Abstract
The harm that invasive species cause to the environment has received a lot of attention. It is therefore appropriate that the current research was undertaken to evaluate the effects of invasion by Nicotiana glauca Graham on soil fertility by looking at (i) its contribution to the mycorrhizal potential of the soil, (ii) its impact on soil richness and diversity in terms of the arbuscular mycorrhizal fungi (AMF) community (iii), and its ability to modify the physicochemical characteristics in the invaded soil, specifically cleaning up heavy metal. The current study was conducted at Al Houz plain (Marrakesh region, Morocco), in heavily infested sites by N. glauca. The spores of AMF were isolated using the wet sieving process; the isolated spores were sorted for morphological features using a binocular microscope. The plant roots were thinned and colored before microscopic observation. The most probable number method was used to assess mycorrhizal soil infectivity. Heavy metal contamination in soils was characterized using an X-ray fluorescence spectrometer, and the pollution load index (PLI) was utilized to assess and compare the level of heavy metal contamination at each station. The ability of N. glauca to reproduce was evaluated in order to support one of its invasive characteristics. The estimate indicated that each plant might produce more than three million seeds. This significant number guarantees the plant a great capacity for reproduction and invasion. The extra-significant mycorrhizal potential, which can take the form of spores, mycelium, or vesicles that can regenerate mycorrhizae, was discovered by conducting soil analysis in the rhizospheric soils of N. glauca. This research demonstrated the strong mycotrophic capability of N. glauca and the large mycorrhizal potential of soils. Between 4.85 and 305.5 mycorrhizal propagules were considered to be the most probable number (MPN) per 100 g of dry soil. Based on color, shape and size, AMF were classified into five morphotypes corresponding to five genera. The isolated taxa of AMF with the most diverse spores were Glomus, Rhizophagus, Paraglomus, Scutellospora, and Sclerocystis. The Glomus genus was found to have spores in significant quantity. Furthermore, N. glauca demonstrated a potential involvement in the phytoremediation of damaged soils, with a high pollution load index demonstrating a particularly high accumulation of heavy metals. N. glauca is a highly mycotrophic plant that can boost soil mycorrhizal propagule stock. N. glauca has also been demonstrated to be a phytoremediation plant capable of cleansing contaminated soils. As a result, N. glauca could be considered as a prospective candidate for application in phytoremediation of polluted soils.
1. Introduction
Invasive plants are defined as species that can survive, reproduce, and spread across the landscape at alarming rates [1]. Invasive plants have evolved into one of the key elements of the Anthropocene as a result of the enormous, continuously expanding human population [2]. The composition, structure, and/or functioning of ecosystems are significantly altered due to their proliferation in natural or semi-natural settings [3] where they cause significant habitat transformation, resulting in the loss or reduction in biodiversity in ecosystems. Thus, they were often referred to as “transformative species” [4].
The tree tobacco Nicotiana glauca Graham (Solanaceae) is a fast-growing perennial evergreen shrub, native to South America in Argentina, Bolivia, Paraguay, and Uruguay [5]. Robert Graham recorded this plant in the Royal Botanic Garden in Edinburgh, where it was introduced from seeds transmitted from Buenos Aires [6]. It is currently considered as a naturalized species in most of the subtropics, including the Mediterranean basin, where it is widely naturalized in Macaronesia, parts of Europe, and Northern Africa [3,7]. This species is also distributed in the subtropical zone of the southern hemisphere in Australia [8] and in South Africa [9]. The circum-Mediterranean countries seem to be the oldest areas regarding the introduction of N. glauca. This species was introduced to the Canary Islands at the beginning of the nineteenth century, and due to its numerous tiny seeds, it is now widely distributed, even in the drier areas of Fuerteventura, Lanzarote, and Graciosa [10]. Tree tobacco has been widely naturalized throughout North Africa, particularly in Morocco, Algeria, Libya, and Egypt. For instance, [11] identified it as a naturalized plant in Algeria. As it was recorded in the Moroccan Plant Catalog, N. glauca was most likely introduced to Morocco before 1931 [12,13]. The tree tobacco is listed in the Global Invasive Species Database and has spread to Central America (Mexico, California, and the Channel Islands), Africa (Morocco, South Africa, and Namibia), Australia, and the Oceanic Islands (St. Helena) [14]. During the German occupation (1884–1914), it was introduced into Namibia in contaminated horse feed, from where it spread to South Africa [8]. N. glauca can grow up to 6 m tall, with loose-branched stems and bluish or greyish-green, pointy, elliptic to lanceolate, alternating leaves, as well as flowers that are 30–40 mm long, greenish to yellow, and some of which are borne in a lax panicle [6]. It features an egg-shaped fruit with a 7–10 mm long, two-valved capsule that produces a large number of tiny seeds that may be dispersed by the wind or water [6]. All plant parts are highly toxic, and numerous studies have shown that N. glauca is extremely toxic to humans, animals, and plants [15]. Despite numerous reports about the devastation wrought by invasive plant species, there are few quantitative data that assess the environmental and ecological impacts of most invasive plants, particularly N. glauca.
The significant growth and development of these plants in the face of extreme environmental conditions (soil pollution, water stress, salt stress, etc.) raise the question of whether the diversity and function of soil microorganisms are one of the factors implied in this progress, and, more importantly, on the symbiotic relationships formed in this biotope.
According to Pickett et al. [16] invasive plants influence the soil fungal community and, as a result, the structure of indigenous plant populations for years after the invasive plant has been eradicated from an area. Arbuscular mycorrhizal fungi (AMF) are mutually beneficial symbionts that interact with roughly 80% of terrestrial vascular plants, providing essentials such as an increased surface area for root water absorption and pathogen resistance [17]. This symbiotic relationship is governed by molecular and genetic signals between the fungus and the host plant, which benefits from growth-limiting nutrients such as phosphate and nitrate while providing carbohydrates formed by photosynthesis to the fungus [18]. Many researchers reported that AMF provide numerous benefits to their host plants, including increased plant growth and mineral nutrients, alleviation of plant abiotic stress, and increased plant resistance to pathogens [19,20]. As a result of the intermediation of plant–plant relations, AMF directly influences the performance of host plants and indirectly influences the diversity and composition of plant communities [21]. Furthermore, the identity and abundance of host plants affect the composition and diversity of AMF communities, demonstrating that interactions between plant species and AMF shape fundamental ecosystem properties. Mycorrhizal status and the role of mycorrhizal symbiosis in plant nutrition provide information on the degree to which plants rely on mycorrhizal symbiosis [21]. Invasive plants, on the other hand, can disrupt fungal mutualistic relationships with native plants once they are introduced into a healthy environment. Furthermore, these exotic plants can disturb mutualistic relationships by altering soil nutrient distribution and soil food connections or by introducing plant pathogens [22]. These changes affecting the mutualistic relationship between native plants and AMF are not always negative; the benefits brought on by exotic plants can last for years after their final removal and have been dubbed “legacy effects” [23]. The concept of legacy effects describes the abiotic and biotic changes caused by invasive species that persist long after they have been removed from an area [24]. Previous research in Morocco had classified N. glauca Graham as a ruderal plant that grows in disturbed or unstable open spaces [25]. Furthermore, it was regarded as a pioneer plant in disturbed areas highly susceptible to anthropogenic action, even capable of colonizing space following an upheaval or modification of ecosystems [26]. N. glauca grows in a wide range of environments, including wet rock gardens, river edges, asphalt roads, inaccessible cliffs, and even old walls and roofs [10,27]. Nègre described it in 1962 as a ruderal nitrophilic plant found only in a pile of rubble near the Koutoubia in Marrakesh. N. glauca is found up to 3700 m above sea level in warm temperate, arid and subtropical, and dry and humid regions [3,28]. In these areas, N. glauca is extremely adaptable to water scarcity and high temperatures [29]. The tree tobacco has successfully adapted to various Moroccan bioclimates and has become a widespread invasive plant in Morocco, particularly in Saharan Morocco, Middle Atlantic Morocco, North Atlantic Morocco, the Eastern Moroccan mountains, the Mediterranean coast, and the Rif [27]. N. glauca, on the other hand, cannot tolerate high salinity or prolonged flooding [30]. N. glauca’s partial tolerance to drought and salinity results from physiological adaptations such as stomatal closure and osmotic adjustment, as well as reduced absorption of excess radiation due to the presence of a waxy layer on leaves [27]. The legacy effects of the invasive plant N. glauca on native plants and the rhizosphere microbial community have not been well studied, and studies on AMF communities are particularly scarce.
