
Medium/Long-Term Efficiency of Struvite for Lettuce (Lactuca sativa L.) Production: Effect on Soil Quality

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Plant Analysis
2.3. Phosphorus Uptake (PU) and Relative Agronomic Efficiency (RAE)
2.4. Soil Physico-Chemical Analysis
2.5. Soil Biological Analysis
2.5.1. Enzyme Activity
2.5.2. Substrate-Induced Respiration (SIR)
2.5.3. Community-Level Physiological Profiling (CLPP) Analysis
2.6. Statistical Analysis
3. Results
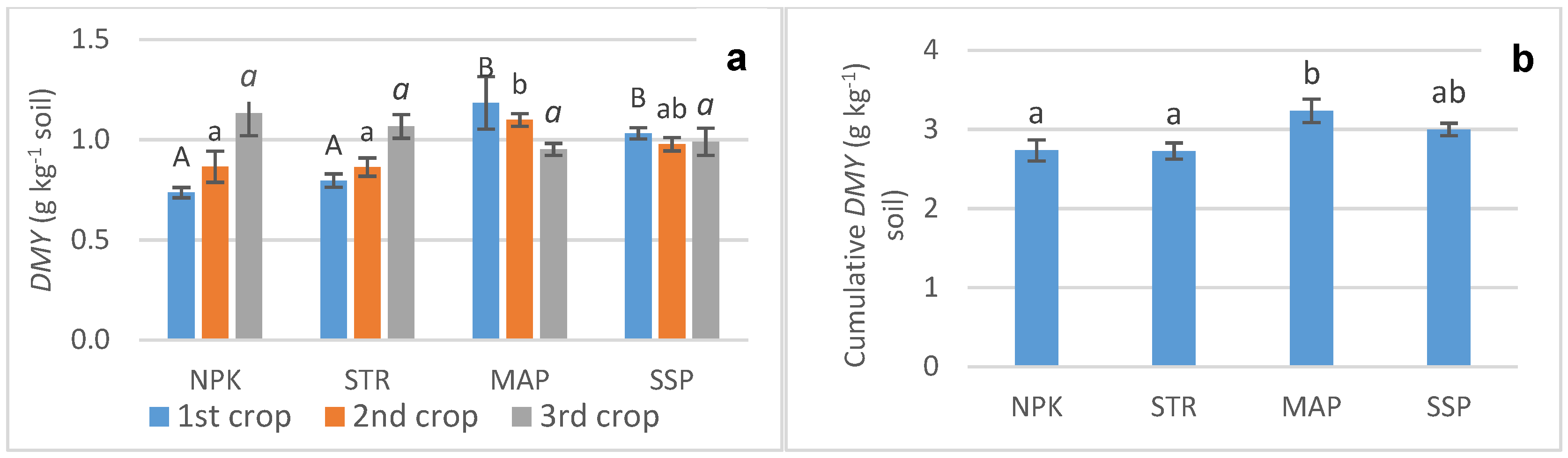
3.1. Biomass Production (DMY)
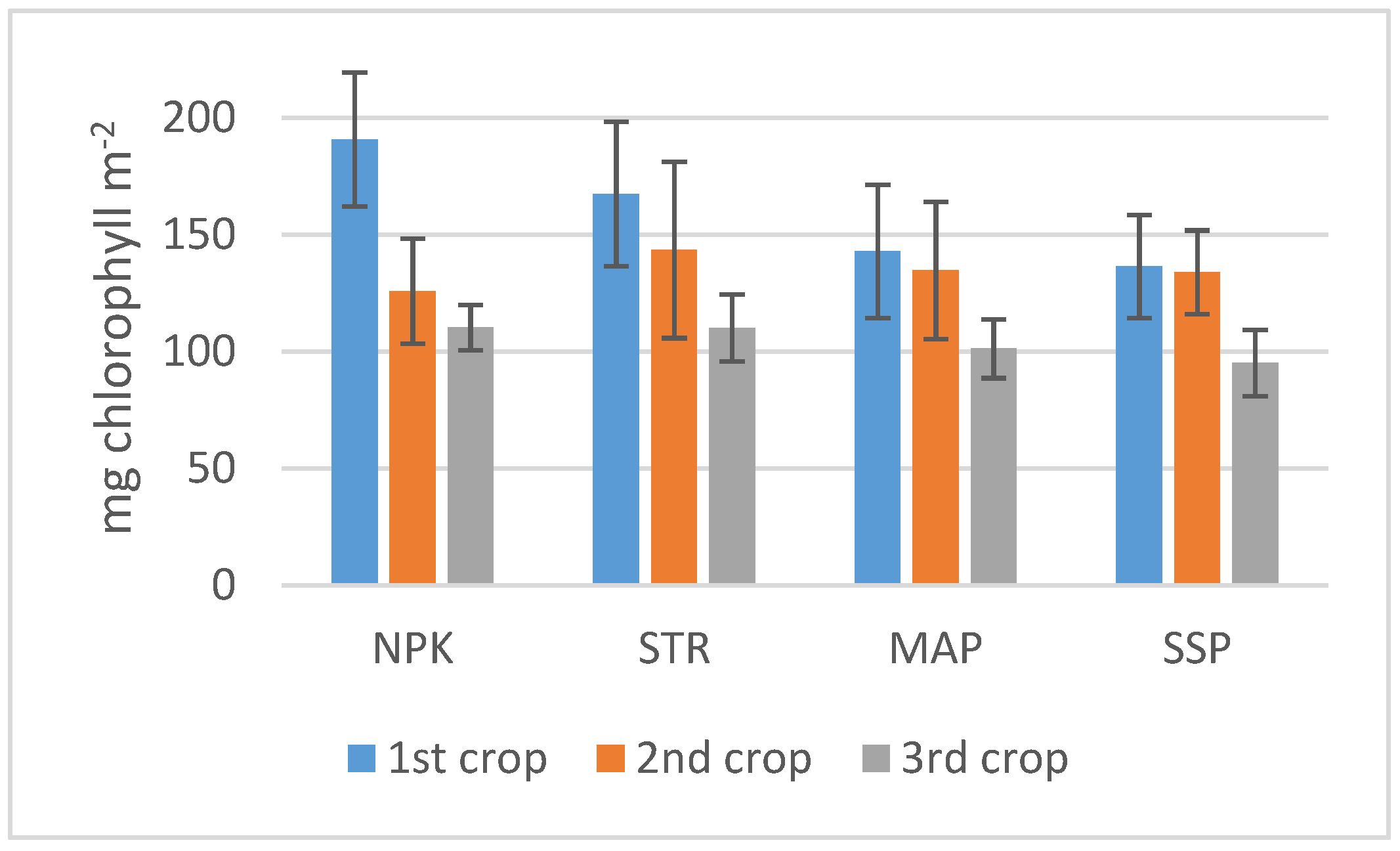
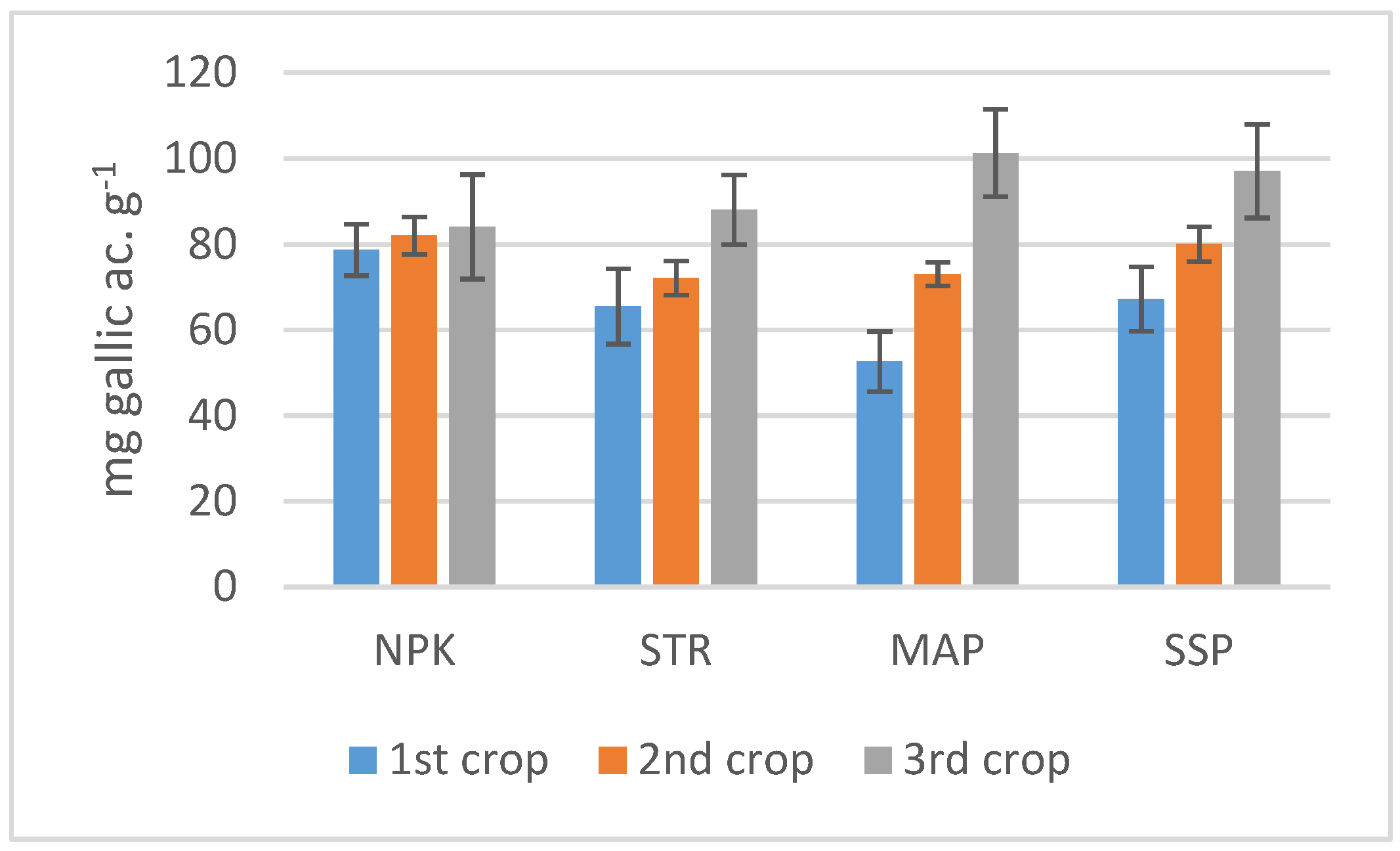
3.2. Plant Analysis
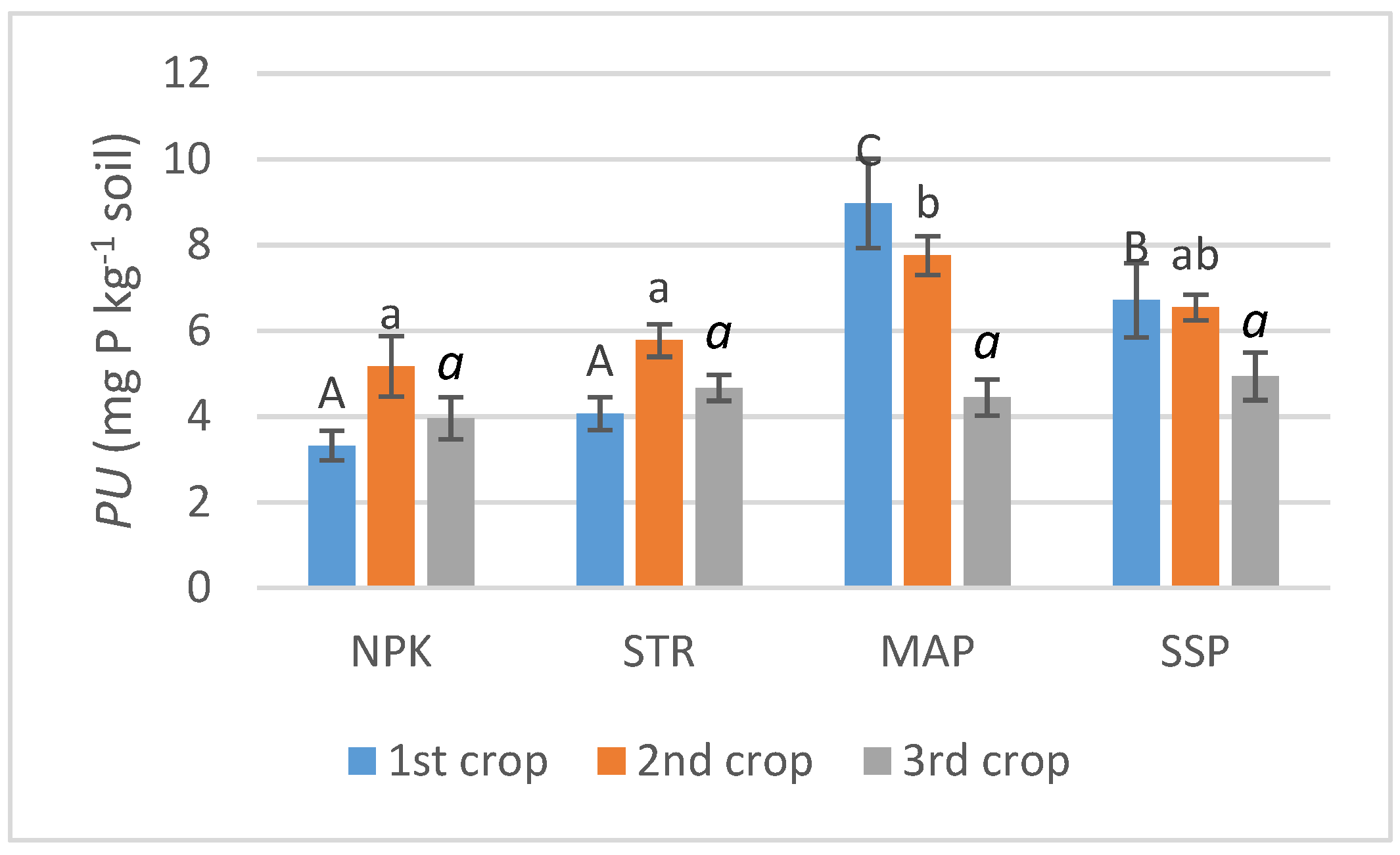
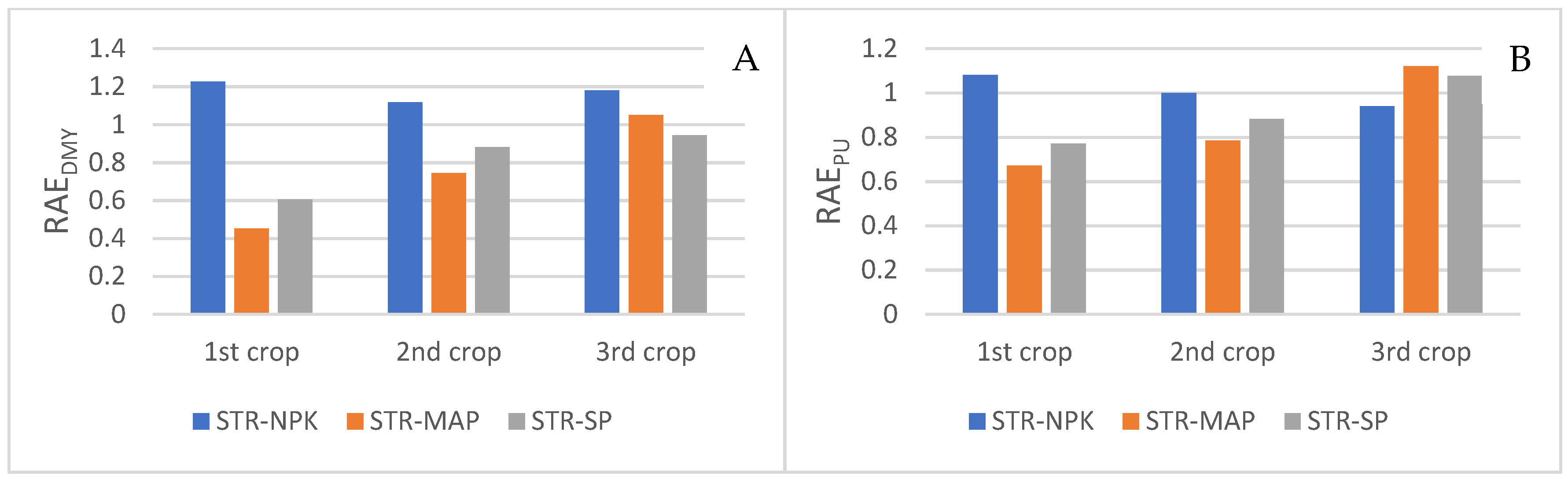
3.3. Phosphorus Uptake (PU) and Relative Agronomic Efficiency (RAE)
3.4. Effect on Soil Properties
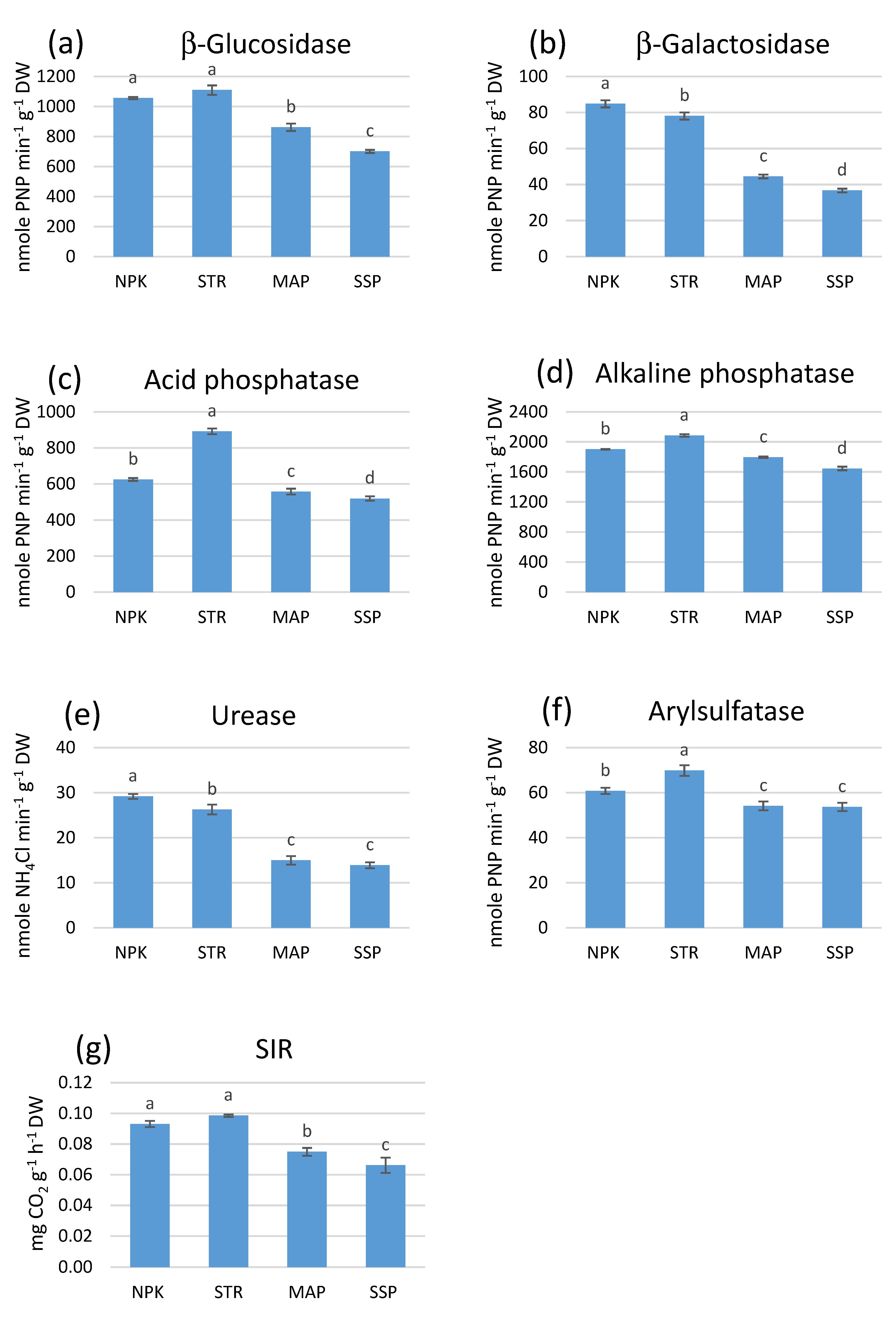
