; Al Rwahnih, M.; Golino, D.; Daubert, S.; Guo, Y. Characterization of latent viral infection of olive trees in the national clonal germplasm repository in California. J. Plant Pathol. 2011, 93, 227–231. [Google Scholar] [CrossRef]
- Afechtal, M.; Mounir, M. Preliminary evaluation of the status of olive-infecting viruses in Morocco. Moroc. J. Agric. Sci. 2020, 1, 1. [Google Scholar]
- El Air, M.; Mahfoudi, N.; Digiaro, M.; Najjar, A.; Elbeaino, T. Detection of olive-infecting viruses in Tunisia. J. Phytopathol. 2011, 159, 283–286. [Google Scholar] [CrossRef]
- Bjeliš, M.; Loconsole, G.; Saponari, M. Presence of viruses in Croatian olive groves. Pomol. Croat. 2007, 13, 165–172. [Google Scholar] [CrossRef]
- Fadel, C.; Digiaro, M.; Choueiri, E.; Elbeaino, T.; Saponari, M.; Savino, V.; Martelli, G.P. On the presence and distribution of olive viruses in Lebanon. EPPO Bull. 2005, 35, 33–36. [Google Scholar] [CrossRef]
- Al Abdullah, A.; Elbeaino, T.; Saponari, M.; Hallak, H.; Digiaro, M. Preliminary evaluation of the status of olive-infecting viruses in Syria. EPPO Bull. 2005, 35, 249–252. [Google Scholar] [CrossRef]
- Youssef, S.A.; Moawed, S.M.; El-Sayed, M.; Shalaby, A.A. Detection of olive tree viruses in Egypt by one-step RT-PCR. Julius-Kühn-Archiv. 2010, 427, 51. [Google Scholar]
- Mathioudakis, M.M.; Saponari, M.; Hasiów-Jaroszewska, B.; Elbeaino, T.; Koubouris, G. Detection of viruses in olive cultivars in Greece, using a rapid and efective RNA extraction method, for certification of virus-tested propagation material. Phytopathol. Mediterr. 2020, 59, 203–211. [Google Scholar] [CrossRef]
- Çakalli, A. Sanitary Assessment of Albanian Olive and Stone Fruit Varieties. Master’s Thesis, Università degli Studi di Bari Aldo Moro—CIHEAM (Istituto Agronomico Mediterraneo di Bari), Bari, Italy, 1999; p. 45. [Google Scholar]
- Luigi, M.; Manglli, A.; Thomaj, F.; Buonaurio, R.; Barba, M.; Faggioli, F. Phytosanitary evaluation of olive germplasm in Albania. Phytopathol. Mediterr. 2009, 48, 280–284. Available online: http://www.jstor.org/stable/26463352 (accessed on 1 February 2023).
- Samara, R.; Alkowni, R.; Qubbaj, T.; Abu-Qaoud, H.; Jarrar, S. Plant diseases associated with olive bark midge in West-Bank Palestine. Res. Crops 2018, 19, 712–719. [Google Scholar] [CrossRef]
- Marn, M.V.; Pleško, I.M. Detection of olive viruses in Slovenia. In Power of Viruses: Programme and Abstracts; Bielen, A., Ježić, M., Jurak, I., Škorić, D., Tomaic, V., Eds.; Hrvatsko Mikrobiološko Društvo: Zagabria, Croatia, 2018; p. 101. [Google Scholar]
- Ruiz-García, A.B.; Canales, C.; Morán, F.; Ruiz-Torres, M.; Herrera-Mármol, M.; Olmos, A. Characterization of Spanish Olive Virome by High Throughput Sequencing Opens New Insights and Uncertainties. Viruses 2021, 13, 2233. [Google Scholar] [CrossRef]
- Martelli, G.P. A Brief Outline of Infectious Diseases of Olive. Palest. Tech. Univ. Res. J. 2013, 1, 10. [Google Scholar] [CrossRef]
- Cutuli, M.; Lo Bianco, R.; Marra, F.P.; Caruso, T. Growth and physiological responses of young olive trees affected by Olive leaf yellowing associated virus. In International Symposium on Physiological Principles and Their Application to Fruit Production; Robinson, T., Ed.; ISHS Acta Horticulturae: Geneva, Switzerland; New York, NY, USA, 2014; pp. 165–168. [Google Scholar] [CrossRef]
- Martelli, G.P.; Prota, U. Malattie e patogeni dell’olivo trasmessi col materiale di propagazione. In I Georgofili, Quaderni, 2nd ed.; Polistampa: Firenze, Italy, 1997; pp. 73–96. [Google Scholar]
- Mollov, D.; Filloux, D.; Grinstead, S.; Bolus, S.; Claude, L.; Fernandez, E.; Julian, C.; Cadresse, T.; Lockhart, B.; Roumagnac, P.; et al. Sugarcane mild mosaic virus: The rediscovery of an identified but unrecognized virus. Plant Pathol. 2022, 72, 312–321. [Google Scholar] [CrossRef]
- GBIF—Global Biodiversity Information Facility. Available online: https://www.gbif.org/ (accessed on 1 June 2023).