In this context, and to better understand this plant’s symbiotic relationship, the current study was conducted to evaluate (i) the mycorrhizal status of N. glauca in degraded Mediterranean soils and its contribution to the mycorrhizal potential of the soil, (ii) the extent and type of AMF community associated with N. glauca rhizosphere soil, and (iii) the ability to modify the physicochemical and biological characteristics of the invaded soil, particularly cleaning up heavy metals.
2. Materials and Methods
2.1. Sampling Sites
This research was conducted at five stations located throughout the Al Houz plain (Marrakesh region, Morocco), which are heavily infested by N. glauca. Three stations are found along the banks of rivers. Two stations are found on the Oued Tensift river’s outskirts; downstream is Oued Tentsift-Safi, which represents the main infiltrated site (station 2) and upstream is Oued Tentsift-Casa (station 3). One station is located on the Oued Lahjar river’s bank (station 4) and two more stations are located in downtown Marrakesh, on Mount El Koudiate (Station1) and the Ourika road boards in the level of Lahebichate locality (station 5) (Figure 1).
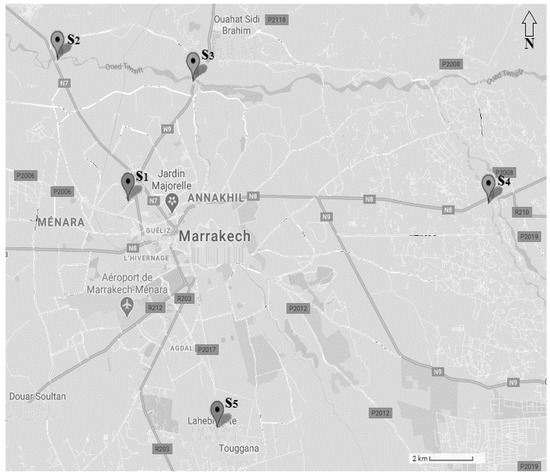
Figure 1.
Localzation of the sampling sites (S1: El Koudiate; S2: Oued Tensift-Safi; S3: Oued Tensift-Casa; S4: Oued Lahjar; S5: Lahebichate).
2.2. Soil Physicochemical Analysis
Soil pH and electric conductivity were measured in soil solution using a pH meter (Basic 20) and a conductometer (Basic 30). Total carbone and organic matter were measured using the Anne method. The mineral composition of soils was determined after mineralization at 600 °C for 6 h. The total nitrogen was determined according to the Kjeldahl digestion, distillation, and titration method [31]. Available phosphorus was determined by colorimetry [32] using a spectrophotometer at 820 nm. The content of potassium was determined by atomic absorption spectrophotometry [33].
2.3. Estimated Seed Production of N. glauca
An adult reproductive plant from each station was evaluated in order to estimate seed production by N. glauca. The plants’ capsules and flowers were all counted. Before dehiscence, five capsules were chosen at random, placed individually in Petri dishes, and dried for two hours at 65 °C. The seeds were counted under a binocular microscope after the dried capsules were manually exploded.
2.4. Soil Sampling and Enumeration of AMF Spores in Soils
To study the mycorrhizal status of N. glauca and the mycorrhizal soil infectivity, soil sampling was limited to areas directly influenced by the plant’s roots (rhizospheric soil) at a depth of 10 cm to 40 cm. In the meantime, control soils (bare soils) were randomly collected far from any influence of N. glauca roots. The spores of AMF were isolated using the method described by Brundrett et al. [34], which involves wet sieving of soil followed by centrifugation in sucrose solution (65%). To accomplish the enumeration of AMF spores, 100 g of rhizospheric soil were suspended in 1 L of tap water. After that, the mixture was stirred and allowed to settle. Following decantation, the supernatant was poured through a series of sieves ranging in size from 800 µm to 50 µm. The first sieve was fine enough to remove the largest organic matter particles while still allowing the desired spores to pass. The second was coarse enough to allow fine soil particles to pass through while retaining the desired spores. The procedure was repeated several times in order to recover as many spores as possible. The sieves were suspended in distilled water and centrifuged at 2000 rpm for 5 min. The centrifugation of water removed a significant amount of floating organic debris. The pellet was then suspended in 65% sucrose for the next step. The samples were then centrifuged at 2000 rpm for 1 min to separate the spores from the denser soil components. The supernatant was removed immediately after centrifugation and filtered under vacuum; the spores were collected on filter paper. Three repetitions were performed for each station. The total number of spores was counted directly under a binocular microscope to estimate the AMF community in soils.
2.5. Morphological Description of AMF Spores Associated with N. glauca
The isolated spores were manually sorted under a binocular microscope based on morphological characters, which are color, shape, and size. Spores with the same color form a homogeneous group or morphotype. The relative abundance of each morphotype was estimated as the proportion of this genus’ spores to all spores in the rhizospheric soil at each station.
2.6. Determination of Mycorrhizal Traits in the Roots of N. glauca
The roots were thinned and colored using a modified Phillips and Hayman [35] method. To remove the attached soil, organic matter, and foreign roots, the roots were washed in a gentle stream of water. For 60 min at 90 °C, the roots were bleached with a 10% KOH solution. After that, the samples were rinsed and acidified with a few drops of 5% lactic acid, which neutralized the remaining KOH. The thinned roots were stained for 15 min at 90 °C with a solution of 0.05% acidic trypan blue diluted in lactoglycerol (1/3 water, 1/3 glycerol, and 1/3 lactic acid). The colored roots were sandwiched between the slide and the coverslip and examined under a microscope. The number of mycorrhizal roots in the total examined roots is represented by the frequency of mycorrhization. The rate of mycorrhization provides information on the total volume of the root colonized by fungi when all fungal forms are considered. It was estimated based on five colonization classes: 0 (no mycorrhization), 1 (trace of mycorrhization), 2 (less than 10% colonization), 3 (between 11% and 50% colonization), 4 (between 51% and 90% colonization), and 5 (more than 91% colonization) [31,36]. The volume occupied by each different mycorrhizal structure was also estimated using microscopic observations (mycelium, vesicles, and arbuscules).
2.7. Mycorrhizal Soil Infectivity in the Rhizhospheric Soil of N. glauca
The most probable number test was used to estimate mycorrhizal soil infectivity (MPN). This method determines the number of propagules (spores, colonized root fragments, and fungal hyphae) capable of initiating mycorrhizal association per 100 g of soil [37]. Despite the fact that propagules counting is indirect, this method is presented as a solution to the problems encountered when using traditional methods of counting AMF propagules [38].
