
Effects of Citric Acid and Humic-like Substances on Yield, Enzyme Activities, and Expression of Genes Involved in Iron Uptake in Tomato Plants


 ,
,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Growth Conditions
2.2. Enzymatic Activity
2.3. Real-Time Reverse-Transcriptase PCR
2.4. Data Analysis
3. Results
3.1. Plant Growth
3.2. Enzymatic Activity
3.2.1. H+-ATPase Activity
3.2.2. PEPC Activity
3.2.3. FRO Activity
3.3. Gene Expression
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Murgia, I.; Marzorati, F.; Vigani, G.; Morandini, P. Plant iron nutrition: The long road from soil to seeds. J. Exp. Bot. 2022, 73, 1809–1824. [Google Scholar] [CrossRef] [PubMed]
- Mori, S. Iron acquisition by plants. Curr. Opin. Plant Biol. 1999, 2, 250–253. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, T.; Nozoye, T.; Nishizawa, N.K. Iron transport and its regulation in plants. Free Radic. Biol. Med. 2019, 133, 11–20. [Google Scholar] [CrossRef]
- Wahba, M.M.; Labib, F.; Zaghloul, A. Management of Calcareous Soils in Arid Region. Int. J. Environ. Pollut. Environ. Model. 2019, 2, 248–258. [Google Scholar]
- Marschner, H.; Römheld, V. Strategies of plants for acquisition of iron. Plant Soil 1994, 165, 261–274. [Google Scholar] [CrossRef]
- Martín-Barranco, A.; Thomine, S.; Vert, G.; Zelazny, E. A quick journey into the diversity of iron uptake strategies in photosynthetic organisms. Plant Signal. Behav. 2021, 16, 1975088. [Google Scholar] [CrossRef] [PubMed]
- Chao, Z.F.; Chao, D.Y. Similarities and differences in iron homeostasis strategies between graminaceous and nongraminaceous plants. New Phytol. 2022, 236, 1655–1660. [Google Scholar] [CrossRef]
- Li, S.; Song, Z.; Liu, X.; Zhou, X.; Yang, W.; Chen, J.; Chen, R. Mediation of Zinc and Iron Accumulation in Maize by ZmIRT2, a Novel Iron-Regulated Transporter. Plant Cell Physiol. 2022, 63, 521–534. [Google Scholar] [CrossRef]
- Santi, S.; Schmidt, W. Dissecting iron deficiency-induced proton extrusion in Arabidopsis roots. New Phytol. 2009, 183, 1072–1084. [Google Scholar] [CrossRef]
- Gupta, P.K.; Balyan, H.S.; Sharma, S.; Kumar, R. Biofortification and bioavailability of Zn, Fe and Se in wheat: Present status and future prospects. Theor. Appl. Genet. 2021, 134, 1–35. [Google Scholar] [CrossRef]
- Schwarz, B.; Bauer, P. FIT, a regulatory hub for iron deficiency and stress signaling in roots, and FIT-dependent and -independent gene signatures. J. Exp. Bot. 2020, 71, 1694–1705. [Google Scholar] [CrossRef] [PubMed]
- Rosenkranz, T.; Oburger, E.; Baune, M.; Weber, G.; Puschenreiter, M. Root exudation of coumarins from soil-grown Arabidopsis thaliana in response to iron deficiency. Rhizosphere 2021, 17, 100296. [Google Scholar] [CrossRef]
- Vélez-Bermúdez, I.C.; Schmidt, W. Plant strategies to mine iron from alkaline substrates. Plant Soil 2023, 483, 1–25. [Google Scholar] [CrossRef]