- Guessab, A.; Lazreg, F.; Elouissi, M.; Elouissi, A.; Daikh, Z. Larvicidal activities of essential oils against Euphyllura olivina Costa (Homoptera: Psyllidae). An. Univ. Din Oradea Fasc. Biol. 2022, 2, 140–148. [Google Scholar]
- Martelli, G.P.; Savino, V.; Catalano, L.; Di Terlizzi, B.; Sabanadzovic, S.; Greco, N. Viruses and certification of olive in Apulia (Southern Italy). In XVI International Symposium on Fruit Tree Virus Diseases; Barba, M., Hadidi, A., Eds.; ISHS Acta Horticulturae: Rome, Italy, 1994; pp. 569–573. [Google Scholar] [CrossRef]
- Zellama, M.S.; Varanda, C.M.; Materatski, P.; Nabi, N.; Hafsa, A.B.; Saamali, B.M.; Félix, M.R. An integrated approach for understanding the high infection rates of olive viruses in Tunisia. Eur. J. Plant Pathol. 2019, 153, 1043–1054. [Google Scholar] [CrossRef]
- Erilmez, S. The comparison of sensitivity of various methods in the detection of olive tree viruses. J. Turk. Phytopathol. 2016, 45, 1–13. [Google Scholar]
- Clark, M.F.; Adams, A. Characteristics of the microplate method of enzyme-linked immunosorbent assay for the detection of plant viruses. J. Gen. Virol. 1977, 34, 475. [Google Scholar] [CrossRef] [PubMed]
- Ferriol, I.; Rangel, E.A.; Panno, S.; Davino, S.; Han, C.G.; Olmos, A.; Rubio, L. Rapid detection and discrimination of fabaviruses by flow-through hybridisation with genus-and species-specific riboprobes. Ann. Appl. Biol. 2015, 167, 26–35. [Google Scholar] [CrossRef]
- Panno, S.; Caruso, A.G.; Bertacca, S.; Mati’c, S.; Davino, S.; Parrella, G. Detection of parietaria mottle virus by RT-qPCR: An emerging virus native of Mediterranean area that undermine tomato and pepper production in Southern Italy. Front. Plant Sci. 2021, 12, 698573. [Google Scholar] [CrossRef]
- Campos, M.D.; Zellama, M.S.; Varanda, C.; Materatski, P.; Peixe, A.; Chaouachi, M.; Félix, M.D.R. Establishment of a sensitive qPCR methodology for detection of the olive-infecting viruses in Portuguese and Tunisian orchards. Front. Plant Sci. 2019, 10, 694. [Google Scholar] [CrossRef]
- Matić, S.; Candian, V.; D’Errico, C.; Pierro, R.; Panno, S.; Davino, S.; Noris, E.; Tedeschi, R. In-Field LAMP Detection of Flavescence Dorée Phytoplasma in Crude Extracts of the Scaphoideus titanus Vector. Agronomy 2022, 12, 1645. [Google Scholar] [CrossRef]
- Notomi, T.; Mori, Y.; Tomita, N.; Kanda, H. Loop-mediated isothermal amplification (LAMP): Principle, features, and future prospects. J. Microbiol. 2015, 53, 1–5. [Google Scholar] [CrossRef]
- Ayukawa, Y.; Hanyuda, S.; Fujita, N.; Komatsu, K.; Arie, T. Novel loop-mediated isothermal amplification (LAMP) assay with a universal QProbe can detect SNPs determining races in plant pathogenic fungi. Sci. Rep. 2017, 7, 4253. [Google Scholar] [CrossRef]