3.5. Effect of Phosphorus Fertilizers on Soil Enzyme Activities and Substrate-Induced Respiration (SIR)
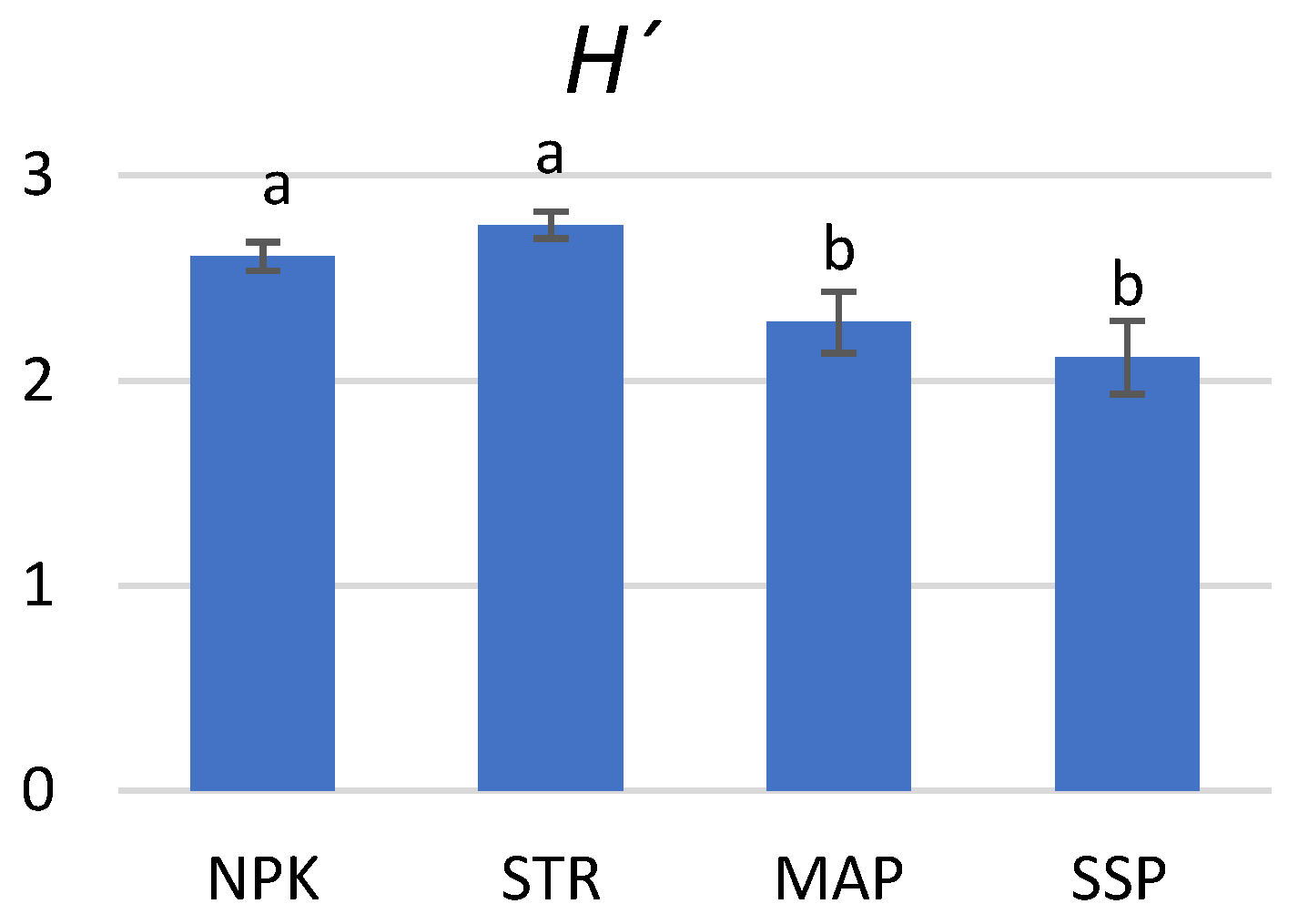
3.6. Effect on Community-Level Physiological Profiles
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- United Nations. World Population Prospects: The 2017 Revision, Key Findings and Advance Tables; Working Paper No. ESA/P/WP/248; United Nations: New York, NY, USA, 2017.
