
Influence of Growing Substrate Preparation on the Biological Efficiency of Pleurotus ostreatus

 ,
, 
 and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Substrate Preparation and Experimental Design
2.2. Chemical Analyses
2.3. Biochemical Analyses
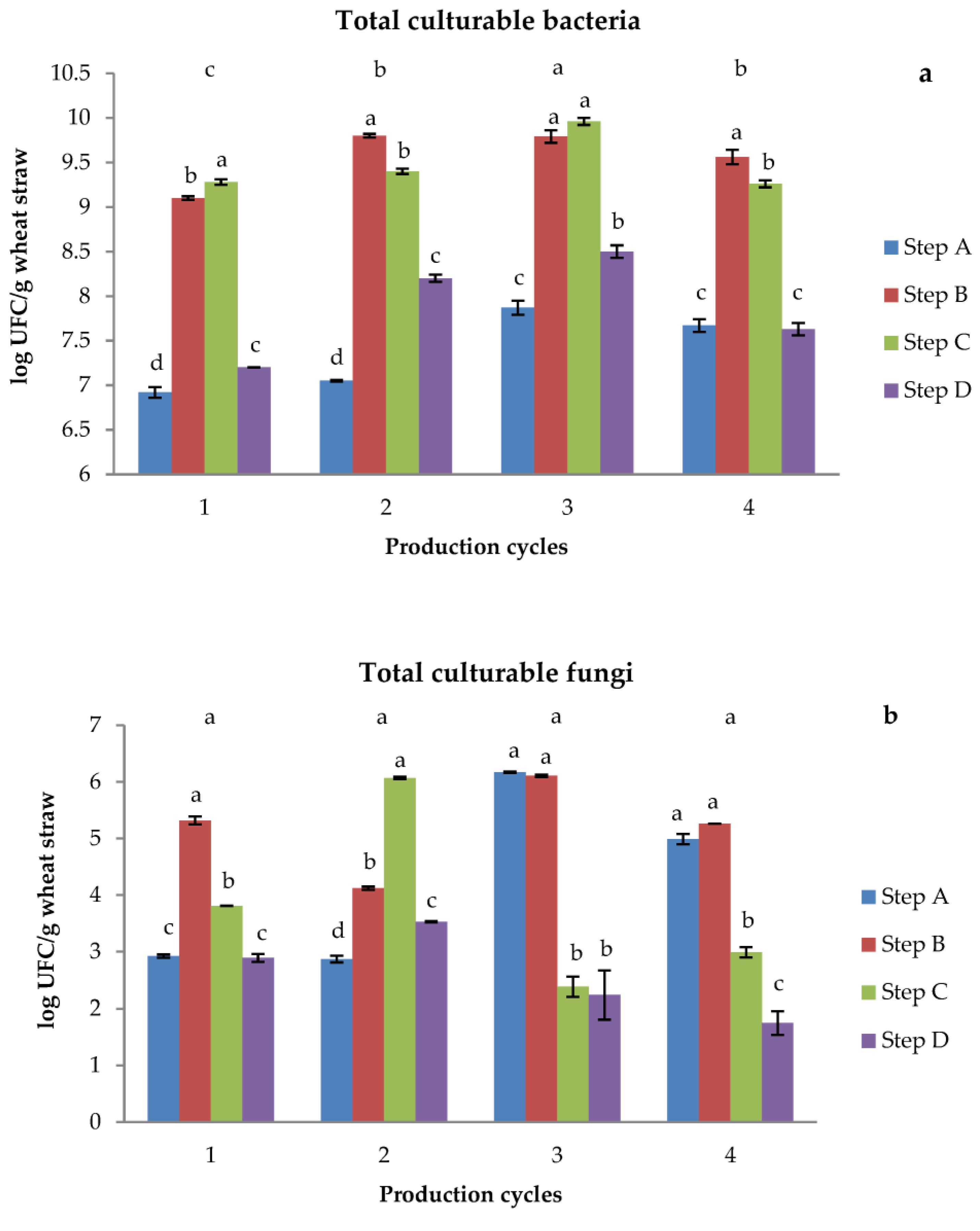
2.3.1. Bacterial and Fungal Counts
2.3.2. Enzymatic and Microbial Activities
2.3.3. DNA Extraction
2.3.4. PCR, Electrophoretic Profiles, and Cluster Analysis
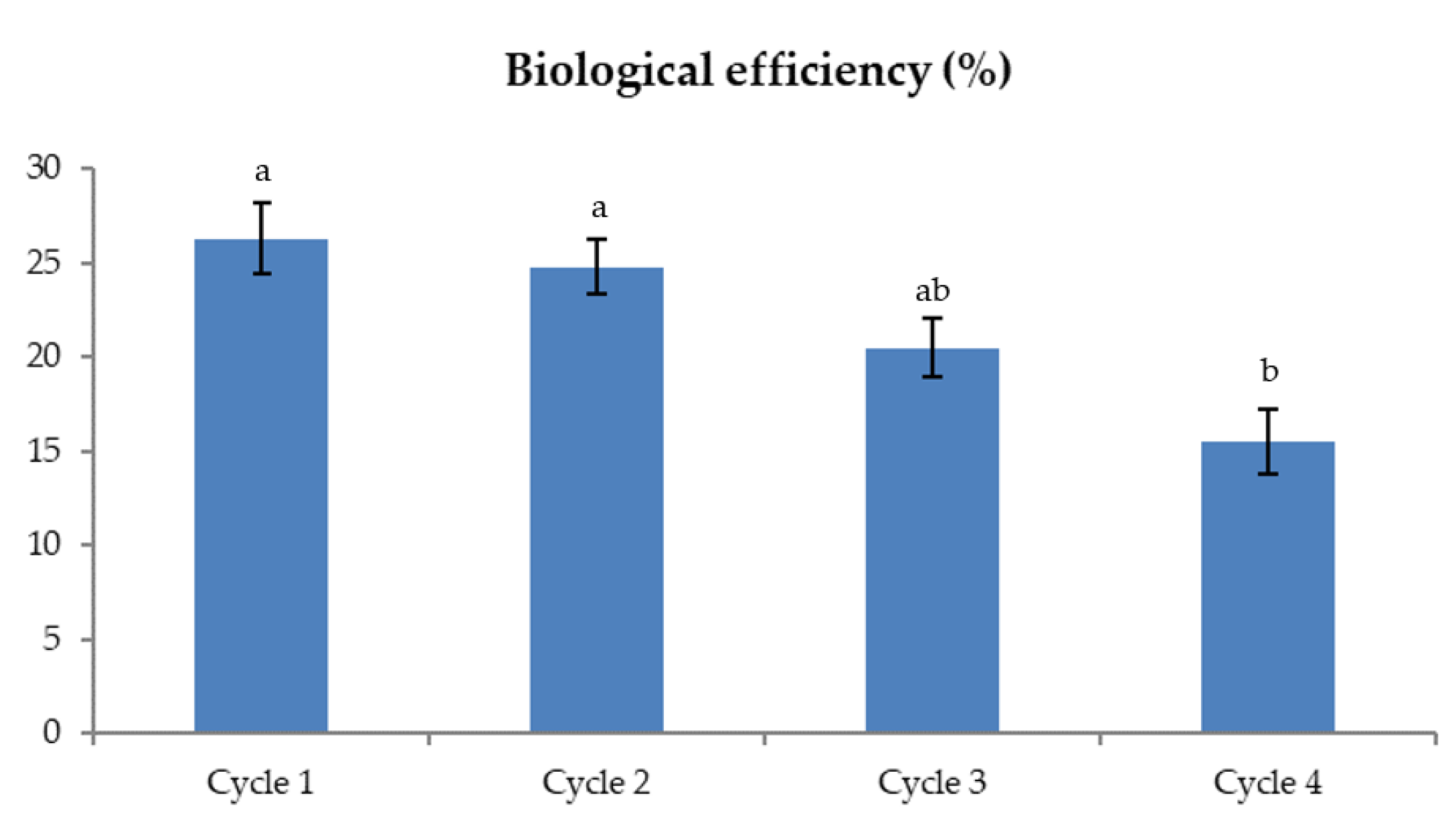
2.4. Biological Efficiency/Mushroom Yields
2.5. Statistical Analysis
3. Results and Discussion
3.1. Chemical Analyses of Raw Straw and Growing Media
3.2. Microbiological Analyses of Raw Straw and Growing Media
3.3. Enzymatic Analyses of Raw Straw and Growing Media
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Melanouri, E.M.; Dedousi, M.; Diamantopoulou, P. Cultivating Pleurotus ostreatus and Pleurotus eryngii mushroom strains on agro-industrial residues in solid-state fermentation. Part II: Effect on productivity and quality of carposomes. Carbon Resour. Convers. 2022, 5, 52–60. [Google Scholar] [CrossRef]
- Muswati, C.; Simango, K.; Tapfumaneyi, L.; Mutetwa, M.; Ngezimana, W. The Effects of Different Substrate Combinations on Growth and Yield of Oyster Mushroom (Pleurotus ostreatus). Int. J. Agron. 2021, 2021, 9962285. [Google Scholar] [CrossRef]
- Abou Fayssal, S.; Alsanad, M.A.; Yordanova, M.H.; El Sebaaly, Z.; Najjar, R.; Sassine, Y.N. Effect of olive pruning residues on substrate temperature and production of oyster mushroom (Pleurotus ostreatus). Acta Hortic. 2021, 1327, 245–252. [Google Scholar] [CrossRef]
- Wang, D.; Sakoda, A.; Suziki, M. Biological efficiency and nutritional value of Pleurotus ostreatus cultivated on spent beer grain. Bioresour. Technol. 2001, 78, 293–300. [Google Scholar] [CrossRef]
- Bellettini, M.B.; Fiorda, F.A.; Maieves, H.A.; Teixeira, G.L.; Ávila, S.; Hornung, P.S.; Maccari Júnior, A.; Hoffmann Ribani, R. Factors affecting mushroom Pleurotus spp. Saudi J. Biol. Sci. 2019, 26, 633–646. [Google Scholar] [CrossRef]