- Khan, M.; Wang, R.; Li, B.; Liu, P.; Weng, Q.; Chen, Q. Comparative evaluation of the LAMP assay and PCR-based assays for the rapid detection of Alternaria solani. Front. Microbiol. 2018, 9, 2089. [Google Scholar] [CrossRef]
- Panno, S.; Matić, S.; Tiberini, A.; Caruso, A.G.; Bella, P.; Torta, L.; Stassi, R.; Davino, S. Loop mediated isothermal amplification: Principles and applications in plant virology. Plants 2020, 9, 461. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef]
- Davino, S.; Panno, S.; Arrigo, M.; La Rocca, M.; Caruso, A.G.; Bosco, G.L. Planthology: An application system for plant diseases management. Chem. Eng. Trans. 2017, 58, 619–624. [Google Scholar] [CrossRef]
- Gottwald, T.R.; Hughes, G. A new survey method for Citrus tristeza virus disease assessment. In Proceedings of the XIV International Organization of Citrus Virologists (1957–2010), Campinas, Brazil, 19–18 September 1998; Volume 14, pp. 77–87. [Google Scholar] [CrossRef]
- Zhao, J.; Zhang, X.; Hong, Y.; Liu, Y. Chloroplast in plant-virus interaction. Front. Microbiol. 2016, 7, 1565. [Google Scholar] [CrossRef]
- Albanese, G.; Saponari, M.; Faggioli, F. Phytosanitary certification. In Olive Germoplasm—The Olive Cultivation, Table Olive and Olive Oil Industry in Italy; Muzzalupo, I., Ed.; IntechOpen: London, UK, 2012; pp. 107–132. [Google Scholar] [CrossRef]
- Martelli, G.P.; Salerno, M.; Savino, V.; Prota, U. An appraisal of diseases and pathogens of olive. In IV International Symposium on Olive Growing; Vitagliano, C., Martelli, G.P., Eds.; ISHS Acta Horticulturae: Valenzano, Italy, 2000; pp. 701–708. [Google Scholar] [CrossRef]
- Montilon, V.; Potere, O.; Susca, L.; Bottalico, G. Phytosanitary Rules for the Movement of Olive (Olea europaea L.) Propagation Material into the European Union (EU). Plants 2023, 12, 699. [Google Scholar] [CrossRef]
- Kiddle, G.; Hardinge, P.; Buttigieg, N.; Gandelman, O.; Pereira, C.; McElgunn, C.J.; Rizzoli, M.; Jackson, R.; Appleton, N.; Moore, C.; et al. GMO detection using a bioluminescent real time reporter (BART) of loop mediated isothermal amplification (LAMP) suitable for field use. BMC Biotechnol. 2012, 12, 15. [Google Scholar] [CrossRef]
- Schrader, C.; Schielke, A.; Ellerbroek, L.; Johne, R. PCR inhibitors–occurrence, properties and removal. J. Appl. Microbiol 2012, 113, 1014–1026. [Google Scholar] [CrossRef]
- Zhao, L.; Li, G.; Gao, Y.; Zhu, Y.; Liu, J.; Zhu, X. Reverse transcription loop-mediated isothermal amplification assay for detecting tomato chlorosis virus. J. Virol. Methods 2015, 213, 93–97. [Google Scholar] [CrossRef]
- Zhang, Y.; Xie, Z.; Fletcher, J.D.; Wang, Y.; Wang, R.; Guo, Z.; He, Y. Rapid and sensitive detection of Lettuce yellows and cucumber mosaic virus infecting lettuce (Lactuca sativa L.) by reverse transcription loop-mediated isothermal amplification. Plant Pathol. J. 2020, 36, 76–86. [Google Scholar] [CrossRef]