To carry out this test, various rhizospheric soil samples were collected from the five targeted stations as well as from bare soil (control) (S1: El Koudiate; S2: Oued Tensift-Safi; S3: Oued Tensift- Casa; S4: Oued Lahjar; S5: Lahebichate; CS: bare soil or control). Six dilutions were performed for each composite soil by carefully mixing this soil with a disinfected sandy soil (121 °C, 2 h) in proportions (1, 1/4, 1/16, 1/64, 1/256, 1/1024), with five repetitions for each dilution. The native mycorrhizal complex naturally associated with N. glauca was trapped using maize as an endophytic plant. Pre-germinated corn seeds were planted in plastic cups containing 100 g of soil dilution. The plants were seeded after four weeks of cultivation, and their root system was thinned, stained, and examined under a microscope. When the root system showed signs of infection, it was classified as positive, and when there was no colonization, it was classified as negative. The table of Fisher and Yates [39] was used to estimate the most probable number. The following formula was used to calculate the most probable number of propagules (MPN). Log10 MPN = x Loga − k. Where x is the mean number of infected plants, a is the dilution factor, and y = s − x with s being the number of dilutions; x and y are required to determine k in the Fisher and Yates table. The estimated standard deviation of Log 10 MPN is given by the following formula [40].
2.8. Heavy Metal Analysis in N. glauca Rhizhospheric Soils
Because of its dependability and speed of measurement, a portable X-ray fluorescence spectrometer (FPXRF—Field Portable X-ray Fluorescence) was used to characterize heavy metal pollution in soils. It is a non-destructive analytical technique for determining the qualitative and quantitative composition of a soil sample. The basic idea behind XRF fluorescence is to excite the atoms in the sample to be analyzed with “primary” X-rays, which then emit “secondary” X-rays. The X-rays emitted are indicative of the sample’s elementary atomic composition as well as the mass concentration of each element. Metallic trace element (MTE) concentrations measured at various stations were compared to normal world averages of uncontaminated soils provided by Bowen [41].
2.9. The Pollution Load Index
The pollution load index (PLI) was used to assess and compare the level of heavy-metal pollution at each station. The PLI was calculated for each site individually, as reported by Tomlinson et al. [42]. . n represents the total number of trace metallic elements in the soil. CFi is the concentration factor of each metallic trace element in soil, defined as the ratio of each MTE concentration in a given soil to the MTE’s natural concentration in soil (geochemical background).
The natural MTE concentrations used are provided by Bowen [41]. A pollution load index (PLI) greater than one indicates polluted soil, whereas a PLI less than one indicates that the targeted soil is not polluted by the considered MTE.
2.10. Statistical Analysis
In all assays, three sample replicates were performed, and the mean values were estimated together with the standard deviation (SD). Data were processed with analysis of variance (ANOVA) at a threshold of statistical significance set at 5% using the XL Stat software. Simultaneously, Tukey’s HSD test was used to compare the means to demonstrate significant statistical differences in all variables. Then, Spearman rank correlation coefficients were calculated to examine relationships between root colonization, the number of spores from individual species, and soil characteristics. Principal components analysis (PCA) was performed between AMF spore populations and some soil properties to determine which factors were responsible for the variability in spore number.
3. Results
3.1. Physicochemical Characteristics of the Rhizospheric Soil of N. glauca and Bare Soil
A physicochemical comparison of N. glauca-affected rhizospheric soils with unaffected soils revealed noticeable impacts of the plant’s establishment in these soils. Together with soil enrichment in organic matter, total nitrogen, available phosphorus, and potassium, extensive soil acidity was also exhibited. Similarly, the C/N ratio measurement demonstrated that there was high biological activity in the rhizospheric soils under N. glauca (Table 1).

Table 1.
Physico chemical traits in rhizospheric soils of N. glauca plants and bare soils (Control CS) in the targeted sites.
3.2. Estimate of N. glauca Seeds Production
At each of the five locations studied the estimated average seed production from an adult reproductive plant of N. glauca revealed 3000 ± 127 fruits with 1054 ± 216 seeds per fruit. As a result, it is generally assumed that a single plant is capable of producing between three and four million seeds throughout its lifetime.
3.3. Counting the Spores of AMF Associated with N. glauca
The presence of AMF spores in the rhizospheric soil of N. glauca was detected in all analyzed samples. The total number of spores per 100 g varied significantly between stations, with a very large total number of spores in the case of Lahebichate (S5) (700 spores) and El Koudiate station (S1) (600 spores), which represent the city suburb, whereas the abundance of spores appeared to be lower in the stations located along the rivers’ edges, with a density of 350 spores at the level of the Oued Lahjar river’s bank (S4) and 180 spores in the case of Tensift-Casa station (S3). The lowest spore density was observed in the rhizospheric soil in Oued Tensift-Safi station (S2) (130 spores/100 g), which is five times lower than at El Koudiate station (Figure 2).
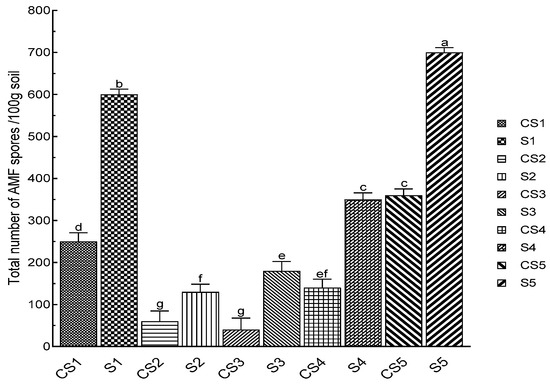
Figure 2.
The total number of arbuscular mycorrhizal fungi spores in the rhizospheric soils of N. glauca in the targeted sites. The bar charts (means ± SD) indexed by the same small letters are not significantly different according to Tukey’s test (p < 0.05). (S1: El Koudiate; S2: Oued Tensift-Safi; S3: Oued Tensift-Casa; S4: Oued Lahjar; S5: Lahebichate; CS: bare soil in different stations).
Following the establishment of N. glauca, considerable soil enrichment in fungal spores was observed in the stations under study. For instance, there were notable changes between the bare soil (CS) far from N. glauca ’s plant involvement and the rhizospheric soil. In comparison to bare soils, the number of isolated spores is four times higher in rhizospheric soils (Figure 2).
3.4. Morphological Differentiaition of AMF Spores Morphotypes Associated with N. glauca
The relative abundance of various spore morphotypes was used to describe the organization of the N. glauca-associated AMF population. At least five distinct morphotypes were visible when the spores were examined under a binocular magnifier at high magnification. These morphotypes varied in color, shape, and size. Figure 2 displays the number of spores at each studied site, and Figure 3 compares the relative abundance of each morphotype at the various sites (Figure 3).
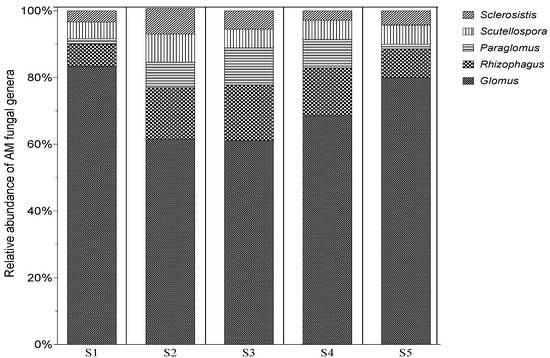
Figure 3.
The relative abundance (RA) of the main AM fungal genera in the rhizosphere soils of N. glauca at five sampling sites in Morocco. RA (%) was estimated as the proportion of a recognized genus’ spores to all spores in the rhizospheric soil at each station (S1: El Koudiate; S2: OuedTensift-Safi; S3: OuedTensift-Casa; S4: Oued Lahjar; S5: Lahebichate).
The findings indicated that Glomus was the morphotype genus that was most prevalent across all stations (76%), followed by Rhizophagus (10%), Scutellospora (6%), Paraglomus (4%) and Sclerocystis (4%).