- Yang, W.; Guo, F.; Wan, Z. Yield and size of oyster mushroom grown on rice/wheat straw basal substrate supplemented with cotton seed hull. Saudi J. Biol. Sci. 2013, 20, 333–338. [Google Scholar] [CrossRef]
- Getachew, A.; Negassa, A.K.; Aga, M.C. Production of Oyster Mushroom (Pleurotus ostreatus) on Substrate Composed from Wheat Straw, Waste Paper and Cotton Seed Waste. Int. J. Microbiol. Biotechnol. 2019, 4, 38–44. [Google Scholar] [CrossRef]
- Economou, C.N.; Diamantopoulou, P.A.; Philippoussis, A.N. Valorization of spent oyster mushroom substrate and laccase recovery through successive solid state cultivation of Pleurotus, Ganoderma, and Lentinula strains. Appl. Microbiol. Biotechnol. 2017, 101, 5213–5222. [Google Scholar] [CrossRef]
- Zied, D.C.; Prado, E.P.; Dias, E.S.; Pardo, J.E.; Pardo-Gimenez, A. Use of peanut waste for oyster mushroom substrate supplementation-oyster mushroom and peanut waste. Braz. J. Microbiol. 2019, 50, 1021–1029. [Google Scholar] [CrossRef]
- Sadh, P.K.; Duhan, S.; Duhan, J.S. Agro-industrial wastes and their utilization using solid state fermentation: A review. Bioresour. Bioprocess. 2018, 5, 1. [Google Scholar] [CrossRef]
- Kumar, P.; Kumar, V.; Eid, E.M.; AL-Huqail, A.A.; Adelodun, B.; Abou Fayssal, S.; Goala, M.; Arya, A.K.; Bachheti, A.; Andabaka, Ž.; et al. Spatial Assessment of Potentially Toxic Elements (PTE) Concentration in Agaricus bisporus Mushroom Collected from Local Vegetable Markets of Uttarakhand State, India. J. Fungi 2022, 8, 452. [Google Scholar] [CrossRef]
- Yang, Y.R.; Guo, Y.X.; Wang, Q.Y.; Hu, B.Y.; Tian, S.Y.; Yang, Q.Z.; Cheng, Z.A.; Chen, Q.J.; Zhang, G.Q. Impacts of composting duration on physicochemical properties and microbial communities during short-term composting for the substrate for oyster mushrooms. Sci. Total Environ. 2022, 847, 157673. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.X.; Chen, Q.J.; Qin, Y.; Yang, Y.R.; Yang, Q.Z.; Wang, Y.X.; Cheng, Z.A.; Cao, N.; Zhang, G.Q. Succession of the microbial communities and function prediction during short-term peach sawdust-based composting. Bioresour. Technol. 2021, 332, 125079. [Google Scholar] [CrossRef] [PubMed]