- Le Mouël, C.; Forslund, A. How Can We Feed the World in 2050? A Review of the Responses from Global Scenario Studies. Eur. Rev. Agric. Econ. 2017, 44, 541–591. [Google Scholar] [CrossRef]
- Tilman, D.; Balzer, C.; Hill, J.; Befort, B.L. Global Food Demand and the Sustainable Intensification of Agriculture. Proc. Natl. Acad. Sci. USA 2011, 108, 20260–20264. [Google Scholar] [CrossRef] [PubMed]
- Barker, A.V.; Pilbeam, D.J. Handbook of Plant Nutrition, 2nd ed.; Press, C., Ed.; CRC Press: Boca Raton, FL, USA, 2015. [Google Scholar]
- Dodds, W.K.; Bouska, W.W.; Eitzmann, J.L.; Pilger, T.J.; Pitts, K.L.; Riley, A.J.; Schloesser, J.T.; Thornbrugh, D.J. Eutrophication of U. S. Freshwaters: Analysis of Potential Economic Damages. Environ. Sci. Technol. 2009, 43, 12–19. [Google Scholar] [CrossRef]
- Withers, P.J.A.; Neal, C.; Jarvie, H.P.; Doody, D.G. Agriculture and Eutrophication: Where Do We Go from Here? Sustainability. 2014, 6, 5853–5875. [Google Scholar] [CrossRef]
- Savci, S. An Agricultural Pollutant: Chemical Fertilizer. Int. J. Environ. Sci. Dev. 2012, 3, 73–80. [Google Scholar] [CrossRef]
- Reijnders, L. Phosphorus Resources, Their Depletion and Conservation, a Review. Resour. Conserv. Recycl. 2014, 93, 32–49. [Google Scholar] [CrossRef]
- Cordell, D.; Neset, T.S.S. Phosphorus Vulnerability: A Qualitative Framework for Assessing the Vulnerability of National and Regional Food Systems to the Multi-Dimensional Stressors of Phosphorus Scarcity. Glob. Environ. Chang. 2014, 24, 108–122. [Google Scholar] [CrossRef]
- Elser, J.J. Phosphorus: A Limiting Nutrient for Humanity? Curr. Opin. Biotechnol. 2012, 23, 833–838. [Google Scholar] [CrossRef]
- European Commission. Critical Raw Materials Resilience: Charting a Path towards Greater Security and Sustainability. In Communication from the Commission to the European Parliament, the Council, the European Economic and Social Committee and the Committee of the Regions; European Commission: Brussels, Belgium, 2020. [Google Scholar]
- Cordell, D.; White, S. Peak Phosphorus: Clarifying the Key Issues of a Vigorous Debate about Long-Term Phosphorus Security. Sustainability 2011, 3, 2027–2049. [Google Scholar] [CrossRef]
- Heckenmüller, M.; Narita, D.; Klepper, G. Global Availability of Phosphorus and Its Implications for Global Food Supply: An Economic Overview; Kiel Working Paper, No. 1897; Kiel Institute for the World Economy (IfW): Kiel, Germany, 2014. [Google Scholar]
- Cordell, D.; Drangert, J.O.; White, S. The Story of Phosphorus: Global Food Security and Food for Thought. Glob. Environ. Chang. 2009, 19, 292–305. [Google Scholar] [CrossRef]
- Smil, V. Phosphorus in the Environment: Natural Flows and Human Interferences. Annu. Rev. Energy Environ. 2000, 25, 53–88. [Google Scholar] [CrossRef]
- European Fertilizer Manufacturers Association. Phosphorus: Essential Element for Food Production; European Fertilizer Manufacturers Association (EFMA): Brussels, Belgium, 2000; pp. 1–38. [Google Scholar]
- Chojnacka, K.; Moustakas, K.; Witek-Krowiak, A. Bio-Based Fertilizers: A Practical Approach towards Circular Economy. Bioresour. Technol. 2020, 295, 122223. [Google Scholar] [CrossRef]
- De-Bashan, L.E.; Bashan, Y. Recent Advances in Removing Phosphorus from Wastewater and Its Future Use as Fertilizer (1997–2003). Water Res. 2004, 38, 4222–4246. [Google Scholar] [CrossRef] [PubMed]
- Shu, L.; Schneider, P.; Jegatheesan, V.; Johnson, J. An Economic Evaluation of Phosphorus Recovery as Struvite from Digester Supernatant. Bioresour. Technol. 2006, 97, 2211–2216. [Google Scholar] [CrossRef] [PubMed]
- Barnard, J.L. Elimination of Eutrophication through Resource Recovery. In International Conference on Nutrient Recovery from Wastewater Streams; IWA Publishing: Vancouver, BC, Canada, 2009; ISBN 9781843392323. [Google Scholar]
- Qiao, J.; Li, X.; Li, F.; Liu, T.; Young, L.Y.; Huang, W.; Sun, K.; Tong, H.; Hu, M. Humic Substances Facilitate Arsenic Reduction and Release in Flooded Paddy Soil. Environ. Sci. Technol. 2019, 53, 5034–5042. [Google Scholar] [CrossRef]
- European Commission. Directive 2000/60/EC of the European Parliament and of the Council of 23 October 2000 Establishing a Framework for the Community Action in the Field of Water Policy; European Commission: Brussels, Belgium, 2000.
- European Commission. Directive 91/271/EEC of the European Council of 21 May 1991 Concerning Urban Waste Water Treatment; European Commission: Brussels, Belgium, 1991; pp. 40–52.
- Gilbert, N. The Disappearing Nutrient. Nature 2009, 461, 716–718. [Google Scholar] [CrossRef] [PubMed]
- European Commission. A New Circular Economy Action Plan for a Cleaner and More Competitive Europe. In Communication from the Commission to the European Parliament, the Council, the European Economic and Social Committee and the Committee of the Regions; European Commission: Brussels, Belgium, 2020. [Google Scholar]
- European Commission. Commission Delegated Regulation (EU) 2021/2086 of 5 July 2021 Amending Annexes II and IV to Regulation (EU) 2019/1009 of the European Parliament and of the Council for the Purpose of Adding Precipitated Phosphate Salts and Derivates as a Component Materia; European Commission: Brussels, Belgium, 2021.
- Huygens, D.; Saveyn, H.G.M.; Tonini, D.; Eder, P.; Delgado Sancho, L. Technical Proposals for Selected New Fertilising Materials under the Fertilising Products Regulation (Regulation (EU) 2019/1009); European Commission: Brussels, Belgium, 2019; ISBN 9789276098881.