3.5. Determination of Mycorrhizal Traits in N. glauca Plant Roots
Microscopic observation of the roots of N. glauca plants in the various selected sites showed that all the plants exhibited AMF colonization. However, the mycorrhizal colonization assessment showed significant differences between the roots of the plants at the different sites. The mycorrhial colonization rate is considerable in site 1 (El Koudiate, 62%) and in site 5 (Lahebichate, 50%), while the sites on the river banks had shown relatively lower rates (site 4, Oued Lehjer, 38%; Site 2 and 3, Oued Tensift-safi and Oued Tensift-casa 25%). The different structures of the symbiotic association (arbuscules, vesicles, hyphae) were constantly present with varying rates in all the examined roots (Table 2).
Table 2.
Substantial mycorrhizal traits in roots of N. glauca plants in the targeted sites.
3.6. Mycorrhizal Potential in Rhizospheric Soils of N. glauca (MPN)
The results on the mycorrhization of the roots of corn seedlings cultivated on the different dilutions of the considered rhizospheric soils of N. glauca and non-rhizospheric soil (bare soil/control) were transformed, using the model and the table of Fisher and Yates [35], in several fungal propagules in 100 g of soil capable of regenerating a symbiosis. Plants that presented at least one point of infection were considered mycorrhizal plants. The results showed that the corn seedlings planted in the control soils were slightly infected by mycorrhizal fungi, which provides information on the poverty of these soils in fungal propagules. On the other hand, the roots of corn plants grown in rhizospheric soils had shown infections with high intensities. For the rhizospheric soils of the El Koudiate (S1) and Lahebichate (S5) stations, all the plants cultivated in the diluted or undiluted rhizospheric soil were infected by AMF (100% infection). Seedlings grown in rhizospheric soils associated with the Oued Tensift-Safi (S2), Oued Tensift-Casa (S3), and Oued Lahjar (S4) rivers had shown relatively moderate mycorrhizal infections. The most probable number of mycorrhizal propagules per 100 g of rhizospheric soil was estimated to be 305.5 at stations S1 and S5, 175.79 at station S4, 44.15 at S3, 19.27 at S2, and finally only 4.85 propagules at the control soil CS2 (Figure 4).
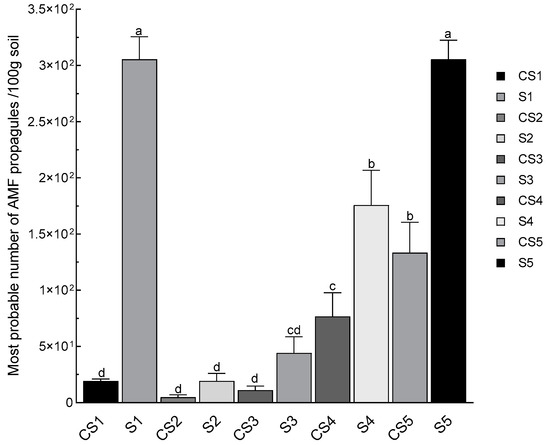
Figure 4.
The most probable number of fungal propagules in rhizospheric soils of N. glauca in the targeted stations. (S1: El Koudiate; S2: Oued Tensift-Safi; S3: Oued Tensift-Casa; S4: Oued Lahjar; S5: Lahebichate; CS: bare soil non-rhizospheric in different stations). The bar charts (means ± SD) indexed by the same small letter are not significantly different according to Tukey’s test (p < 0.05).
3.7. Heavy Metal Analysis and Pollution Load Index (PLI)
The effects of an N. glauca invasion on soil pollution were assessed in order to test the hypothesis that this plant is capable of rehabilitating degraded soils, especially those polluted by metallic trace elements. The pollution load index showed that the downtown station (El Koudiate S1) is the only one with a PLI that exceeds 1, and consequently it is the only station where pollution has been confirmed (Figure 5). The proximity to human activities explains the pollution in this station. In addition, the PLI in the rhizospheric soil of N. glauca is significantly lower compared to the non-rhizospheric soil in two stations (El Koudiate and Lahebichate), which ensures the capacity of N. glauca to accumulate MTEs by absorption from the soil. Figure 5 showed in a more explanatory way the results obtained by the PLI. Station S1 is assessed as the most polluted station by PLI; this is explained by the existence of MTE (Zn, Cu, As, Mo) at concentrations above ordinary soil levels [38]. Certain METs, such as zinc, lead, copper, and arsenic, are found at all stations, although only arsenic is present at a higher concentration than in ordinary soils at all stations (Figure 6).
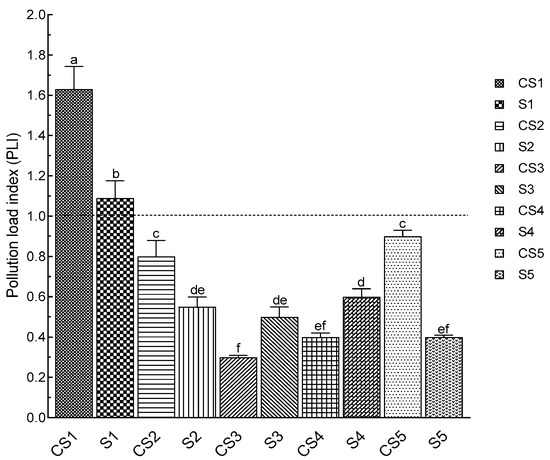
Figure 5.
The pollution load index (PLI) of non-rhizospheric (CS) and rhizospheric (S) soils of N. glauca in the targeted stations. (S1: El Koudiate; S2: Oued Tensift-Safi; S3: Oued Tensift-Casa; S4: Oued Lahjar; S5: Lahebichate). The bar charts (means ± SD) indexed by the same small letter are not significantly different according to Tukey’s test (p < 0.05).
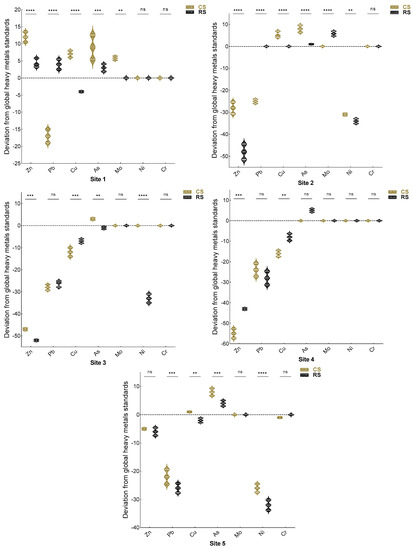
Figure 6.
Effect of N. glauca on deviations from global averages of normal heavy metals in uncontaminated soils given by Bowen (1979). For each heavy metal assessed, violin plots (means ± SD) with asterisks indicate significant differences between rhizospheric and non-rhizospheric soil (α = 0.05), as determined using Student’s t-test (** p < 0.01; *** p < 0.001; **** p < 0.0001 and ns, no-significance p > 0.05). (S1: El Koudiate; S2: Oued Tensift-Safi; S3: Oued Tensift-Casa; S4: Oued Lahjar; S5: Lahebichate). [Zn = zinc, Pb = lead, Cu = copper, As = arsenic, Mo = molybdenum, Ni = nickel and Cu = chromium].