- Kong, W.; Sun, B.; Zhang, J.; Zhang, Y.; Gu, L.; Bao, L.; Liu, S. Metagenomic analysis revealed the succession of microbiota and metabolic function in corncob composting for preparation of cultivation medium for pleurotus ostreatus. Bioresour. Technol. 2020, 306, 123156. [Google Scholar] [CrossRef] [PubMed]
- Joshi, A.K.; Kumar, U.; Mishra, V.K.; Chand, R.; Chatrath, R.; Naik, R.; Biradar, S.; Singh, R.P.; Budhlakoti, N.; Devulapalli, R.; et al. Variations in straw fodder quality and grain-Straw relationships in a mapping population of 287 diverse spring wheat lines. Field Crops Res. 2019, 243, 107627. [Google Scholar] [CrossRef] [PubMed]
- Roth, M.G.; Mourtzinis, S.; Gaska, J.M.; Mueller, B.; Roth, A.; Smith, D.L.; Conley, S.P. Wheat grain and straw yield, grain quality, and disease benefits associated with increased management intensity. Agron. J. 2021, 113, 308–320. [Google Scholar] [CrossRef]
- Fanadzo, M.; Zireva, D.T.; Dube, E.; Mashingaidze, A.B. Evaluation of various substrates and supplements for biological efficiency of Pleurotus sajor-caju and Pleurotus ostreatus. Afr. J. Biotechnol. 2010, 9, 2756–2761. [Google Scholar] [CrossRef]
- Trinchera, L.; Leita, P.; Sequi, P. Metodi di Analisi per i Fertilizzanti; Ministero delle Politiche Agricole Alimentari e Forestali: Rome, Italy, 2006. [Google Scholar]
- Ciavatta, C.; Antisari, L.V.; Sequi, P. Determination of organic carbon in soils and fertilizers. Commun. Soil Sci. Plant Anal. 1989, 20, 759–773. [Google Scholar] [CrossRef]
- Englyst, H.; Cummings, J.H. Simplified method for the measurement of total non-starch polysaccharides by gas-liquid chromatography of constituent sugars as alditol acetates. Analyst 1984, 109, 937–942. [Google Scholar] [CrossRef]
- Englyst, H.; Wiggins, H.S.; Cummings, J.H. Determination of the non-starch polysaccharides in plant foods by gas-liquid chromatography of constituent sugars as alditol acetates. Analyst 1982, 107, 307–318. [Google Scholar] [CrossRef]