- Kataki, S.; West, H.; Clarke, M.; Baruah, D.C. Phosphorus Recovery as Struvite: Recent Concerns for Use of Seed, Alternative Mg Source, Nitrogen Conservation and Fertilizer Potential. Resour. Conserv. Recycl. 2016, 107, 142–156. [Google Scholar] [CrossRef]
- Maaß, O.; Grundmann, P.; Von Bock Und Polach, C. Added-Value from Innovative Value Chains by Establishing Nutrient Cycles via Struvite. Resour. Conserv. Recycl. 2014, 87, 126–136. [Google Scholar] [CrossRef]
- Munir, M.T.; Li, B.; Boiarkina, I.; Baroutian, S.; Yu, W.; Young, B.R. Phosphate Recovery from Hydrothermally Treated Sewage Sludge Using Struvite Precipitation. Bioresour. Technol. 2017, 239, 171–179. [Google Scholar] [CrossRef]
- Rahman, M.M.; Salleh, M.A.M.; Rashid, U.; Ahsan, A.; Hossain, M.M.; Ra, C.S. Production of Slow Release Crystal Fertilizer from Wastewaters through Struvite Crystallization—A Review. Arab. J. Chem. 2014, 7, 139–155. [Google Scholar] [CrossRef]
- Mancho, C.; Diez-Pascual, S.; Alonso, J.; Gil-Díaz, M.; Lobo, M.C. Assessment of Recovered Struvite as a Safe and Sustainable Phosphorous Fertilizer. Environments 2023, 10, 22. [Google Scholar] [CrossRef]
- Li, X.Z.; Zhao, Q.L. Recovery of Ammonium-Nitrogen from Landfill Leachate as a Multi-Nutrient Fertilizer. Ecol. Eng. 2003, 20, 171–181. [Google Scholar] [CrossRef]
- Negrea, A.; Lupa, L.; Negrea, P.; Ciopec, M.; Muntean, C. Simultaneous Removal of Ammonium and Phosphate Ions from Wastewaters and Characterization of the Resulting Product. Chem. Bull. “POLITEHNICA” Univ. 2010, 55, 136–142. [Google Scholar]
- El Rafie, S.; Hawash, S.; Shalaby, M.S. Evaluation of Struvite Precipitated from Chemical Fertilizer Industrial Effluents. Adv. Appl. Sci. Res. 2013, 4, 113–123. [Google Scholar]
- Gell, K.; van Groenigen, J.W.; Cayuela, M.L. Residues of Bioenergy Production Chains as Soil Amendments: Immediate and Temporal Phytotoxicity. J. Hazard. Mater. 2011, 186, 2017–2025. [Google Scholar] [CrossRef] [PubMed]
- Hertzberger, A.J.; Cusick, R.D.; Margenot, A.J. A Review and Meta-Analysis of the Agricultural Potential of Struvite as a Phosphorus Fertilizer. Soil Sci. Soc. Am. J. 2020, 84, 653–671. [Google Scholar] [CrossRef]
- Robles-Aguilar, A.A.; Schrey, S.D.; Postma, J.A.; Temperton, V.M.; Jablonowski, N.D. Phosphorus Uptake from Struvite Is Modulated by the Nitrogen Form Applied. J. Plant Nutr. Soil Sci. 2020, 183, 80–90. [Google Scholar] [CrossRef]
- Plaza, C.; Sanz, R.; Clemente, C.; Fernández, J.M.; González, R.; Polo, A.; Colmenarejo, M.F. Greenhouse Evaluation of Struvite and Sludges from Municipal Wastewater Treatment Works as Phosphorus Sources for Plants. J. Agric. Food Chem. 2007, 55, 8206–8212. [Google Scholar] [CrossRef]
- Ponce, R.; Sa, M.E. Efficacy of Magnesium Ammonium Phosphate Recovered from Wastewater on White Lupin Plant. A Greenhouse Experiment. Agrochimica 2008, 52, 352–359. [Google Scholar]
- Ricardo, G.P.; López-de-Sá, E.G.; Plaza, C. Lettuce Response to Phosphorus Fertilization with Struvite Recovered from Municipal Wastewater. HortScience 2009, 44, 426–430. [Google Scholar] [CrossRef]
- Ryu, H.D.; Lim, C.S.; Kim, Y.K.; Kim, K.Y.; Lee, S.I. Recovery of Struvite Obtained from Semiconductor Wastewater and Reuse as a Slow-Release Fertilizer. Environ. Eng. Sci. 2012, 29, 540–548. [Google Scholar] [CrossRef]
- Wen, G.; Huang, L.; Zhang, X.; Hu, Z. Uptake of Nutrients and Heavy Metals in Struvite Recovered from a Mixed Wastewater of Human Urine and Municipal Sewage by Two Vegetables in Calcareous Soil. Environ. Technol. Innov. 2019, 15, 100384. [Google Scholar] [CrossRef]
- Antonini, S.; Arias, M.A.; Eichert, T.; Clemens, J. Greenhouse Evaluation and Environmental Impact Assessment of Different Urine-Derived Struvite Fertilizers as Phosphorus Sources for Plants. Chemosphere 2012, 89, 1202–1210. [Google Scholar] [CrossRef] [PubMed]
- Gell, K.; de Ruijter, F.J.; Kuntke, P.; de Graaff, M.; Smit, A.L. Safety and Effectiveness of Struvite from Black Water and Urine as a Phosphorus Fertilizer. J. Agric. Sci. 2011, 3, 67. [Google Scholar] [CrossRef]
- Nongqwenga, N.; Muchaonyerwa, P.; Hughes, J.; Odindo, A.; Bame, I. Possible Use of Struvite as an Alternative Phosphate Fertilizer. J. Soil Sci. Plant Nutr. 2017, 17, 581–593. [Google Scholar] [CrossRef]
- Ryu, H.D.; Lim, C.S.; Kang, M.K.; Lee, S.I. Evaluation of Struvite Obtained from Semiconductor Wastewater as a Fertilizer in Cultivating Chinese Cabbage. J. Hazard. Mater. 2012, 221–222, 248–255. [Google Scholar] [CrossRef]
- Bastida, F.; Jehmlich, N.; Martínez-Navarro, J.; Bayona, V.; García, C.; Moreno, J.L. The Effects of Struvite and Sewage Sludge on Plant Yield and the Microbial Community of a Semiarid Mediterranean Soil. Geoderma 2019, 337, 1051–1057. [Google Scholar] [CrossRef]
- Degryse, F.; Baird, R.; da Silva, R.C.; McLaughlin, M.J. Dissolution Rate and Agronomic Effectiveness of Struvite Fertilizers—Effect of Soil PH, Granulation and Base Excess. Plant Soil 2017, 410, 139–152. [Google Scholar] [CrossRef]
- Talboys, P.J.; Heppell, J.; Roose, T.; Healey, J.R.; Jones, D.L.; Withers, P.J.A. Struvite: A Slow-Release Fertiliser for Sustainable Phosphorus Management? Plant Soil 2016, 401, 109–123. [Google Scholar] [CrossRef]
- Li, B.; Boiarkina, I.; Yu, W.; Huang, H.M.; Munir, T.; Wang, G.Q.; Young, B.R. Phosphorous Recovery through Struvite Crystallization: Challenges for Future Design. Sci. Total Environ. 2019, 648, 1244–1256. [Google Scholar] [CrossRef]
- Arcas-Pilz, V.; Rufí-Salís, M.; Parada, F.; Petit-Boix, A.; Gabarrell, X.; Villalba, G. Recovered Phosphorus for a More Resilient Urban Agriculture: Assessment of the Fertilizer Potential of Struvite in Hydroponics. Sci. Total Environ. 2021, 799, 149424. [Google Scholar] [CrossRef] [PubMed]
- Robles-Aguilar, A.A.; Grunert, O.; Hernandez-Sanabria, E.; Mysara, M.; Meers, E.; Boon, N.; Jablonowski, N.D. Effect of Applying Struvite and Organic N as Recovered Fertilizers on the Rhizosphere Dynamics and Cultivation of Lupine (Lupinus angustifolius). Front. Plant Sci. 2020, 11, 572741. [Google Scholar] [CrossRef]
- Karpinska, A.; Ryan, D.; Germaine, K.; Dowling, D.; Forrestal, P.; Kakouli-Duarte, T. Soil Microbial and Nematode Community Response to the Field Application of Recycled Bio-Based Fertilisers in Irish Grassland. Sustainability 2021, 13, 12342. [Google Scholar] [CrossRef]
- Verstraete, W.; Mertens, B. Chapter 5. The Key Role of Soil Microbes. In Vital Soil; Doelman, P., Eijsackers, H.J.P., Eds.; Developments in Soil Science; Elsevier: Amsterdam, The Netherlands, 2004; Volume 29, pp. 127–157. [Google Scholar]
- Alkorta, I.; Aizpurua, A.; Riga, P.; Albizu, I.; Amézaga, I.; Garbisu, C. Soil Enzyme Activities as Biological Indicators of Soil Health. Rev. Environ. Health 2003, 18, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Bowles, T.M.; Acosta-Martínez, V.; Calderón, F.; Jackson, L.E. Soil Enzyme Activities, Microbial Communities, and Carbon and Nitrogen Availability in Organic Agroecosystems across an Intensively-Managed Agricultural Landscape. Soil Biol. Biochem. 2014, 68, 252–262. [Google Scholar] [CrossRef]
- Nannipieri, P.; Kandeler, E.; Ruggiero, P. Enzyme Activities and Microbiological and Biochemical Processes in Soil. In Enzymes in the Environment; CRC Press: Boca Raton, FL, USA, 2002; pp. 1–34. [Google Scholar]
- Bending, G.D.; Turner, M.K.; Rayns, F.; Marx, M.C.; Wood, M. Microbial and Biochemical Soil Quality Indicators and Their Potential for Differentiating Areas under Contrasting Agricultural Management Regimes. Soil Biol. Biochem. 2004, 36, 1785–1792. [Google Scholar] [CrossRef]
- Sowerby, A.; Emmett, B.; Beier, C.; Tietema, A.; Peñuelas, J.; Estiarte, M.; Van Meeteren, M.J.M.; Hughes, S.; Freeman, C. Microbial Community Changes in Heathland Soil Communities along a Geographical Gradient: Interaction with Climate Change Manipulations. Soil Biol. Biochem. 2005, 37, 1805–1813. [Google Scholar] [CrossRef]
- Rutgers, M.; Schouten, A.J.; Bloem, J.; Van Eekeren, N.; De Goede, R.G.M.; Jagers Op Akkerhuis, G.A.J.M.; Van Der Wal, A.; Mulder, C.; Brussaard, L.; Breure, A.M. Biological Measurements in a Nationwide Soil Monitoring Network. Eur. J. Soil Sci. 2009, 60, 820–832. [Google Scholar] [CrossRef]
- Rutgers, M.; Wouterse, M.; Drost, S.M.; Breure, A.M.; Mulder, C.; Stone, D.; Creamer, R.E.; Winding, A.; Bloem, J. Monitoring Soil Bacteria with Community-Level Physiological Profiles Using BiologTM ECO-Plates in The Netherlands and Europe. Appl. Soil Ecol. 2016, 97, 23–35. [Google Scholar] [CrossRef]
- Zhao, F.; McGrath, S.P.; Crosland, A.R. Comparison of Three Wet Digestion Methods for the Determination of Plant Sulphur by Inductively Coupled Plasma Atomic Emission Spectroscopy (ICP-AES). Commun. Soil Sci. Plant Anal. 1994, 25, 407–418. [Google Scholar] [CrossRef]
- Inskeep, W.P.; Bloom, P.R. Extinction Coefficients of Chlorophyll a and b in N,N-Dimethylformamide and 80% Acetone. Plant Physiol. 1985, 77, 483–485. [Google Scholar] [CrossRef] [PubMed]
- Ainsworth, E.A.; Gillespie, K.M. Estimation of Total Phenolic Content and Other Oxidation Substrates in Plant Tissues Using Folin-Ciocalteu Reagent. Nat. Protoc. 2007, 2, 875–877. [Google Scholar] [CrossRef] [PubMed]
- MAPA. Métodos Oficiales de Análisis; Secretaria General Técnica, Ministerio de Agricultura, Pesca y Alimentación: Madrid, Spain, 1994; Volume III, pp. 219–324. [Google Scholar]
- Walkley, A.; Black, I.A. An examination of Degtjareff method for determining soil organic matter and a proposed modification of the chromic acid titration method. Soil Sci. 1934, 37, 29–38. [Google Scholar] [CrossRef]
- Olsen, S.R.; Cole, C.V.; Watanabe, F.S.; Dean, L.A. Estimation of Available Phosphorus in Soils by Extraction with Sodium Bicarbonate; USDA-Circular, 939; US Department of Agriculture: Washington, DC, USA, 1954.