- Jiménez, M.R.; Casanova, L.; Saavedra, T.; Gama, F.; Suárez, M.P.; Correia, P.J.; Pestana, M. Responses of tomato (Solanum lycopersicum L.) plants to iron deficiency in the root zone. Folia Hortic. 2019, 31, 223–234. [Google Scholar] [CrossRef]
- Kabir, A.H.; Paltridge, N.G.; Able, A.J.; Paull, J.G.; Stangoulis, J.C.R. Natural variation for Fe-efficiency is associated with upregulation of Strategy I mechanisms and enhanced citrate and ethylene synthesis in Pisum sativum L. Planta 2012, 235, 1409–1419. [Google Scholar] [CrossRef] [PubMed]
- Santi, S.; Cesco, S.; Varanini, Z.; Pinton, R. Two plasma membrane H+-ATPase genes are differentially expressed in iron-deficient cucumber plants. Plant Physiol. Biochem. 2005, 43, 287–292. [Google Scholar] [CrossRef]
- Fan, Z.; Wu, Y.; Zhao, L.; Fu, L.; Deng, L.; Deng, J.; Ding, D.; Xiao, S.; Deng, X.; Peng, S.; et al. MYB308-mediated transcriptional activation of plasma membrane H+-ATPase 6 promotes iron uptake in citrus. Hortic. Res. 2022, 9, uhac088. [Google Scholar] [CrossRef]
- Filiz, E.; Kurt, F. FIT (Fer-like iron deficiency-induced transcription factor) in plant iron homeostasis: Genome-wide identification and bioinformatics analyses. J. Plant Biochem. Biotechnol. 2019, 28, 143–157. [Google Scholar] [CrossRef]
- Ferreira, C.M.H.; López-Rayo, S.; Lucena, J.J.; Soares, E.V.; Soares, H. Evaluation of the Efficacy of Two New Biotechnological-Based Freeze-Dried Fertilizers for Sustainable Fe Deficiency Correction of Soybean Plants Grown in Calcareous Soils. Front. Plant Sci. 2019, 10, 1335. [Google Scholar] [CrossRef]
- Zanin, L.; Tomasi, N.; Cesco, S.; Varanini, Z.; Pinton, R. Humic substances contribute to plant iron nutrition acting as chelators and biostimulants. Front. Plant Sci. 2019, 10, 675. [Google Scholar] [CrossRef]
- Pérez-Labrada, F.; Benavides-Mendoza, A.; Juárez-Maldonado, A.; Solís-Gaona, S.; González-Morales, S. Organic acids combined with Fe-chelate improves ferric nutrition in tomato grown in calcisol soil. J. Soil Sci. Plant Nutr. 2020, 20, 673–683. [Google Scholar] [CrossRef]
- Al-Balawna, Z.A.; Abu-Abdoun, I.I. Fate of Citric Acid Addition on Mineral Elements Availability in Calcareous Soils of Jordan Valley. Int. Res. J. Pure Appl. Chem. 2021, 22, 82–89. [Google Scholar] [CrossRef]
- Olego, M.Á.; Cuesta Lasso, M.; Quiroga, M.J.; Visconti, F.; López, R.; Garzón-Jimeno, E. Effects of Leonardite Amendments on Vineyard Calcareous Soil Fertility, Vine Nutrition and Grape Quality. Plants 2022, 11, 356. [Google Scholar] [CrossRef]
- Zanin, L.; Tomasi, N.; Rizzardo, C.; Gottardi, S.; Terzano, R.; Alfeld, M.; Janssens, K.; De Nobili, M.; Mimmo, T.; Cesco, S. Iron allocation in leaves of Fe-deficient cucumber plants fed with natural Fe complexes. Physiol. Plant. 2015, 154, 82–94. [Google Scholar] [CrossRef] [PubMed]