- Tiberini, A.; Tomlinson, J.; Micali, G.; Fontana, A.; Albanese, G.; Tomassoli, L. Development of a reverse transcription-loop-mediated isothermal amplification (LAMP) assay for the rapid detection of Onion yellow dwarf virus. J. Virol. Methods 2019, 271, 113680. [Google Scholar] [CrossRef]
- Caruso, A.G.; Ragona, A.; Bertacca, S.; Montoya, M.A.M.; Panno, S.; Davino, S. Development of an In-Field Real-Time LAMP Assay for Rapid Detection of Tomato Leaf Curl New Delhi Virus. Plants 2023, 12, 1487. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Genus | Family | GenBank Accession No. |
|---|---|---|---|
| Citrus tristeza virus (CTV) | Closterovirus | Closteroviridae | EU937521 AF001623 Y18420 |
| Grapevine leafroll-associated virus 1 (GLRaV-1) | NC016509 | ||
| Grapevine leafroll-associated virus 2 (GLRaV-2) | Ampelovirus | DQ286725 | |
| Grapevine leafroll-associated virus 3 (GLRaV-3) | AF037268 | ||
| Tomato chlorosis virus (ToCV) | Crinivirus | RNA1: AY903447 RNA2: AY903448 | |
| Tomato infectious chlorosis virus (TICV) | RNA1: FJ815440 RNA2: FJ815441 | ||
| Cucurbit yellow stunting disorder virus (CYSDV) | RNA1: NC004809 RNA2: NC004810 | ||
| Actinidia virus 1 (AcV1) | Olivavirus | KX857665 | |
| Persimmon virus B (PeVB) | AB923924 |
| Country | Region | Province | No. of Samples Collected | Cultivar |
|---|---|---|---|---|
| Italy | Sicily | Agrigento | 20 | Giarraffa |
| Trapani | 20 | Nocellara del Belice | ||
| Calabria | Crotone | 20 | Carolea | |
| Apulia | Taranto | 20 | Ogliarola barese | |
| Lazio | Frosinone | 20 | Leccino | |
| Umbria | Terni | 20 | Frantoio | |
| Spain | Comunitat Valenciana | Valencia | 20 | Serrana Espadán, Picual |
| Castellón | 20 | Serrana Espadán, Villalonga | ||
| Andalusia | Jaén | 20 | Picual, Villalonga | |
| Granada | 20 | Arbequina |
| Primer Name | Binding Sites (Genbank Acc. No. MW056495) | Genomic Position | Nucleotide Sequence (5′-3′) | Annealing Temperature—Ta (°C) | Amplicon Size (bp) |
|---|---|---|---|---|---|
| OLYaV-21F | 21–44 | 5′ UTR-ORF1a | ATCAATTGAAGAAAACCACTCCC | 59 | 450 |
| OLYaV-471R | 450–471 | AGTACCTCCCACGACGTATTG | |||
| OLYaV-30F | 30–54 | 5′ UTR-ORF1a | GAAAACCACTCCCTTCAATTCAAT | 61 | 497 |
| OLYaV-527R | 505–527 | GAAGAACTATTGATTGGCTTGGG | |||
| OLYaV-36F | 36–59 | 5′ UTR-ORF1a | CCACTCCCTTCAATTCAATAACAT | 60 | 428 |
| OLYaV-444R | 444–464 | CCCACGACGTATTGACCACTC | |||
| OLYaV-46F | 46–67 | 5′ UTR-ORF1a | CAATTCAATAACATAACATACC | 60 | 345 |
| OLYaV-371R | 371–391 | ATGTGACTTTTGACTGAGGTA | |||
| OLYaV-495F | 495–517 | ORF1a | GAAGAACACTCCCAAGCCAATC | 60 | 646 |
| OLYaV-1141R | 1120–1141 | CACACTCTCTCTTGTAAGTCCC | |||
| OLYaV-742F | 742–763 | ORF1a | GCGGCTTGGGTCATTAAGGGT | 60 | 427 |