- De Mastro, F.; Traversa, A.; Brunetti, G.; Debiase, G.; Cocozza, C.; Nigro, F. Soil culturable microorganisms as affected by different soil managements in a two year wheat-faba bean rotation. Appl. Soil Ecol. 2020, 149, 103533. [Google Scholar] [CrossRef]
- Yu, X.; Liu, Y.; Cui, Y.; Cheng, Q.; Zhang, Z.; Lu, J.H.; Meng, Q.; Teng, L.; Ren, X. Measurement of filter paper activities of cellulase with microplate-based assay. Saudi J. Biol. Sci. 2016, 23, S93–S98. [Google Scholar] [CrossRef] [PubMed]
- Mangalassery, S.; Mooney, S.; Sparkes, D.; Fraser, W.; Sjögersten, S. Impacts of zero tillage on soil enzyme activities, microbial characteristics and organic matter functional chemistry in temperate soils. Eur. J. Soil Biol. 2015, 68, 9–17. [Google Scholar] [CrossRef]
- Li, Q.; Coffman, A.M.; Ju, L.-K. Development of reproducible assays for polygalacturonase and pectinase. Enzym. Microb. Technol. 2015, 72, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Zervakis, G.I.; Koutrotsios, G.; Katsaris, P. Composted versus raw olive mill waste as substrates for the production of medicinal mushrooms: An assessment of selected cultivation and quality parameters. BioMed. Res. Int. 2013, 2013, 546830. [Google Scholar] [CrossRef] [PubMed]
- Cocozza, C.; Parente, A.; Zaccone, C.; Mininni, C.; Santamaria, P.; Miano, T. Comparative management of offshore posidonia residues: Composting vs. energy recovery. Waste Manag. 2011, 31, 78–84. [Google Scholar] [CrossRef]
- Miles, P.G.; Chang, S.T. Mushrooms: Cultivation, Nutritional Value, Medicinal Effect, and Environmental Impact, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2004; pp. 95–101. [Google Scholar]
- Kim, M.; Kim, W.S.; Tripathi, B.M.; Adams, J. Distinct bacterial communities dominate tropical and temperate zone leaf litter. Microbial. Ecol. 2014, 67, 837–848. [Google Scholar] [CrossRef]
- Mussatto, S.I.; Teixeira, J.A. Lignocellulose as Raw Material in Fermentation Processes. Curr. Res. Top. Appl. Microbiol. Microb. Biotechnol. 2010, 2, 897–907. [Google Scholar]