3.8. Inter-Dependencies between all Studied Variables
The results of the principal component analysis (PCA) revealed that the first three PCs described the majority of the data variance (79.86%), with the first two contributing the most (Figure 7A). The PC1, which accounts for 45.75% of the overall variability, was highly linked with the EC, OM%, TC%, N%, P, K, MPN, amount of AMF spores, MF%, MC%, V%, and A% and represented a clear group. The following variables had high loadings for the second PC: H%, MF%, Cr, As, Zn, and C/N, with the latter three being negatively linked with this dimension. The physicochemical parameters and AMF features of rhizospheric soil and bare soil samples collected from five different geographic sites in Morocco varied considerably, as shown by the PCA biplot in Figure 7A. Likewise, the investigated physicochemical measurements, except pH values, had positive correlations with each other and with AMF variables, except for the H% (Figure 7A). For the examination of metallic trace elements (MTEs) in the soil of N. glauca, Pearson’s correlation coefficient (r) analysis revealed significant strong/moderate relationships between the following elements: Zn-C/N (r = 0.56, p = 0.001); Zn-pH (r = −0.81, p = 0.001); Zn-P (r = 0.55, p = 0.001); Zn-MPN (r = 0.51, p = 0.009); Zn-Cu (r = 0.64, p = 0. 001); Zn-As (r = 0.65, p = 0.001); As-Cr (r = 0.714, p = 0.05); Pb-MC% (r = 0.43, p = 0.018); Pb-V% (r = 0.43, p = 0.017); As-C/N (r = 0.54, p = 0.002); As-MF% (r = −0.50, p = 0.011); As-H% (r = −0.55, p = 0.001); Mo-MPN (r = −0.42, p = 0.028); Mo-Pb (r = 0.49, p = 0.007); Mo-Cu (r = 0.51, p = 0.005); Ni-H% (r = −0.40, p = 0.031); and Cr-pH (r = −0.53, p = 0.003). In general, negative medium-to-low correlations were discovered between MTEs (deviations from the global MTEs standard) and the majority of physicochemical characteristics and mycorrhizal fungal traits (Figure 7B).
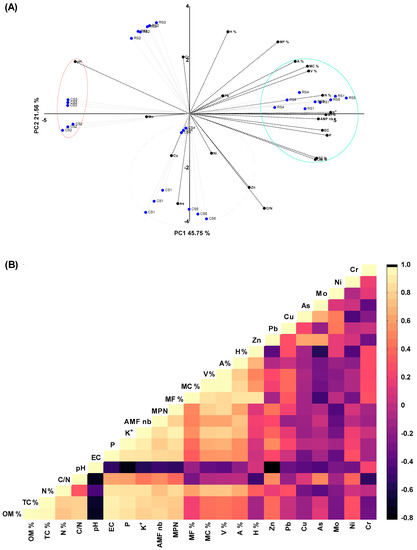
Figure 7.
Multivariate statistical analysis of mycorrhizal characteristics and physicochemical features in rhizospheric and bare soils of N. glauca plants: biplot of principal component analysis (PCA) (A) and heatmap of the Pearson correlation matrix (B).
4. Discussion
In the current study, the tree tobacco, N. glauca, showed production of an enormous number of seeds per plant at the study site. This figure is likely to rise as flowering and fruiting occur in Morocco three times a year, in the spring, summer, and fall [27]. Estimates of N. glauca seed production are hazy based on observations from various sources [43]. For instance, adults in the Canary Islands can produce between 10,000 and 1,000,000 seeds per year [33], while in Brazil, Fabricante et al. [44] noticed an average of 2121 ± 940 fruits per plant and 644 ± 50 seeds per fruit, i.e., 700,000 to 2,100,000 seeds per plant. In Namibia, the average yield is 700 to 1300 seeds per fruit and 2500 fruits per plant, for a total yield of approximately 1,750,000 to 3,250,000 seeds per plant [45]. This large production of tiny seeds with near-perfect germination rates, even in the face of limiting environmental factors [46], explains N. glauca’s invasion potential. Seeds can germinate in temperatures ranging from 7 °C to 30 °C during all four seasons, even in areas with high soil salinity, and soil pH does not appear to be a limiting factor [46]. These distinguishing characteristics give N. glauca an advantage over native plants in saline conditions in arid and semi-arid environments. Water transport is most likely the most effective long-distance natural dispersal mode [7,33]. Furthermore, animals have been reported to act as local, secondary dispersants [47]. In addition, N. glauca was purposefully introduced as an ornamental plant, resulting in its spread both nationally and internationally [5].
The AMF spore densities estimated in rhizospheric soils from river banks (the station on the bank of the Oued Lahjar valley and the two others on the border of the Tensift river) were lower than the densities recorded in the other invaded stations in the Marrakesh city suburbs (El Koudiate and Lahebichate). Low spore densities at river mouths may be due to these soils’ sandy and silty texture. In general, sandy soils have lower levels of fungal spores than other soil textures [48]. Furthermore, these areas are waterlogged several times a year, regardless of season, due to torrential rains and floods. According to morphological analysis, the number of unique morphotypes among the many AMF spore types connected to N. glauca has revealed significant variety. At least five different morphotypes could be distinguished based on the color, the shape and the size of the spores. In fact, the morphological technique has demonstrated its limitations in describing the variety of mycorrhizal fungus in earlier investigations in Mediterranean ecosystems [37]. El Mrabet et al. [49] identified five distinct morphotypes under Euporbia beaumierana, while Ouahmane et al. [50] differentiated nine morphotypes of AMF linked with Cupressus atlantica. Research that made use of morphological features frequently only looked at Genus rank. Glomus, Rhizophagus, Acaulospora, Entrophospora, Scutellospora, and Gigaspora were the principal genera with recorded morphotypes [51].
The study of N. glauca Graham’s mycorrhizal traits revealed that this exotic species is highly mycotrophic. Its dense root system is heavily colonized by AMF, and at the end of the symbiotic fungi development cycle, the high sporulation enriches the environment with spores and other forms of propagules. The Most Probable Number (MPN) method of estimating the infectious potential of the soil revealed that non-rhizospheric soils are deficient in fungal propagules capable of generating a mycorrhizal symbiosis, which provides information on the weak biological fertility of these soils. Numerous studies have revealed that soil mycorrhizal potential is extremely low under degradation conditions [48,52]. However, the rhizospheric soils of land stations (S1 El Koudiate, S4 Lahebichate) exhibited a very high mycorrhizal infectious potential, with up to 305.5 propagules per 100 g recorded. Previous spore count results revealed the presence of 700 spores in the rhizospheric soil, indicating that spores are not the only propagules capable of colonizing the root system. This difference is primarily due to the presence of propagules other than spores, specifically mycelia fragments, vesicles, and mycorrhizal roots, which increase the soil’s mycorrhizal potential [52]. Even though they contain more spores, the soils near the rivers have a low mycorrhizal infectious potential (between 19.27 and 44.15 propagules per 100 g). On the one hand, the effect of soil disturbance on spore viability can explain the negative correlation found between MPN and spore density. The same result was confirmed by Diop et al. [53], who demonstrated that disturbed ecosystem soils contain many non-viable spores that are not detected by the MPN method [54]. This discovery was also made in the soil of Retama monosperma following the degradation of Algeria’s coastal ecosystem due to the overexploitation of coastal sands [55]. Despite having the most spore-rich soil, Retama monosperma had the lowest mycorrhizal infectious potential of the studied vegetation. This negative correlation, on the other hand, can be linked to many problems associated with the MPN method, such as the effects of the length and temperature of the host plant’s growth period, the specificity of the fungi towards the host, edaphic variables, and dormancy in some species [54].