- Zamboni, A.; Zanin, L.; Tomasi, N.; Avesani, L.; Pinton, R.; Varanini, Z.; Cesco, S. Early transcriptomic response to Fe supply in Fe-deficient tomato plants is strongly influenced by the nature of the chelating agent. BMC Genom. 2016, 17, 35. [Google Scholar] [CrossRef] [PubMed]
- Cieschi, M.T.; Lucena, J.J. Leonardite iron humate and synthetic iron chelate mixtures in Glycine max nutrition. J. Sci. Food Agric. 2021, 101, 4207–4219. [Google Scholar] [CrossRef]
- Steiner, A.A. A universal method for preparing nutrient solutions of a certain desired composition. Plant Soil 1961, 15, 134–154. [Google Scholar] [CrossRef]
- Rabotti, G.; Zocchi, G. Plasma membrane-bound H+-ATPase and reductase activities in Fe-deficient cucumber roots. Physiol. Plant. 1994, 90, 779–785. [Google Scholar] [CrossRef]
- Palmgren, M.G.; Askerlund, P.; Fredrikson, K.; Widell, S.; Sommarin, M.; Larsson, C. Sealed Inside-Out and Right-Side-Out Plasma Membrane Vesicles. Plant Physiol. 1990, 92, 871–880. [Google Scholar] [CrossRef]
- Nisi, P.D.; Zochi, G. Phosphoenolpyruvate carboxylase in cucumber (Cucumis sativus L.) roots under iron deficiency: Activity and kinetic characterization. J. Exp. Bot. 2000, 51, 1903–1909. [Google Scholar] [CrossRef]
- Romera, F.; Welch, R.; Norvell, W.; Schaefer, S.; Kochian, L. Ethylene involvement in the over-expression of Fe(III)-chelate reductase by roots of E107 pea [Pisum sativum L. (brz, brz)] and chloronerva tomato (Lycopersicon esculentum L.) mutant genotypes. Biometals 1996, 9, 38–44. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Rio, D.C.; Ares, M.; Hannon, G.J.; Nilsen, T.W. Purification of RNA Using TRIzol (TRI Reagent). Cold Spring Harb. Protoc. 2010, 2010, pdb-prot5439. [Google Scholar] [CrossRef] [PubMed]
- Paolacci, A.R.; Celletti, S.; Catarcione, G.; Hawkesford, M.J.; Astolfi, S.; Ciaffi, M. Iron deprivation results in a rapid but not sustained increase of the expression of genes involved in iron metabolism and sulfate uptake in tomato (Solanum lycopersicum L.) seedlings. J. Integr. Plant Biol. 2014, 56, 88–100. [Google Scholar] [CrossRef]
- Diamantopoulos, P.D.; Aivalakis, G.; Flemetakis, E.; Katinakis, P. Expression of three β-type carbonic anhydrases in tomato fruits. Mol. Biol. Rep. 2013, 40, 4189–4196. [Google Scholar] [CrossRef]
- Larionov, A.; Krause, A.; Miller, W. A standard curve based method for relative real time PCR data processing. BMC Bioinform. 2005, 6, 62. [Google Scholar] [CrossRef]
- Dembélé, D.; Kastner, P. Fold change rank ordering statistics: A new method for detecting differentially expressed genes. BMC Bioinform. 2014, 15, 14. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Pérez-Labrada, F.; Mendoza, A.B.; Valdez-Aguilar, L.A.; Robledo-Torres, V. Citric acid in the nutrient solution increases the mineral absorption in potted tomato grown in calcareous soil. Pakistan J. Bot. 2016, 48, 67–74. [Google Scholar]