| OLYaV-1166R | 1147–1166 | GGGCAAAAATAGTGAGAGCGAC |
| Primer Name | Nucleotide Sequence (5′-3′) | Amplicon Size (bp) |
|---|---|---|
| OLYaV-F3 | CTGGTCTCTACGTACAGGGA | 195 |
| OLYaV-B3 | GGTACTGCCTCAGTTCCCA | |
| OLYaV-FIP | CCGGTAACGACCTTCAGCCTTCCCCGCTGTGCAGAACAAC | - |
| OLYaV-BIP | AAGAAAACGGGCTGTAGCCCATGCCGTCAATGTTACGAGC | |
| OLYaV-LoopF | TCTGTGCAGATCTAGATTTGGGA | - |
| OLYaV-LoopB | GAGGAAAGCAGCGACCACC |
| Sample | Real-Time RT-LAMP Reaction Time (Min) | ||
|---|---|---|---|
| Assay #1 | Assay #2 | Assay #3 | |
| OLYaV V64 isolate | 13.7 | 14.1 | 14.4 |
| Negative control (NC) | - | - | - |
| Starting DNA Concentration (50 ng/μL) (1.34 × 1010 Copies) | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Assay | 100 | 10−1 | 10−2 | 10−3 | 10−4 | 10−5 | 10−6 | 10−7 | 10−8 | 10−9 | 10−10 | 10−11 | 10−12 | 10−13 | 10−14 | 10−15 |
| End-point RT-PCR | + | + | + | + | + | + | + | + | + | + | + | + | − | − | − | − |
| LAMP reaction time plateau (min) (mean values ± SD) | 13 ± 0.1 | 13.9 ± 0.2 | 15.2 ± 0.2 | 16.8 ± 0.2 | 17.4 ± 0.1 | 20.1 ± 0.4 | 22 ± 0.2 | 24.4 ± 0.4 | 25.1 ± 0.3 | 26.3 ± 0.4 | 31.2 ± 0.4 | 35.1 ± 0.3 | 43.4 ± 0.4 | − | − | − |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Caruso, A.G.; Bertacca, S.; Ragona, A.; Agrò, G.; Font-San-Ambrosio, M.I.; Alfaro-Fernández, A.; Sánchez, R.E.; Panno, S.; Davino, S. Detection by Sensitive Real-Time Reverse Transcription Loop-Mediated Isothermal Amplification of Olive Leaf Yellowing Associated Virus and Its Incidence in Italy and Spain. Horticulturae 2023, 9, 702. https://doi.org/10.3390/horticulturae9060702
Caruso AG, Bertacca S, Ragona A, Agrò G, Font-San-Ambrosio MI, Alfaro-Fernández A, Sánchez RE, Panno S, Davino S. Detection by Sensitive Real-Time Reverse Transcription Loop-Mediated Isothermal Amplification of Olive Leaf Yellowing Associated Virus and Its Incidence in Italy and Spain. Horticulturae. 2023; 9(6):702. https://doi.org/10.3390/horticulturae9060702
Chicago/Turabian StyleCaruso, Andrea Giovanni, Sofia Bertacca, Arianna Ragona, Graziella Agrò, Maria Isabel Font-San-Ambrosio, Ana Alfaro-Fernández, Rocío Estévez Sánchez, Stefano Panno, and Salvatore Davino. 2023. "Detection by Sensitive Real-Time Reverse Transcription Loop-Mediated Isothermal Amplification of Olive Leaf Yellowing Associated Virus and Its Incidence in Italy and Spain" Horticulturae 9, no. 6: 702. https://doi.org/10.3390/horticulturae9060702
APA StyleCaruso, A. G., Bertacca, S., Ragona, A., Agrò, G., Font-San-Ambrosio, M. I., Alfaro-Fernández, A., Sánchez, R. E., Panno, S., & Davino, S. (2023). Detection by Sensitive Real-Time Reverse Transcription Loop-Mediated Isothermal Amplification of Olive Leaf Yellowing Associated Virus and Its Incidence in Italy and Spain. Horticulturae, 9(6), 702. https://doi.org/10.3390/horticulturae9060702