- Katanda, Y.; Zvomuya, F.; Flaten, D.; Cicek, N. Hog-Manure-Recovered Struvite: Effects on Canola and Wheat Biomass Yield and Phosphorus Use Efficiencies. Soil Sci. Soc. Am. J. 2016, 80, 135–146. [Google Scholar] [CrossRef]
- Cabeza, R.; Steingrobe, B.; Römer, W.; Claassen, N. Effectiveness of Recycled P Products as P Fertilizers, as Evaluated in Pot Experiments. Nutr. Cycl. Agroecosystems 2011, 91, 173–184. [Google Scholar] [CrossRef]
- Achat, D.L.; Sperandio, M.; Daumer, M.L.; Santellani, A.C.; Prud’Homme, L.; Akhtar, M.; Morel, C. Plant-Availability of Phosphorus Recycled from Pig Manures and Dairy Effluents as Assessed by Isotopic Labeling Techniques. Geoderma 2014, 232–234, 24–33. [Google Scholar] [CrossRef]
- Bonvin, C.; Etter, B.; Udert, K.M.; Frossard, E.; Nanzer, S.; Tamburini, F.; Oberson, A. Plant Uptake of Phosphorus and Nitrogen Recycled from Synthetic Source-Separated Urine. Ambio 2015, 44, 217–227. [Google Scholar] [CrossRef]
- Thompson, L. Field Evaluation of the Availability for Corn and Soybean of Phosphorus Recovered as Struvite from Corn Fiber Processing for Bioenergy. Master’s Thesis, Iowa State University, Ames, IA, USA, 2013. [Google Scholar]
- Ackerman, J.N.; Zvomuya, F.; Cicek, N.; Flaten, D. Evaluation of Manure-Derived Struvite as a Phosphorus Source for Canola. Can. J. Plant Sci. 2013, 93, 419–424. [Google Scholar] [CrossRef]
- Vogel, T.; Nelles, M.; Eichler-Löbermann, B. Phosphorus Effects of Recycled Products from Municipal Wastewater on Crops in a Field Experiment. Plant Soil Environ. 2017, 63, 475–482. [Google Scholar] [CrossRef]
- Vogel, T.; Nelles, M.; Eichler-Löbermann, B. Phosphorus Application with Recycled Products from Municipal Waste Water to Different Crop Species. Ecol. Eng. 2015, 83, 466–475. [Google Scholar] [CrossRef]
- Benjannet, R.; Nyiraneza, J.; Khiari, L.; Cambouris, A.; Fuller, K.; Hann, S.; Ziadi, N. Potato Response to Struvite Compared with Conventional Phosphorus Fertilizer in Eastern Canada. Agron. J. 2020, 112, 1360–1376. [Google Scholar] [CrossRef]
- Achat, D.L.; Daumer, M.L.; Sperandio, M.; Santellani, A.C.; Morel, C. Solubility and Mobility of Phosphorus Recycled from Dairy Effluents and Pig Manures in Incubated Soils with Different Characteristics. Nutr. Cycl. Agroecosystems 2014, 99, 1–15. [Google Scholar] [CrossRef]
- Johnston, A.E.; Richards, I.R. Effectiveness of Different Precipitated Phosphates as Phosphorus Sources for Plants. Soil Use Manag. 2003, 19, 45–49. [Google Scholar] [CrossRef]
- Massey, M.S.; Davis, J.G.; Ippolito, J.A.; Sheffield, R.E. Effectiveness of Recovered Magnesium Phosphates as Fertilizers in Neutral and Slightly Alkaline Soils. Agron. J. 2009, 101, 323–329. [Google Scholar] [CrossRef]
- Ylivainio, K.; Lehti, A.; Jermakka, J.; Wikberg, H.; Turtola, E. Predicting Relative Agronomic Efficiency of Phosphorus-Rich Organic Residues. Sci. Total Environ. 2021, 773, 145618. [Google Scholar] [CrossRef] [PubMed]
- Huygens, D.; Saveyn, H.G.M. Agronomic Efficiency of Selected Phosphorus Fertilisers Derived from Secondary Raw Materials for European Agriculture. A Meta-Analysis. Agron. Sustain. Dev. 2018, 38, 52. [Google Scholar] [CrossRef]
- Prater, J. Improved Production of Magnesium Ammonium Phosphate (Struvite) from Landfill Leachate 2014 Final Report; University of Wisconsin: Madison, WI, USA, 2015. [Google Scholar]
- Perez, R.C.; Steingrobe, B.; Römer, W.; Claassen, N. Plant Availability of P Fertilizers Recycled from Sewage Sludge and Meat-and-Bone Meal in Field and Pot Experiments. In International Conference on Nutrient Recovery from Wastewater Streams; IWA Publishing: Vancouver, BC, Canada, 2009. [Google Scholar]
- Acosta-Martínez, V.; Cruz, L.; Sotomayor-Ramírez, D.; Pérez-Alegría, L. Enzyme Activities as Affected by Soil Properties and Land Use in a Tropical Watershed. Appl. Soil Ecol. 2007, 35, 35–45. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L.; Lauber, C.L.; Weintraub, M.N.; Ahmed, B.; Allison, S.D.; Crenshaw, C.; Contosta, A.R.; Cusack, D.; Frey, S.; Gallo, M.E.; et al. Stoichiometry of Soil Enzyme Activity at Global Scale. Ecol. Lett. 2008, 11, 1252–1264. [Google Scholar] [CrossRef] [PubMed]
- Rietz, D.N.; Haynes, R.J. Effects of Irrigation-Induced Salinity and Sodicity on Soil Microbial Activity. Soil Biol. Biochem. 2003, 35, 845–854. [Google Scholar] [CrossRef]
- Nannipieri, P.; Giagnoni, L.; Landi, L.; Renella, G. Phosphorus in Action; Springer: Berlin/Heidelberg, Germany, 2011; Volume 26, pp. 215–243. [Google Scholar] [CrossRef]
- Allison, V.J.; Condron, L.M.; Peltzer, D.A.; Richardson, S.J.; Turner, B.L. Changes in Enzyme Activities and Soil Microbial Community Composition along Carbon and Nutrient Gradients at the Franz Josef Chronosequence, New Zealand. Soil Biol. Biochem. 2007, 39, 1770–1781. [Google Scholar] [CrossRef]
- Eivazi, F.; Tabatabai, M.A. Factors Affecting Glucosidase and Galactosidase Activities in Soils. Soil Biol. Biochem. 1990, 22, 891–897. [Google Scholar] [CrossRef]