- Massimi, M.; Radócz, L.; Csótó, A. Impact of Organic Acids and Biological Treatments in Foliar Nutrition on Tomato and Pepper Plants. Horticulturae 2023, 9, 413. [Google Scholar] [CrossRef]
- Sharma, S.; Anand, N.; Bindraban, P.S.; Pandey, R. Foliar Application of Humic Acid with Fe Supplement Improved Rice, Soybean, and Lettuce Iron Fortification. Agriculture 2023, 13, 132. [Google Scholar] [CrossRef]
- Zia-ur-Rehman, M.; Bani Mfarrej, M.F.; Usman, M.; Azhar, M.; Rizwan, M.; Alharby, H.F.; Bamagoos, A.A.; Alshamrani, R.; Ahmad, Z. Exogenous application of low and high molecular weight organic acids differentially affected the uptake of cadmium in wheat-rice cropping system in alkaline calcareous soil. Environ. Pollut. 2023, 329, 121682. [Google Scholar] [CrossRef] [PubMed]
- Cieschi, M.T.; Polyakov, A.Y.; Lebedev, V.A.; Volkov, D.S.; Pankratov, D.A.; Veligzhanin, A.A.; Perminova, I.V.; Lucena, J.J. Eco-friendly iron-humic nanofertilizers synthesis for the prevention of iron chlorosis in soybean (Glycine max) grown in calcareous soil. Front. Plant Sci. 2019, 10, 413. [Google Scholar] [CrossRef] [PubMed]
- Justi, M.; Silva, C.A.; Rosa, S.D. Organic acids as complexing agents for iron and their effects on the nutrition and growth of maize and soybean. Arch. Agron. Soil Sci. 2022, 68, 1369–1384. [Google Scholar] [CrossRef]
- Jalali, M.; Jalali, M. Effect of Low-Molecular-Weight Organic Acids on the Release of Phosphorus from Amended Calcareous Soils: Experimental and Modeling. J. Soil Sci. Plant Nutr. 2022, 22, 4179–4193. [Google Scholar] [CrossRef]
- Karadihalli Thammaiah, M.; Pandey, R.N.; Purakayastha, T.J.; Chobhe, K.A.; Vashisth, A.; Chandra, S.; Pawar, A.B.; Trivedi, A. Impact of Low Molecular Weight Organic Acids on Soil Phosphorus Release and Availability to Wheat. Commun. Soil Sci. Plant Anal. 2022, 53, 2497–2508. [Google Scholar] [CrossRef]
- Zhao, K.; Wang, C.; Xiao, X.; Li, M.; Zhao, W.; Wang, Y.; Yang, Y. The Hormetic Response of Soil P Extraction Induced by Low-Molecular-Weight Organic Acids. Processes 2023, 11, 216. [Google Scholar] [CrossRef]
- Sun, Q.; Liu, J.; Huo, L.; Li, Y.C.; Li, X.; Xia, L.; Zhou, Z.; Zhang, M.; Li, B. Humic acids derived from Leonardite to improve enzymatic activities and bioavailability of nutrients in a calcareous soil. Int. J. Agric. Biol. Eng. 2020, 13, 200–205. [Google Scholar] [CrossRef]
- Abdulla, A.A.; Esmai, A.O.; Yaseen, H.S. Combination Influence of Humic Acid and Chelated Iron on yield and quality of Broccoli (Brassica oleracea L.) in Erbil, Iraqi Kurdistan Region. ZANCO J. Pure Appl. Sci. 2023, 35, 126–135. [Google Scholar] [CrossRef]
- Gayomba, S.R.; Zhai, Z.; Jung, H.; Vatamaniuk, O.K. Local and systemic signaling of iron status and its interactions with homeostasis of other essential elements. Front. Plant Sci. 2015, 6, 716. [Google Scholar] [CrossRef]