- Ma, N.L.; Khoo, S.C.; Peng, W.; Ng, C.M.; Teh, C.H.; Park, Y.-K.; Lam, S.S. Green application and toxic risk of used diaper and food waste as growth substitute for sustainable cultivation of oyster mushroom (Pleurotus ostreatus). J. Clean. Prod. 2020, 268, 122272. [Google Scholar] [CrossRef]
- Zhang, H.-L.; Wei, J.-K.; Wang, Q.-H.; Yang, R.; Gao, X.-J.; Sang, Y.-X.; Cai, P.-P.; Zhang, G.-Q.; Chen, Q.-J. Lignocellulose utilization and bacterial communities of millet straw based mushroom (Agaricus bisporus) production. Sci. Rep. 2019, 9, 1151. [Google Scholar] [CrossRef]
- Peksen, A.; Yakupoglu, G. Tea waste as a supplement for the cultivation of Ganoderma lucidum. World J. Microbiol. Biotechnol. 2009, 25, 611–618. [Google Scholar] [CrossRef]
- Liang, Z.; Wu, C.; Shieh, Z.; Cheng, S. Utilisation of grass plants for cultivation of Pleurotus citrinopeleatus. Int. Biodeterior. Biodegrad. 2009, 63, 509–514. [Google Scholar] [CrossRef]
- Frimpong–Manso, J.; Obodai, M.; Dzomeku, M.; Apertorgbor, M.M. Influence of rice husk on biological efficiency and nutrient content of Pleurotus ostreatus (Jacq. ex. Fr.) Kummer. Int. Food Res. J. 2011, 18, 249–254. [Google Scholar]
- Wang, X.; He, P.; Xu, X.; Qiu, S.; Zhao, S. Characteristics of rice straw decomposition and bacterial community succession for 2 consecutive years in a paddy field in southeastern China. Sci. Rep. 2022, 12, 20893. [Google Scholar] [CrossRef]
- Fan, F.L.; Yin, C.; Tang, Y.; Li, Z.; Song, A.; Wakelin, S.A.; Zou, J.; Liang, Y. Probing potential microbial coupling of carbon and nitrogen cycling during decomposition of maize residue by 13C-DNA-SIP. Soil Biol. Biochem. 2014, 70, 12–21. [Google Scholar] [CrossRef]
- Paterson, E.; Osler, G.; Dawson, L.A.; Gebbing, T.; Sim, A.; Ord, B. Labile and recalcitrant plant fractions are utilized by distinct microbial communities in soil: Independent of the presence of roots and mycorrhizal fungi. Soil Biol. Biochem. 2008, 40, 1103–1113. [Google Scholar] [CrossRef]
- Marschner, P.; Umar, S.; Baumann, K. The microbial community composition changes rapidly in the early stages of decomposition of wheat residue. Soil Biol. Biochem. 2011, 43, 445–451. [Google Scholar] [CrossRef]