This research has demonstrated that N. glauca is a highly mycotrophic plant capable of increasing the stock of mycorrhizal propagules, which is an effective source of mycorrhizal inoculum in highly disturbed and degraded soils with low mycorrhizal potential. As a result, wild tobacco is regarded as an ideal plant for rehabilitating degraded soils by managing mycorrhizal infectious potential. This mycotrophic property may be due to the positive interaction between N. glauca and the AMF community in soils. The positive feedback mechanisms regulate the production of beneficial secondary metabolites for AMF [56], and N. glauca can modify the environment created by the mycorrhizal symbiosis around the plant roots, dubbed the mycorrhizosphere, by fostering its growth and development on degraded soils. Such a positive reaction between plants and soil-based microorganisms such as AMF is crucial in the distribution of plant communities at an early stage of ecological succession [57], especially in degraded soils. As a result, tree tobacco is a pioneer plant that colonizes disturbed and ruderal areas [25,26]. Due to the mycotrophic property highlighted in the current study, N. glauca demonstrates that it could play critical roles in plant successions in degraded soils. Indeed, it has been reported that very mycotrophic pioneer species settle at the beginning of plant succession on degraded soils and then promote the development of other plant species via a “nurse plant” effect [48,58]. Previous research has highlighted the significance of nurse plants as vectors of mycorrhizal fungi propagation, which is regarded as a critical factor in all strategies focusing on the rehabilitation of degraded soils, particularly in Morocco, to optimize the performance of reforestation programs using high mycotrophic plants.
By assessing heavy metal levels in soils and determining the accumulation rate using the pollution load index (PLI), the current study discovered that N. glauca might accumulate Metallic Trace Elements in its tissues. Previous research has confirmed N. glauca Graham’s phytoremediation ability. Barazani et al. [59] discovered that N. glauca is the only perennial shrub that grows in a solid waste-contaminated site in Israel’s Negev desert. The concentrations of the seven heavy metals found at this contaminated site (Cu, Fe, Mn, Zn, Ni, Cd, and Pb) were three to 43 times higher than at the uncontaminated site, with zinc (2027 ppm), manganese (628 ppm), and lead (329 ppm) being particularly high. N. glauca accumulated more Cu, Zn, and Fe in the roots and stem than in plants from an uncontaminated site. Controlled experiments have also confirmed this plant’s hyperaccumulation potential. Zinc concentrations in aerial and root parts of N. glauca grown in contaminated soils were 9.5 and 4.1 times higher, respectively, than in plants grown in control soil, whereas the copper concentration in the roots of plants grown in contaminated soil was significantly higher than that of plants grown in control soil. Plant growth was significantly slowed in contaminated soils. However, no stress symptoms such as necrosis, wilting, or chlorosis were observed, indicating that N. glauca is resistant to metallic contaminants. The presence of a metal tolerance gene (MTP) is one of the heavy metal tolerance mechanisms demonstrated in N. glauca [60]. When this gene is transferred into yeast, it completes Zn and Co tolerance. This gene’s encoded proteins have been found to be located on vacuolar membranes. These findings suggest that this MTP works by sequestering Zn and Co in vacuoles, reducing their toxicity. In addition, N. glauca Graham was chosen for biotechnological modification because it demonstrated the best physiological characteristics for phytoremediation among a number of wild plant species growing on soils contaminated with hazardous industrial wastes in eastern Spain [61]. Tolerance to metals such as Pb and Cd increased dramatically after Agrobacterium-mediated transformation, with roots 160% longer than in wild-type plants. Furthermore, transformed plants grown in mining soils with high concentrations of Pb (1572 ppm) accumulated twice as much of this heavy metal as the wild type. These findings suggest that transformed N. glauca is a very promising new tool for phytoremediation. Due to its powerful root system capable of trapping metals from deep and weak soils, nutrient requirements adaptation to a wide geographic distribution around the world, rapid growth and high biomass production, resistance to drought and heavy metals, repellent property for herbivores which prevents its entry into the food chain, easy harvesting, and important biotechnological properties, N. glauca is universally regarded as the ideal plant for phytoremediation of contaminated soils. However, because it produces a large number of seeds, Gisbert et al. [61] recommend controlling the reproductive system to limit spread beyond the repair area. Furthermore, N. glauca is widely cultivated as an ornamental plant; its use in public gardens contributes to the reduction in lead and other byproducts of gasoline combustion in many cities’ streets and roads.
According to the current study and others, N. glauca is a phytoremediation plant capable of accumulating metals and thus cleaning the environment through intrinsic mechanisms. The association of N. glauca with soil fungal symbionts improves the restorative ability of this invasive species by increasing the plant’s effective absorption surface. In degraded and poor soils under extreme conditions, AMF hyphae of Mycorrhized roots extend beyond the exhaustion zone surrounding the roots, resulting in improved heavy metal uptake and better plant development.
5. Conclusions
The survey carried out in the areas invaded by N. glauca Graham in the Marrakesh-Safi region allowed us to draw interesting conclusions about how these plants affect their invaded habitats. These findings made it possible to accept the study’s hypothesis, which states that this plant has a positive impact on invaded areas. This research has demonstrated that N. glauca is a highly mycotrophic plant that can increase the stock of mycorrhizal propagules in rhizospheric soils, making it a potential and effective source of mycorrhizal inoculum in these disturbed and degraded zones with low mycorrhizal potential. Furthermore, N. glauca has been shown to be a phytoremediation plant capable of cleaning polluted soils. These accumulative properties, combined with other fascinating morphological, physiological, and genetic traits, make this species a promising candidate to be used in the phytoremediation of polluted soils. However, in order to consider N. glauca as a key plant in the rehabilitation of degraded soils without the risk of this invasion on natural environments, a more comprehensive multidirectional approach on the pathway of a better valuation of this invasive species must be identified.
Author Contributions
Writing original draft, preparation: H.D. and M.B. (Mohammed Bouskout) Review and editing, data curation: H.-A.N., A.A.A. and M.B. (Mohammed Bourhia) Conceptualization, supervision, writing, review, and editing: L.O. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Researchers Supporting Project number (RSP-2023/236), King Saud University, Riyadh, Saudi Arabia.
Data Availability Statement
The data are included in the article.