- Palmgren, M.G. Plant plasma membrane H+-ATPases: Powerhouses for Nutrient Uptake. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2001, 52, 817–845. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zhao, R.; Yu, J.; Gu, J.; Li, Y.; Chen, W.; Guo, W. Functional analysis of plasma membrane H+-ATPases in response to alkaline stress in blueberry. Sci. Hortic. 2022, 306, 111453. [Google Scholar] [CrossRef]
- Canellas, L.P.; Olivares, F.L.; Aguiar, N.O.; Jones, D.L.; Nebbioso, A.; Mazzei, P.; Piccolo, A. Humic and fulvic acids as biostimulants in horticulture. Sci. Hortic. 2015, 196, 15–27. [Google Scholar] [CrossRef]
- Elena, A.; Diane, L.; Eva, B.; Marta, F.; Roberto, B.; Zamarreño, A.M.; García-Mina, J.M. The root application of a purified leonardite humic acid modifies the transcriptional regulation of the main physiological root responses to Fe deficiency in Fe-sufficient cucumber plants. Plant Physiol. Biochem. 2009, 47, 215–223. [Google Scholar] [CrossRef]
- Mora, V.; Bacaicoa, E.; Zamarreño, A.-M.; Aguirre, E.; Garnica, M.; Fuentes, M.; García-Mina, J.-M. Action of humic acid on promotion of cucumber shoot growth involves nitrate-related changes associated with the root-to-shoot distribution of cytokinins, polyamines and mineral nutrients. J. Plant Physiol. 2010, 167, 633–642. [Google Scholar] [CrossRef]
- Alghamdi, S.A.; Al-Ghamdi, F.A.M.; El-Zohri, M.; Al-Ghamdi, A.A.M. Modifying of calcareous soil with some acidifying materials and its effect on Helianthus annuus (L.) growth. Saudi J. Biol. Sci. 2023, 30, 103568. [Google Scholar] [CrossRef]
- Astolfi, S.; Pii, Y.; Mimmo, T.; Lucini, L.; Miras-Moreno, M.B.; Coppa, E.; Violino, S.; Celletti, S.; Cesco, S. Single and Combined Fe and S Deficiency Differentially Modulate Root Exudate Composition in Tomato: A Double Strategy for Fe Acquisition? Int. J. Mol. Sci. 2020, 21, 4038. [Google Scholar] [CrossRef]
- Martinez-Cuenca, M.-R.; Iglesias, D.J.; Talon, M.; Abadia, J.; Lopez-Millan, A.-F.; Primo-Millo, E.; Legaz, F. Metabolic responses to iron deficiency in roots of Carrizo citrange [Citrus sinensis (L.) Osbeck. × Poncirus trifoliata (L.) Raf.]. Tree Physiol. 2013, 33, 320–329. [Google Scholar] [CrossRef]
- Covarrubias, J.I.; Rombolà, A.D. Organic acids metabolism in roots of grapevine rootstocks under severe iron deficiency. Plant Soil 2015, 394, 165–175. [Google Scholar] [CrossRef]
- Alhendawi, R.A.M.; Mohamed, A.A.M. The influence of high pH on maize growth and utilization of micronutrients under various concentrations of bicarbonates. Am. J. Agric. Environ. Sci. 2015, 15, 259–264. [Google Scholar] [CrossRef]
- Kong, D.; Chen, C.; Wu, H.; Li, Y.; Li, J.; Ling, H.-Q. Sequence Diversity and Enzyme Activity of Ferric-Chelate Reductase LeFRO1 in Tomato. J. Genet. Genomics 2013, 40, 565–573. [Google Scholar] [CrossRef]