- Makhatov, Z.; Alibayev, N.; Konarbayeva, Z.; Makhatov, B.; Makhatova, A.; Doltayeva, B.; Torlanova, B.; Arystanbaev, K.; Shagrayeva, B. Enzymatic depolymerization of wheat straw polysaccharides. In Proceedings of the IOP Conference Series: Earth and Environmental Science, 2nd International Conference on Energetics, Civil and Agricultural Engineering 2021 (ICECAE 2021), Tashkent, Uzbekistan, 14–16 October 2021; Volume 939, p. 012005. [Google Scholar] [CrossRef]
- Iqbal, H.M.N.; Ahmed, I.; Zia, M.A.; Irfan, M. Purification and characterization of the kinetic parameters of cellulose produced from wheat straw by Trichoderma viride under SSF and its detergent compatibility. Adv. Biosci. Biotechnol. 2011, 2, 149–156. [Google Scholar] [CrossRef]
- De Mastro, F.; Brunetti, G.; Traversa, A.; Blagodatskaya, E. Fertilization promotes microbial growth and minimum tillage increases nutrient-acquiring enzyme activities in a semiarid agro-ecosystem. Appl. Soil Ecol. 2022, 177, 104529. [Google Scholar] [CrossRef]
- Chiliveri, S.R.; Koti, S.; Linga, V.R. Retting and degumming of natural fibers by pectinolytic enzymes produced from Bacillus tequilensis SV11-UV37 using solid state fermentation. SpringerPlus 2016, 5, 559. [Google Scholar] [CrossRef]
- Ćilerdžić, J.; Galić, M.; Stajić, M. From pomiculture waste to biotechnological raw material: Efficient transformation using ligninosomes and cellulosomes from Pleurotus spp. Bioresour. Bioprocess. 2022, 9, 66. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Preparation Steps | Material Sampled |
|---|---|
| A | Raw straw |
| B | Wet straw, chopped and augmented with soy flour |
| C | Substrate at the end of 5 days of composting |
| D | Substrate after steam pasteurization |
| Mushroom Production Cycles | Starting Time | Straw Age |
|---|---|---|
| Cycle 1 | July 2019 | Fresh |
| Cycle 2 | October 2019 | 3 months after harvest |
| Cycle 3 | January 2020 | 6 months after harvest |
| Cycle 4 | April 2020 | 9 months after harvest |
| Moisture | Ashes | pH | OC | TN | C/N | |
|---|---|---|---|---|---|---|
| % | g·kg −1 | |||||