- Melero, S.; López-Garrido, R.; Murillo, J.M.; Moreno, F. Conservation Tillage: Short- and Long-Term Effects on Soil Carbon Fractions and Enzymatic Activities under Mediterranean Conditions. Soil Tillage Res. 2009, 104, 292–298. [Google Scholar] [CrossRef]
- Derrien, D.; Marol, C.; Balesdent, J. The Dynamics of Neutral Sugars in the Rhizosphere of Wheat. An Approach By13C Pulse-Labelling and GC/C/IRMS. Plant Soil 2004, 267, 243–253. [Google Scholar] [CrossRef]
- Zhang, X.; Dippold, M.A.; Kuzyakov, Y.; Razavi, B.S. Spatial Pattern of Enzyme Activities Depends on Root Exudate Composition. Soil Biol. Biochem. 2019, 133, 83–93. [Google Scholar] [CrossRef]
- Bandick, A.K.; Dick, R.P. Field Management Effects on Soil Enzyme Activities. Soil Biol. Biochem. 1999, 31, 1471–1479. [Google Scholar] [CrossRef]
- Li, J.; Tong, X.; Awasthi, M.K.; Wu, F.; Ha, S.; Ma, J.; Sun, X.; He, C. Dynamics of Soil Microbial Biomass and Enzyme Activities along a Chronosequence of Desertified Land Revegetation. Ecol. Eng. 2018, 111, 22–30. [Google Scholar] [CrossRef]
- Dodgson, K.S.; White, G.F.; Fitzgerald, J. Sulfatases of Microbial Origin; CRC Press: Boca Raton, FL, USA, 1982. [Google Scholar]
- Deng, S.P.; Tabatabai, M.A. Effect of Tillage and Residue Management on Enzyme Activities in Soils: III. Phosphatases and Arylsulfatase. Biol. Fertil. Soils 1997, 24, 141–146. [Google Scholar] [CrossRef]
- Ryan, D.; Karpinska, A.; Forrestal, P.J.; Ashekuzzaman, S.M.; Kakouli-Duarte, T.; Dowling, D.N.; Germaine, K.J. The Impact of Bio-Based Fertilizer Integration Into Conventional Grassland Fertilization Programmes on Soil Bacterial, Fungal, and Nematode Communities. Front. Sustain. Food Syst. 2022, 6, 1–17. [Google Scholar] [CrossRef]
- Cregut, M.; Piutti, S.; Vong, P.C.; Slezack-Deschaumes, S.; Crovisier, I.; Benizri, E. Density, Structure, and Diversity of the Cultivable Arylsulfatase-Producing Bacterial Community in the Rhizosphere of Field-Grown Rape and Barley. Soil Biol. Biochem. 2009, 41, 704–710. [Google Scholar] [CrossRef]
- Cregut, M.; Piutti, S.; Slezack-Deschaumes, S.; Benizri, E. Compartmentalization and Regulation of Arylsulfatase Activities in Streptomyces Sp., Microbacterium Sp. and Rhodococcus Sp. Soil Isolates in Response to Inorganic Sulfate Limitation. Microbiol. Res. 2013, 168, 12–21. [Google Scholar] [CrossRef]
- Degens, B.P.; Harris, J.A. Development of a Physiological Approach to Measuring the Catabolic Diversity of Soil Microbial Communities. Soil Biol. Biochem. 1997, 29, 1309–1320. [Google Scholar] [CrossRef]
- Winding, A.; Hendriksen, N.B. Comparison of CLPP and Enzyme Activity Assay for Functional Characterization of Bacterial Soil Communities. J. Soils Sediments 2007, 7, 411–417. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| pH | EC | N | OM | P | Ca | Mg | Na | K | Pb | Cd | Cu | Ni | Zn | Cr | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| dS/m | % | % | mg/kg | ||||||||||||
| Soil | 8.57 ± 0.05 | 0.22 ± 0.07 | 0.15 ± 0.004 | 1.49 ± 0.03 | 29 ± 2 | 2877 ± 46 | 310 ± 23 | 17 ± 9 | 379 ± 43 | 19 ± 1 | <DL | 16 ± 1 | 13 ± 1 | 52 ± 5 | 57 ± 2 |
| STR | NPK | MAP | SSP | |
|---|---|---|---|---|
| Water soluble P2O5 (%) | 1.3 | 14.1 | 46.5 | 17.5 |
| Citrate-soluble P2O5 (%) | 22.3 | 15.2 | 61.6 | 18.0 |
| Soluble in mineral acids P2O5 (%) | 28.8 | 15.1 | 61.5 | 19.2 |
| Total N (%) | 5.7 | 14.4 | 12.2 | 0.5 |
| Ammonium N (%) | 5.5 | 14.1 | 11.9 | 0.3 |
| Nitrate N (%) | 0.1 | 0.2 | 0.2 | 0.1 |
| Ureic N (%) | <1.0 | <1.0 | <1.0 | <1.0 |
| K water soluble (in K2O) (%) | <1.0 | 15.3 | <1 | <1 |
| Cd (mg/kg DM) | <0.5 | <0.5 | <0.5 | 18.5 |
| Cu (mg/kg DM) | <20.0 | <20.0 | <20.0 | 21.5 |
| Cr (mg/kg DM) | <10.0 | <10.0 | <10.0 | 38.0 |
| Hg (mg/kg DM) | <0.4 | <0.4 | <0.4 | <0.4 |
| Ni (mg/kg DM) | <5.0 | <5.0 | <5.0 | 31.6 |
| Pb (mg/kg DM) | <5.0 | <5.0 | <5.0 | <5.0 |
| Zn (mg/kg DM) | <25.0 | <25.0 | <25.0 | 286 |
| As (mg/kg DM) | <2.0 | <2.0 | <2.0 | 5 |
| B (mg/kg DM) | <4.0 | <4.0 | <4.0 | 19.2 |
| Mo (mg/kg DM) | <0.5 | <0.5 | <0.5 | 17.9 |
| Mn (mg/kg DM) | 36.2 | 142.0 | <10.0 | 20.5 |
| N (%) | Ca (mg/kg) | Mg (mg/kg) | Na (mg/kg) | K (mg/kg) | P (mg/g) | ||
|---|---|---|---|---|---|---|---|
| 1st crop | NPK | 3.29 ± 0.18 | 12,946 ± 551 | 3140 ± 567 a | 1335 ± 601 | 114,122 ± 14,849 | 4.48 ± 0.31 a |
| STR | 3.51 ± 0.22 | 12,750 ± 471 | 3273 ± 26 ab | 891 ± 317 | 104,835 ± 2845 | 5.09 ± 0.32 a | |
| MAP | 3.08 ± 0.41 | 12,936 ± 1324 | 4390 ± 444 b | 1338 ± 461 | 100,234 ± 11,827 | 7.57 ± 0.88 b | |
| SSP | 3.36 ± 0.08 | 13,854 ± 1888 | 4263 ± 351 b | 1213 ± 369 | 119,833 ± 4434 | 6.46 ± 0.69 ab | |
| 2nd crop | NPK | 3.78 ± 0.16 | 9793 ± 2359 | 3339 ± 190 | 898 ± 352 | 106,764 ± 6027 | 6.11 ± 0.85 a |
| STR | 3.63 ± 0.32 | 10,010 ± 2614 | 3355 ± 514 | 1135 ± 209 | 99,485 ± 4656 | 6.68 ± 0.20 a | |
| MAP | 3.70 ± 0.28 | 11,705 ± 2736 | 3973 ± 528 | 1657 ± 1022 | 96,080 ± 11,380 | 6.72 ± 0.38 a | |
| SSP | 3.69 ± 0.46 | 13,487 ± 965 | 3896 ± 79 | 1161 ± 216 | 96,646 ± 8048 | 7.06 ± 0.33 a | |
| 3rd crop | NPK | 3.35 ± 0.25 | 14,843 ± 2946 | 3727 ± 592 | 1795 ± 521 | 84,791 ± 6894 | 3.47 ± 0.14 a |