- Larbi, A.; Morales, F.; Abadía, A.; Abadía, J. Changes in iron and organic acid concentrations in xylem sap and apoplastic fluid of iron-deficient Beta vulgaris plants in response to iron resupply. J. Plant Physiol. 2010, 167, 255–260. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Liu, S.; Li, F.; Sun, M.; Liang, Z.; Sun, Z.; Yu, F.; Li, H. The low ferric chelate reductase activity and high apoplastic pH in leaves cause iron deficiency chlorosis in ‘Huangguan’ pears grafted onto quince A grown in calcareous soil. Sci. Hortic. 2023, 310, 111754. [Google Scholar] [CrossRef]
- Zamboni, A.; Zanin, L.; Tomasi, N.; Pezzotti, M.; Pinton, R.; Varanini, Z.; Cesco, S. Genome-wide microarray analysis of tomato roots showed defined responses to iron deficiency. BMC Genom. 2012, 13, 101. [Google Scholar] [CrossRef] [PubMed]
- Skogerboe, R.K.; Wilson, S.A. Reduction of ionic species by fulvic acid. Anal. Chem. 1981, 53, 228–232. [Google Scholar] [CrossRef]
- Struyk, Z.; Sposito, G. Redox properties of standard humic acids. Geoderma 2001, 102, 329–346. [Google Scholar] [CrossRef]
- Yang, F.; Tang, C.; Antonietti, M. Natural and artificial humic substances to manage minerals, ions, water, and soil microorganisms. Chem. Soc. Rev. 2021, 50, 6221–6239. [Google Scholar] [CrossRef]
- Cieschi, M.T.; Lucena, J.J. Iron and Humic Acid Accumulation on Soybean Roots Fertilized with Leonardite Iron Humates under Calcareous Conditions. J. Agric. Food Chem. 2018, 66, 13386–13396. [Google Scholar] [CrossRef]
- Tahjib-Ul-Arif, M.; Zahan, M.I.; Karim, M.M.; Imran, S.; Hunter, C.T.; Islam, M.S.; Mia, M.A.; Hannan, M.A.; Rhaman, M.S.; Hossain, M.A.; et al. Citric Acid-Mediated Abiotic Stress Tolerance in Plants. Int. J. Mol. Sci. 2021, 22, 7235. [Google Scholar] [CrossRef]
- Tomasi, N.; De Nobili, M.; Gottardi, S.; Zanin, L.; Mimmo, T.; Varanini, Z.; Römheld, V.; Pinton, R.; Cesco, S. Physiological and molecular characterization of Fe acquisition by tomato plants from natural Fe complexes. Biol. Fertil. Soils 2013, 49, 187–200. [Google Scholar] [CrossRef]
- Lee, S.; Rahman, M.M.; Nakanishi, H.; Nishizawa, N.K.; An, G.; Nam, H.G.; Jeon, J.-S. Concomitant Activation of OsNAS2 and OsNAS3 Contributes to the Enhanced Accumulation of Iron and Zinc in Rice. Int. J. Mol. Sci. 2023, 24, 6568. [Google Scholar] [CrossRef] [PubMed]
- Vigani, G.; Pii, Y.; Celletti, S.; Maver, M.; Mimmo, T.; Cesco, S.; Astolfi, S. Mitochondria dysfunctions under Fe and S deficiency: Is citric acid involved in the regulation of adaptive responses? Plant Physiol. Biochem. 2018, 126, 86–96. [Google Scholar] [CrossRef] [PubMed]
- Waseem, M.; Ahmad, F. The phosphoenolpyruvate carboxylase gene family identification and expression analysis under abiotic and phytohormone stresses in Solanum lycopersicum L. Gene 2019, 690, 11–20. [Google Scholar] [CrossRef]
- An, Y.; Zhou, P.; Xiao, Q.; Shi, D. Effects of foliar application of organic acids on alleviation of aluminum toxicity in alfalfa. J. Plant Nutr. Soil Sci. 2014, 177, 421–430. [Google Scholar] [CrossRef]