Acknowledgments
The authors acknowledge Researchers Supporting Project number (RSP 2023/236), King Saud University, Riyadh, Saudi Arabia for financial support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- van Wilgen, B.; Richardson, D.; Higgins, S.I. Integrated control of invasive alien plants in terrestrial ecosystems. L Use Water Resour. Res. 2001, 1, 1732-2016-140256. [Google Scholar]
- Usma, A.; Ahmad, M.; Zafar, M.; Sultana, S.; Ullah, F.; Saqib, S.; Ayaz, A.; Zaman, W. Palynological Study of Weed Flora from Potohar Plateau. Agronomy 2022, 12, 2500. [Google Scholar] [CrossRef]
- Cronk, Q.C.B.; Fuller, J.L. Plant Invaders: The Threat to Natural Ecosystems; Earthscan Publications Ltd. Routledge: London, UK, 2014; ISBN 1134203586. [Google Scholar] [CrossRef]
- Richardson, D.M.; Pyšek, P.; Rejmánek, M.; Barbour, M.G.; Panetta, F.D.; West, C.J. Naturalization and invasion of alien plants: Concepts and definitions. Divers. Distrib. 2000, 6, 93–107. [Google Scholar] [CrossRef]
- Ollerton, J.; Watts, S.; Connerty, S.; Lock, J.; Parker, L.; Wilson, I.; Schueller, S.; Nattero, J.; Cocucci, A.A.; Izhaki, I. Pollination ecology of the invasive tree tobacco Nicotiana glauca: Comparisons across native and non-native ranges. J. Pollinat. Ecol. 2012, 9, 85–95. [Google Scholar] [CrossRef]
- Jameson, R.; Jardine, W.; Rogers, H.D. The Edinburgh New Philosophical Journal: Exhibiting a View of the Progressive Discoveries and Improvements in the Sciences and the Arts; A. and C. Black: London, UK, 1841; Volume 30. [Google Scholar]
- Bogdanović, S.; Mitić, B.; Ruščić, M.; Dolina, K. Nicotiana glauca Graham (Solanaceae), a new invasive plant in Croatia. Acta Bot. Croat. 2006, 65, 203–209. [Google Scholar]
- Florentine, S.K.; Westbrooke, M.E.; Gosney, K.; Ambrose, G.; O’Keefe, M. The arid land invasive weed Nicotiana glauca R. Graham (Solanaceae): Population and soil seed bank dynamics, seed germination patterns and seedling response to flood and drought. J. Arid Environ. 2006, 66, 218–230. [Google Scholar] [CrossRef]
- Steenkamp, P.A.; Van Heerden, F.R.; Van Wyk, B.-E. Accidental fatal poisoning by Nicotiana glauca: Identification of anabasine by high performance liquid chromatography/photodiode array/mass spectrometry. Forensic Sci. Int. 2002, 127, 208–217. [Google Scholar] [CrossRef]
- Kunkel, G. Biogeography and Ecology in the Canary Islands; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2012; Volume 30, ISBN 940101566X. [Google Scholar]
- Quezel, P.; Santa, S. Nouvelle Flore de l’Algérie et des Régionsdésertiques et Méridionales, Tome II; Edition CNRS: Paris, France, 1963. [Google Scholar]
- Jahandiez, E.; Maire, R. Catalogue Des Plantes Du Maroc:(Spermatophytes et Ptéridophytes). Tome Premier. Ptéridophytes, Gymnospermes et Monocotylédones; Minerva: Alger, Algeria, 1931; 150p. [Google Scholar]
- Taleb, A.; Bouhache, M. Etat actuel de nosconnaissances sur les plantesenvahissantes au Maroc. Invasive Plants Mediterr. Type Reg. World 2006, 99–107. [Google Scholar]
- Schueller, S.K. Self-pollination in island and mainland populations of the introduced hummingbird-pollinated plant. Nicotiana glauca (Solanaceae). Am. J. Bot. 2004, 91, 672–681. [Google Scholar] [CrossRef]
- Furer, V.; Hersch, M.; Silvetzki, N.; Breuer, G.S.; Zevin, S.; Zevin, S. Nicotiana glauca (Tree Tobacco) Intoxication—Two Cases in One Family. J. Med. Toxicol. 2011, 7, 47–51. [Google Scholar] [CrossRef]
- Pickett, B.; Maltz, M.; Aronson, E. Impacts of invasive plants on soil fungi and implications for restoration. In Diversity and Ecology of Invasive Plants; IntechOpen: London, UK, 2019; pp. 1–18. [Google Scholar]
- Sally, E.; Smith, D.J.R. Mycorrhizal Symbiosis; Academic Press: Cambridge, MA, USA, 2008; Volume 137, ISBN 9780123705266. [Google Scholar]
- Kapoor, R.; Sharma, D.; Bhatnagar, A.K. Arbuscular mycorrhizae in micropropagation systems and their potential applications. Sci. Hortic. 2008, 116, 227–239. [Google Scholar] [CrossRef]
- Zardak, S.G.; Dehnavi, M.M.; Salehi, A.; Gholamhoseini, M. Effects of using arbuscular mycorrhizal fungi to alleviate drought stress on the physiological traits and essential oil yield of fennel. Rhizosphere 2018, 6, 31–38. [Google Scholar] [CrossRef]
- El Kinany, S.; Achbani, E.; Faggroud, M.; Ouahmane, L.; El Hilali, R.; Haggoud, A.; Bouamri, R. Effect of organic fertilizer and commercial arbuscular mycorrhizal fungi on the growth of micropropagated date palm cv. Feggouss. J. Saudi. Soc. Agric. Sci. 2019, 18, 411–417. [Google Scholar] [CrossRef]
- Neuenkamp, L.; Moora, M.; Öpik, M.; Davison, J.; Gerz, M.; Männistö, M.; Jairus, T.; Vasar, M.; Zobel, M. The role of plant mycorrhizal type and status in modulating the relationship between plant and arbuscular mycorrhizal fungal communities. New Phytol. 2018, 220, 1236–1247. [Google Scholar] [CrossRef] [PubMed]
- Ehrenfeld, J.G. Ecosystem consequences of biological invasions. Annu. Rev. Ecol. Evol. Syst. 2010, 41, 59–80. [Google Scholar] [CrossRef]
- Kulmatiski, A.; Beard, K.H. Long-term plant growth legacies overwhelm short-term plant growth effects on soil microbial community structure. Soil Biol. Biochem. 2011, 43, 823–830. [Google Scholar] [CrossRef]
- Cuddington, K. Legacy effects: The persistent impact of ecological interactions. Biol. Theory 2011, 6, 203–210. [Google Scholar] [CrossRef]
- Ater, M.; Ennabili, A. Flore (“Pteridophyta” et “Spermatophyta”) des zones humides du Maroc Méditérranéen: Inventaire et écologie. Acta Bot. Malacit. 1996, 21, 221–239. [Google Scholar]
- Bammi, J.; Mouhiddine, M.; Fassi, D.; Douira, A. Contribution à la connaissance de la végétation des Doukkala-Abda (Maroc Atlantique): Approcheéco-géomorphologique. J. Anim. Plant Sci. 2014, 20, 3202–3211. [Google Scholar]
- Fennane, M.; Tattou, M.I. Flore vasculaire du Maroc: Inventaire et chorologie. Trav. Inst. Sci. Rabat Sér. Botanique 2005, 1, 483. [Google Scholar]
- González, A.; Tezara, W.; Rengifo, E.; Herrera, A. Ecophysiological responses to drought and salinity in the cosmopolitan invader Nicotiana glauca. Brazilian J. Plant Physiol. 2012, 24, 213–222. [Google Scholar] [CrossRef]
- Curt, M.D.; Fernández, J. Production of Nicotiana glauca RC Graham aerial biomass in relation to irrigation regime. Biomass 1990, 23, 103–115. [Google Scholar] [CrossRef]
- Brandes, D. Nicotiana glauca als invasive Pflanze auf Fuerteventura. Adventivpflamen. Beiträgezu Biol. Vor. und Ausbreitungsdynamik von Archäophyten und NeophytenMitteleuropa. Tag. Des Braunschw. Kolloqu. Vom 2001, 3, 39–57. [Google Scholar]
- Kjeldahl, J. A New Method for the Determination of Nitrogen in Organic Matter. Z. Für Anal. Chem. 1883, 22, 366–382. [Google Scholar] [CrossRef]
- Olsen, R.S.; Watanabe, F.S. A Method to Determine a Phosphorous Adsorption Maximum of Soils as Measured by Langmuir Isotherms. Soil Sci. Soc. Am. J. 1957, 21, 144–149. [Google Scholar] [CrossRef]