| STR | 3.35 ± 0.16 | 11,619 ± 704 | 3311 ± 196 | 1769 ± 387 | 83,875 ± 5706 | 4.40 ± 0.28 b | |
| MAP | 3.15 ± 0.25 | 12,084 ± 2546 | 3424 ± 139 | 1904 ± 622 | 71,374 ± 12,626 | 4.64 ± 0.33 b | |
| SSP | 3.14 ± 0.29 | 11,952 ± 968 | 3299 ± 198 | 2016 ± 391 | 78,564 ± 5768 | 4.95 ± 0.28 b |
| pH | C.E. | N | P | Ca | Mg | Na | K | Pb | Cd | Cu | Ni | Zn | Cr | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| dS/m | % | mg/kg | |||||||||||||
| 1st crop | NPK | 8.02 ± 0.23 | 0.544 ± 0.162 ab | 0.16 ± 0.02 | 68 ± 8 a | 3065 ± 258 | 383 ± 28 a | 37 ± 9 | 549 ± 88 | 15 ± 3.86 | <DL | 16 ± 0.50 | 12 ± 0.58 | 47 ± 2.38 | 37 ± 4.51 |
| STR | 8.32 ± 0.11 | 0.212 ± 0.031 a | 0.16 ± 0.01 | 83 ± 21 a | 2787 ± 59 | 456 ± 10 c | 25 ± 3 | 407 ± 7 | 14 ± 3.50 | <DL | 15 ± 0.50 | 12 ± 0.50 | 46 ± 2.22 | 34 ± 1.50 | |
| MAP | 8.26 ± 0.16 | 0.397 ± 0.246 ab | 0.17 ± 0.04 | 136 ± 52 a | 2666 ± 95 | 402 ± 6 b | 36 ± 13 | 453 ± 101 | 14 ± 1.29 | <DL | 14 ± 0.50 | 14 ± 1.29 | 47 ± 0.82 | 36 ± 1.29 | |
| SSP | 8.09 ± 0.18 | 0.571 ± 0.101 b | 0.15 ± 0.01 | 103 ± 39 a | 2882 ± 266 | 413 ± 26 b | 41 ± 15 | 442 ± 60 | 13 ± 0.50 | <DL | 15 ± 0.50 | 12 ± 0.50 | 48 ± 1.63 | 34 ± 1.26 | |
| 2nd crop | NPK | 8.17 ± 0.07 | 0.503 ± 0.169 | 0.14 ± 0.01 | 40 ± 5 a | 2859 ± 250 | 358 ± 12 | 32 ± 7 | 363 ± 42 | 16 ± 0.50 | <DL | 16 ± 0.50 | 13 ± 0.50 | 56 ± 1.71 | 35 ± 0.82 |
| STR | 8.23 ± 0.08 | 0.375 ± 0.091 | 0.15 ± 0.01 | 82 ± 9 c | 3081 ± 125 | 404 ± 30 | 32 ± 6 | 354 ± 39 | 16 ± 0.58 | <DL | 16 ± 0.58 | 12 ± 0.58 | 51 ± 4.08 | 34 ± 9.11 | |
| MAP | 8.38 ± 0.23 | 0.324 ± 0.092 | 0.14 ± 0.01 | 69 ± 13 bc | 3130 ± 172 | 390 ± 23 | 45 ± 3 | 301 ± 45 | 16 ± 0.50 | <DL | 17 ± 1.71 | 13 ± 2.06 | 51 ± 3.50 | 33 ± 4.51 | |
| SSP | 8.15 ± 0.28 | 0.544 ± 0.279 | 0.14 ± 0.02 | 68 ± 12 b | 3220 ± 271 | 401 ± 53 | 47 ± 26 | 322 ± 65 | 17 ± 0.00 | <DL | 17 ± 0.58 | 11 ± 0.58 | 55 ± 2.00 | 35 ± 0.58 | |
| 3rd crop | NPK | 8.03 ± 0.16 | 0.493 ± 0.291 | 0.15 ± 0.01 | 46 ± 8 a | 3097 ± 166 | 411 ± 39 | 44 ± 21 | 316 ± 52 | 19 ± 0.58 | <DL | 16 ± 0.50 | 15 ± 2.22 | 56 ± 0.96 | 36 ± 4.04 |
| STR | 8.16 ± 0.14 | 0.379 ± 0.084 | 0.15 ± 0.01 | 61 ± 10 b | 2830 ± 170 | 452 ± 56 | 49 ± 11 | 271 ± 18 | 19 ± 0.50 | <DL | 16 ± 0.50 | 14 ± 0.50 | 55 ± 0.96 | 39 ± 3.10 | |
| MAP | 8.04 ± 0.26 | 0.694 ± 0.310 | 0.15 ± 0.02 | 66 ± 4 b | 3083 ± 165 | 456 ± 9 | 68 ± 24 | 247 ± 36 | 19 ± 1.26 | <DL | 18 ± 2.06 | 18 ± 8.68 | 57 ± 2.16 | 37 ± 5.29 | |
| SSP | 8.08 ± 0.15 | 0.594 ± 0.198 | 0.14 ± 0.01 | 61 ± 5 b | 2847 ± 244 | 433 ± 17 | 61 ± 16 | 256 ± 35 | 19 ± 1.83 | <DL | 16 ± 0.58 | 14 ± 1.00 | 57 ± 1.89 | 39 ± 5.19 | |
| NPK | STR | MAP | SSP | ||
|---|---|---|---|---|---|
| Amino acids | L-Threonine | 0.413 a | 0.297 b | 0 | 0 |
| L-Serine | 0.730 b | 0.751 b | 0.997 a | 0.243 c | |
| L-Asparagine | 1.157 b | 1.412 a | 1.152 b | 0.543 b | |
| L-Arginine | 0.498 b | 0.920 a | 0.457 b | 0.464 b | |
| Amines | Putrescine | 0.458 a | 0.553 a | 0.326 b | 0.229 b |
| Phenylethyl-amine | 0.611 a | 0.470 b | 0 | 0 | |
| Phenolic compounds | 4-Hydroxy Benzoic Acid | 0.401 b | 0.822 a | 0.358 b | 0.488 b |
| Carbohydrates | D-Galactonic Acid γ-Lactone | 0.414 c | 0.710 a | 0.502 b | 0.563 b |
| Glucose-1-Phosphate | 0.483 a | 0.521 a | 0 | 0 | |
| N-Acetyl-D-Glucosamine | 0.308 c | 0.540 ab | 0.589 a | 0.475 b | |
| D-Mannitol | 0.783 b | 0.919 a | 0.815 ab | 0.747 b | |
| β-Methyl-D-Glucoside | 0.326 a | 0.319 a | 0 | 0 | |
| Carboxylic acids | D-Glucosaminic Acid | 0.486 c | 0.759 a | 0.534 b | 0.453 c |
| Pyruvic Acid Methyl Ester | 0.689 b | 0.888 a | 0.682 b | 0.637 b | |
| D-Malic Acid | 0.340 a | 0.338 a | 0.211 b | 0 | |
| γ-Hydroxybutyric Acid | 0.430 b | 0.876 a | 0 | 0 | |
| D-Galacturonic Acid | 1.726 a | 1.480 b | 1.650 a | 0.865 c | |
| Polymers | Tween 80 | 1.049 a | 1.083 a | 0.904 b | 0.521 c |
| Tween 40 | 0.689 b | 0.888 a | 0.882 a | 0.637 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mancho, C.; Diez-Pascual, S.; Alonso, J.; Gil-Díaz, M.; García-Gonzalo, P.; Lobo, M.C. Medium/Long-Term Efficiency of Struvite for Lettuce (Lactuca sativa L.) Production: Effect on Soil Quality. Horticulturae 2023, 9, 645. https://doi.org/10.3390/horticulturae9060645
Mancho C, Diez-Pascual S, Alonso J, Gil-Díaz M, García-Gonzalo P, Lobo MC. Medium/Long-Term Efficiency of Struvite for Lettuce (Lactuca sativa L.) Production: Effect on Soil Quality. Horticulturae. 2023; 9(6):645. https://doi.org/10.3390/horticulturae9060645
Chicago/Turabian StyleMancho, Carolina, Sergio Diez-Pascual, Juan Alonso, Mar Gil-Díaz, Pilar García-Gonzalo, and M. Carmen Lobo. 2023. "Medium/Long-Term Efficiency of Struvite for Lettuce (Lactuca sativa L.) Production: Effect on Soil Quality" Horticulturae 9, no. 6: 645. https://doi.org/10.3390/horticulturae9060645
APA StyleMancho, C., Diez-Pascual, S., Alonso, J., Gil-Díaz, M., García-Gonzalo, P., & Lobo, M. C. (2023). Medium/Long-Term Efficiency of Struvite for Lettuce (Lactuca sativa L.) Production: Effect on Soil Quality. Horticulturae, 9(6), 645. https://doi.org/10.3390/horticulturae9060645