- Hsieh, E.-J.; Waters, B.M. Alkaline stress and iron deficiency regulate iron uptake and riboflavin synthesis gene expression differently in root and leaf tissue: Implications for iron deficiency chlorosis. J. Exp. Bot. 2016, 67, 5671–5685. [Google Scholar] [CrossRef]
- Li, L.; Cheng, X.; Ling, H.-Q. Isolation and characterization of Fe(III)-chelate reductase gene LeFRO1 in tomato. Plant Mol. Biol. 2004, 54, 125–136. [Google Scholar] [CrossRef]
- Tomasi, N.; Rizzardo, C.; Monte, R.; Gottardi, S.; Jelali, N.; Terzano, R.; Vekemans, B.; De Nobili, M.; Varanini, Z.; Pinton, R.; et al. Micro-analytical, physiological and molecular aspects of Fe acquisition in leaves of Fe-deficient tomato plants re-supplied with natural Fe-complexes in nutrient solution. Plant Soil 2009, 325, 25–38. [Google Scholar] [CrossRef]
- Drincovich, M.F.; Voll, L.M.; Maurino, V.G. Editorial: On the Diversity of Roles of Organic Acids. Front. Plant Sci. 2016, 7, 1592. [Google Scholar] [CrossRef]



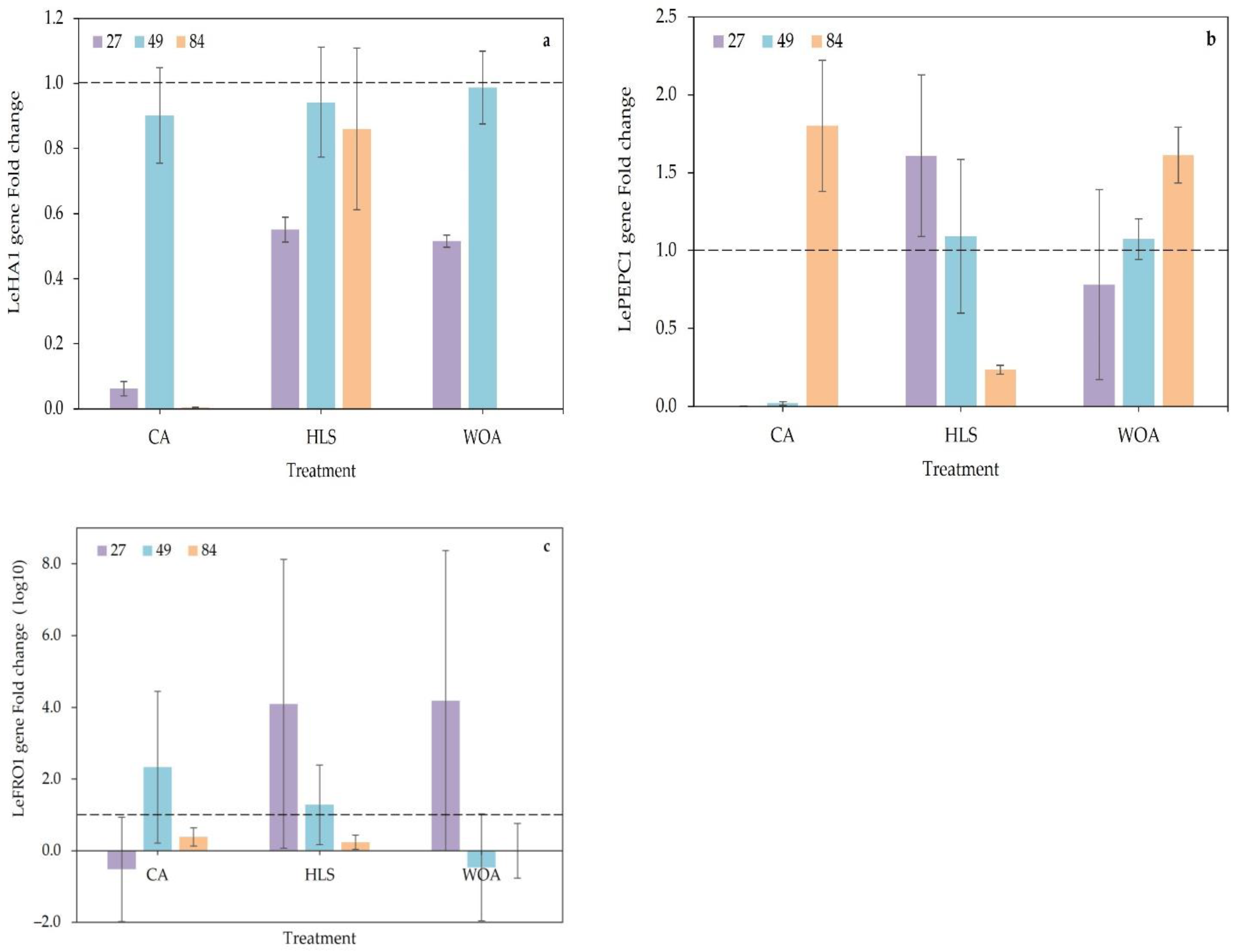{kind=link}
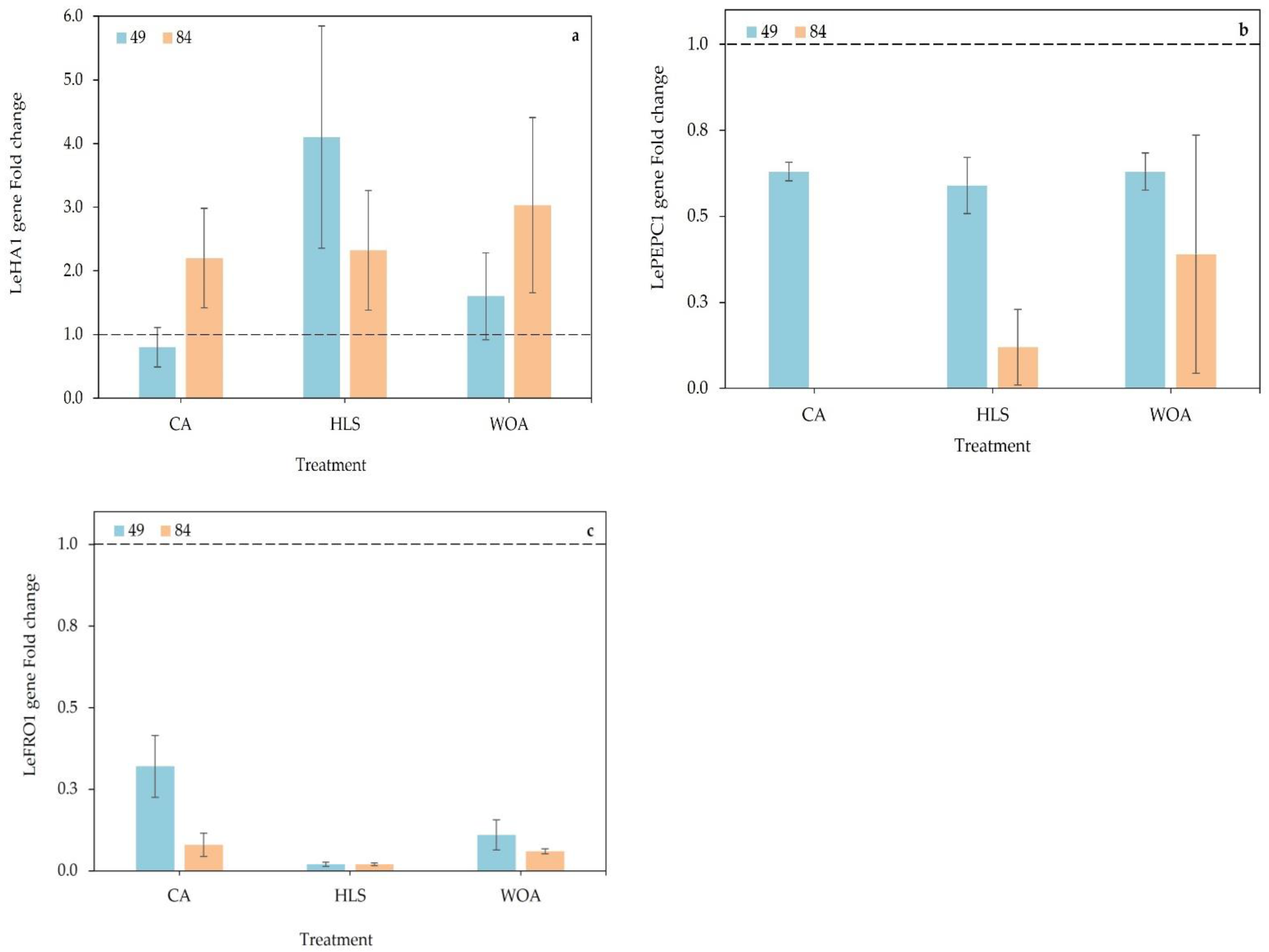{kind=link}
| Name Gene | Nomenclature | Forward Primer 5′-3′ | Reverse Primer 5′-3′ | Tm (°C) |
|---|---|---|---|---|
| Actin | ACTIN | CCCAGGCACACAGGTGTTAT | CAGGAGCAACTCGAAGCTCA | 60 |
| H+-ATPase | LeHA1 | GAACCCTTCATGGGCTCCAA | GCAACTCACGTAGCCTAGCA | 60 |
| PEPC | LePEPC1 | TGCTGCATTGTTCGACAAGC | CAAAAGTTCGCCGAAAGACAAC | 60 |
| FRO | LeFRO1 | GCGGTGTTGAATATGCTAATC | AAACTTTCCATCTCCCTATCG | 60 |
| Sampling | Treatment | Stem Diameter (mm) | Plant Height (cm) | Number of Leaves | SPAD-Unit | Total of Fruit Harvest | Production per Plant (kg) |
|---|---|---|---|---|---|---|---|
| 27 DAT | CA | 10.80 ± 0.52 a† | 45.70 ± 3.38 a | 13.20 ± 0.45 a | 54.19 ± 2.78 a | - | - |
| HLS | 10.24 ± 0.96 a | 39.40 ± 4.39 b | 13.40 ± 0.89 a | 53.65 ± 2.38 a | - | - | |
| WOA | 10.00 ± 0.29 a | 45.50 ± 3.87 a | 12.60 ± 0.89 a | 53.85 ± 2.95 a | - | - | |
| ID | 8.22 ± 1.90 b | 32.80 ± 5.54 c | 10.80 ± 2.17 b | 33.55 ± 5.64 b | - | - | |
| 49 DAT | CA | 11.66 ± 1.21 a | 71.20 ± 3.63 a | 18.00 ± 2.24 a | 55.43 ± 3.42 a | - | - |
| HLS | 12.54 ± 1.03 a | 68.20 ± 8.32 a | 15.20 ± 2.17 b | 57.91 ± 3.51 a | - | - | |
| WOA | 12.02 ± 0.45 a | 73.40 ± 5.94 a | 18.00 ± 0.71 a | 54.85 ± 2.17 a | - | - | |
| ID | 9.74 ± 0.56 b | 53.40 ± 2.30 b | 12.60 ± 1.67 c | 27.51 ± 5.13 b | - | - | |
| 84 DAT | CA | 12.98 ± 1.28 ab | 101.60 ± 11.04 a | 23.00 ± 2.35 a | 54.21 ± 4.12 a | 57.75 ± 6.44 ab | 2.76 ± 0.28 a |
| HLS | 13.84 ± 1.23 a | 102.60 ± 13.45 a | 23.80 ± 1.92 a | 56.03 ± 4.30 a | 61.95 ± 6.73 a | 2.82 ± 0.27 a | |
| WOA | 11.98 ± 0.67 bc | 103.40 ± 4.10 a | 21.20 ± 1.48 a | 48.77 ± 5.91 a | 53.40 ± 4.38 b | 2.67 ± 0.08 a | |
| ID | 11.40 ± 1.18 c | 64.60 ± 9.42 b | 17.00 ± 3.46 b | 17.34 ± 16.92 b | 32.05 ± 3.09 c | 0.77 ± 0.15 b |
| Tissue | Sampling | Treatment | H+-ATPase | PEPC | FRO |