- Gaines, T.P.; Mitchell, G.A. Boron determination in plant tissue by the azomethine H. method. Commun. Soil Sci. Plant Anal. 1979, 10, 1099–1108. [Google Scholar] [CrossRef]
- Brundrett, M.; Bougher, N.; Dell, B.; Grove, T.; Malajczuk, N. Working with mycorrhizas in forestry and agriculture (ACIAR Monograph 32). Canberra Aust. Aust. Cent. Int. Agric. Res. 1996, 374. [Google Scholar]
- Phillips, J.M.; Hayman, D.S. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Trans. Br. Mycol. Soc. 1970, 55, 158-IN18. [Google Scholar] [CrossRef]
- McGonigle, T.P.; Miller, M.H.; Evans, D.G.; Fairchild, G.L.; Swan, J.A. A new method which gives an objective measure of colonization of roots by vesicular—Arbuscular mycorrhizal fungi. New Phytol. 1990, 115, 495–501. [Google Scholar] [CrossRef]
- Ouahmane, L.; Hafidi, M.; Plenchette, C.; Kisa, M.; Boumezzough, A.; Thioulouse, J.; Duponnois, R. Lavandula species as accompanying plants in Cupressus replanting strategies: Effect on plant growth, mycorrhizal soil infectivity and soil microbial catabolic diversity. Appl. Soil Ecol. 2006, 34, 190–199. [Google Scholar] [CrossRef]
- Porter, W.M. The ‘Most Probable Number’ method for enumerating infective propagules of vesicular arbuscular mycorrhizal fungi in soil. Soil Res. 1979, 17, 515–519. [Google Scholar] [CrossRef]
- Fisher, R.A.; Yates, F. Statistical Tables: For Biological, Agricultural and Medical Research; Oliver and Boyd: Edinburgh, UK, 1938. [Google Scholar]
- Cochran, W.G. Estimation of bacterial densities by means of the “most probable number”. Biometrics 1950, 6, 105–116. [Google Scholar] [CrossRef] [PubMed]
- Bowen, H.J.M. Environmental Chemistry of the Elements; Academic Press: Cambridge, MA, USA, 1979; ISBN 0121204502. [Google Scholar]
- Tomlinson, D.L.; Wilson, J.G.; Harris, C.R.; Jeffrey, D.W. Problems in the assessment of heavy-metal levels in estuaries and the formation of a pollution index. Helgoländer Meeresunters. 1980, 33, 566–575. [Google Scholar] [CrossRef]
- Youssef, S.; Lahcen, O.; Abdelaziz, A. Breaking seed dormancy in Cupressus atlantica Gaussen, an endemic and threatened coniferous tree in Morocco. J. For. Res. 2012, 23, 385–390. [Google Scholar] [CrossRef]
- Fabricante, J.R.; de Castro, R.A.; de Araujo, K.C.T.; de Siqueira-Filho, J.A. Ecological Attributes of Alien Nicotiana glauca Graham (Solanacea) And Assessment of The Ssusceptibility of The Species Ooccurring in Brazil. Cienc. Florest. 2015, 25, 959–967. [Google Scholar] [CrossRef]
- Henschel, J.R.; Parr, T. Population changes of alien invasive plants in the Lower Kuiseb River. Dinteria 2010, 31, 5–17. [Google Scholar]
- Florentine, S.K.; Weller, S.; Graz, P.F.; Westbrooke, M.; Florentine, A.; Javaid, M.; Fernando, N.; Chauhan, B.S.; Dowling, K. Influence of selected environmental factors on seed germination and seedling survival of the arid zone invasive species tobacco bush (Nicotiana glauca R. Graham). Rangel. J. 2016, 38, 417–425. [Google Scholar] [CrossRef]
- DiTomaso, J.M.; Kyser, G.B.; Oneto, S.R.; Wilson, R.G.; Orloff, S.B.; Anderson, L.W.; Wright, S.D.; Roncoroni, J.A.; Miller, T.L.; Prather, T.S. Weed control in natural areas in the western United States. Weed Res. Inf. Center Univ. Calif. 2013, 544. [Google Scholar]
- Nieto, A.C.; Pinós, M.C.C.; Cabot, P.; Pitet, M.; Estaún, V. Arbuscular mycorrhizal fungi associated with psammophilic vegetation in Mediterranean coastal sand dunes. Spanish J. Agric. Res. 2010, 8, 96–102. [Google Scholar]
- El Mrabet, S.; Ouahmane, L.; El Mousadik, A.; Msanda, F.; Abbas, Y. The Effectiveness of Arbuscular Mycorrhizal Inoculation and Bio-Compost Addition for Enhancing Reforestation with Argania spinosa in Morocco. Open J. For. 2014, 4, 14–23. [Google Scholar]
- Ouahmane, L.; Duponnois, R.; Hafidi, M.; Kisa, M.; Boumezouch, A.; Thioulouse, J.; Plenchette, C. Some Mediterranean plant species (Lavandula spp. and Thymus satureioides) act as potential ‘plant nurses’ for the early growth of Cupressus atlantica. Plant Ecol. 2006, 185, 123–134. [Google Scholar] [CrossRef]
- Elmostapha, O.; Hanane, D.; Rachid, B.; Lahcen, O. Application of Arbuscular Mycorrhizal Fungi Isolates from Semi-arid Mediterranean Ecosystems as Biofertilizers in Argan Tree Development. J. Soil Sci. Plant Nutr. 2021, 22, 944–955. [Google Scholar] [CrossRef]
- Azcón-Aguilar, C.; Palenzuela, J.; Roldán, A.; Bautista, S.; Vallejo, R.; Barea, J.M. Analysis of the mycorrhizal potential in the rhizosphere of representative plant species from desertification-threatened Mediterranean shrublands. Appl. Soil Ecol. 2003, 22, 29–37. [Google Scholar] [CrossRef]
- Lahcen, O.; Ibrahima, N.; Abdessadek, M.; Abderrahim, F.; Youssef, S.; Mohamed, A. Inoculation of Ceratonia siliqua L. with native arbuscular mycorrhizal fungi mixture improves seedling establishment under greenhouse conditions. Afr. J. Biotechnol. 2012, 11, 16422–16426. [Google Scholar]
- Diop, T.A.; Gueye, M.; Dreyfus, B.L.; Plenchette, C.; Strullu, D.G. Indigenous arbuscular mycorrhizal fungi associated with Acacia albida Del. in different areas of Senegal. Appl. Environ. Microbiol. 1994, 60, 3433–3436. [Google Scholar] [CrossRef]
- An, Z.-Q.; Hendrix, J.W.; Hershman, D.E.; Henson, G.T. Evaluation of the “most probable number”(MPN) and wet-sieving methods for determining soil-borne populations of endogonaceous mycorrhizal fungi. Mycologia 1990, 82, 576–581. [Google Scholar] [CrossRef]
- Bouazza, M.K.; Ighilharizn, Z.; de Lajudie, P.; Duponnois, R.; Bekki, A. Assessing th native arbuscular mycorrhizal symbioses to rehabilitate a degraded coastal sand dune in Algeria. Int. J. Agric. Crop. Sci. 2015, 8, 194–202. [Google Scholar]
- Maier, W.; Schmidt, J.; Nimtz, M.; Wray, V.; Strack, D. Secondary products in mycorrhizal roots of tobacco and tomato. Phytochemistry 2000, 54, 473–479. [Google Scholar] [CrossRef]
- Reynolds, H.L.; Packer, A.; Bever, J.D.; Clay, K. Grassroots ecology: Plant–microbe–soil interactions as drivers of plant community structure and dynamics. Ecology 2003, 84, 2281–2291. [Google Scholar] [CrossRef]
- Barazani, O.; Sathiyamoorthy, P.; Manandhar, U.; Vulkan, R.; Golan-Goldhirsh, A. Heavy metal accumulation by Nicotiana glauca Graham in a solid waste disposal site. Chemosphere 2004, 54, 867–872. [Google Scholar] [CrossRef]
- Shingu, Y.; Kudo, T.; Ohsato, S.; Kimura, M.; Ono, Y.; Yamaguchi, I.; Hamamoto, H. Characterization of genes encoding metal tolerance proteins isolated from Nicotiana glauca and Nicotiana tabacum. Biochem. Biophys. Res. Commun. 2005, 331, 675–680. [Google Scholar] [CrossRef]
- Gisbert, C.; Ros, R.; De Haro, A.; Walker, D.J.; Bernal, M.P.; Serrano, R.; Navarro-Aviñó, J. A plant genetically modified that accumulates Pb is especially promising for phytoremediation. Biochem. Biophys. Res. Commun. 2003, 303, 440–445. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).