|---|---|---|---|---|---|
| Leaf | 27 DAT | CA | 1.46 ± 0.18 b† | 482.3 ± 64.0 ab | 7.2 ± 0.2 b |
| HLS | 1.84 ± 0.07 ab | 267.1 ± 65.7 cd | 7.4 ± 0.1 b | ||
| WOA | 1.57 ± 0.05 ab | 633.6 ± 154.2 a | 7.2 ± 0.3 b | ||
| ID | 3.76 ± 2.22 a | 343.5 ± 56.9 bc | 9.6 ± 0.3 b | ||
| 49 DAT | CA | 0.92 ± 0.11 b | 834.2 ± 151.4 a | 7.7 ± 0.2 b | |
| HLS | 0.89 ± 0.04 b | 856.5 ± 140.6 a | 7.5 ± 0.3 b | ||
| WOA | 0.89 ± 0.05 b | 784.9 ± 194.0 a | 9.1 ± 0.4 b | ||
| ID | 0.65 ± 0.05 c | 552.7 ± 113.0 a | 6.7 ± 0.3 b | ||
| 84 DAT | CA | 2.65 ± 1.33 a | 465.6 ± 221.8 b | 10.7 ± 1.2 b | |
| HLS | 1.58 ± 0.06 a | 759.6 ± 136.5 a | 8.2 ± 0.2 bc | ||
| WOA | 1.25 ± 0.13 a | 141.0 ± 27.0 cd | 10.5 ± 1.1 b | ||
| ID | 1.81 ± 0.63 a | 304.0 ± 99.5 bc | 6.7 ± 1.1 c | ||
| Root | 27 DAT | CA | 0.016 ± 0.0005 b | 51.5 ± 29.7 e | 15.8 ± 2.1 a |
| HLS | 0.017 ± 0.0007 b | 95.2 ± 43.1 de | 17.2 ± 1.5 a | ||
| WOA | 0.017 ± 0.0004 b | 191.8 ± 16.0 cde | 17.3 ± 2.4 a | ||
| ID | 0.017 ± 0.0009 b | 156.9 ± 48.3 cde | 14.5 ± 1.3 a | ||
| 49 DAT | CA | 1.35 ± 0.05 a | 26.2 ± 21.5 b | 22.0 ± 1.2 a | |
| HLS | 1.28 ± 0.02 a | 11.6 ± 7.0 b | 23.2 ± 0.7 a | ||
| WOA | 1.26 ± 0.13 a | 16.6 ± 12.2 b | 22.2 ± 2.9 a | ||
| ID | 1.32 ± 0.05 a | 4.6 ± 0.1 b | 25.3 ± 1.5 a | ||
| 84 DAT | CA | 1.50 ± 0.05 a | 4.5 ± 0.0 d | 21.6 ± 0.9 a | |
| HLS | 1.27 ± 0.11 a | 205.6 ± 21.1 bcd | 20.1 ± 1.7 a | ||
| WOA | 1.29 ± 0.03 a | 11.8 ± 7.6 d | 21.4 ± 1.1 a | ||
| ID | 1.48 ± 0.04 a | 261.9 ± 16.5 bcd | 20.1 ± 1.8 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pérez-Labrada, F.; Benavides-Mendoza, A.; Juárez-Maldonado, A.; Solís-Gaona, S.; González-Morales, S. Effects of Citric Acid and Humic-like Substances on Yield, Enzyme Activities, and Expression of Genes Involved in Iron Uptake in Tomato Plants. Horticulturae 2023, 9, 630. https://doi.org/10.3390/horticulturae9060630
Pérez-Labrada F, Benavides-Mendoza A, Juárez-Maldonado A, Solís-Gaona S, González-Morales S. Effects of Citric Acid and Humic-like Substances on Yield, Enzyme Activities, and Expression of Genes Involved in Iron Uptake in Tomato Plants. Horticulturae. 2023; 9(6):630. https://doi.org/10.3390/horticulturae9060630
Chicago/Turabian StylePérez-Labrada, Fabián, Adalberto Benavides-Mendoza, Antonio Juárez-Maldonado, Susana Solís-Gaona, and Susana González-Morales. 2023. "Effects of Citric Acid and Humic-like Substances on Yield, Enzyme Activities, and Expression of Genes Involved in Iron Uptake in Tomato Plants" Horticulturae 9, no. 6: 630. https://doi.org/10.3390/horticulturae9060630
APA StylePérez-Labrada, F., Benavides-Mendoza, A., Juárez-Maldonado, A., Solís-Gaona, S., & González-Morales, S. (2023). Effects of Citric Acid and Humic-like Substances on Yield, Enzyme Activities, and Expression of Genes Involved in Iron Uptake in Tomato Plants. Horticulturae, 9(6), 630. https://doi.org/10.3390/horticulturae9060630