| Cycle | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. |
| Step | *** | *** | *** | *** | * | n.s. |
| Cycle | ||||||
| 1 | 57.6 a (3.1) | 11.2 a (3.2) | 8.1 a (0.9) | 507 a (1.8) | 6.8 a (0.9) | 74.9 a (10.4) |
| 2 | 58.2 a (3.1) | 14.9 a (6.6) | 7.5 a (0.5) | 486 a (3.8) | 6.4 a (2.1) | 85.2 a (12.9) |
| 3 | 55.3 a (2.7) | 13.2 a (4.6) | 7.9 a (0.5) | 495 a (2.6) | 7.2 a (1.4) | 71.6 a (17.8) |
| 4 | 59.0 a (3.2) | 13.9 a (6.1) | 8.0 a (0.6) | 489 a (3.1) | 7.8 a (1.8) | 66.8 a (14.2) |
| Step | ||||||
| A | 11.8 c (1.3) | 7.4 b (0.7) | 7.0 c (0.2) | 526 a (0.4) | 5.1 b (1.1) | 107.5 a (18.0) |
| B | 67.5 b (1.6) | 10.7 b (0.4) | 7.8 b (0.1) | 510 a (0.2) | 7.7 ab (1.5) | 67.8 a (13.7) |
| C | 74.5 ab (3.1) | 16.9 a (3.6) | 8.4 ab (0.5) | 467 b (2.0) | 7.5 ab (1.1) | 63.2 a (11.8) |
| D | 76.3 a (5.8) | 18.2 a (2.8) | 8.4 a (0.4) | 474 b (1.3) | 7.9 a (0.7) | 60.0 a (4.2) |
| Total Sugars | Soluble Sugars | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ARA | GAL | GLU | XYL | TOT | ARA | GAL | GLU | XYL | FRU | SUC | TOT | |
| % dry weight | mg·kg−1 | |||||||||||
| Cycle | *** | * | *** | ** | ** | n.s. | n.s. | *** | n.s. | * | *** | ** |
| Step | n.s. | * | * | * | * | * | * | ** | n.s. | * | *** | *** |
| Cycle | ||||||||||||
| 1 | 4.4 a (0.2) | 1.2 a (0.2) | 50.2 a (1.1) | 24.5 a (0.9) | 80.3 a (1.7) | 0.8 a (0.1) | 0.6 a (0.1) | 76.0 a (6.2) | 0.3 a (0.1) | 7.1 a (1.3) | 123.0 a (20.4) | 207.8 a (50.2) |
| 2 | 2.6 b (0.4) | 0.8 b (0.1) | 47.2 ab (2.3) | 19.7 b (2.1) | 70.2 ab (4.3) | 1.0 a (0.2) | 0.2 a (0.1) | 24.2 b (2.7) | 0.0 a (0.0) | 7.6 a (1.0) | 67.1 b (8.6) | 100.2 b (18.3) |
| 3 | 2.4 b (0.5) | 0.8 b (0.1) | 42.0 c (2.7) | 20.5 b (1.2) | 65.6 b (4.4) | 0.6 a (0.1) | 0.4 a (0.1) | 5.7 c (1.1) | 0.0 a (0.0) | 1.4 b (0.8) | 7.3 c (2.8) | 15.4 c (7.2) |
| 4 | 2.8 b (0.5) | 1.0 ab (0.3) | 46.0 b (1.6) | 20.8 b (2.7) | 70.6 ab (6.1) | 0.7 a (0.3) | 0.3 a (0.1) | 5.0 c (0.9) | 0.2 a (0.0) | 2.4 b (0.7) | 10.0 c (3.4) | 18.5 c (4.2) |
| Step | ||||||||||||
| A | 3.2 a (0.9) | 1.0 ab (0.2) | 48.0 a (3.2) | 22.9 a (2.0) | 75.1 a (2.9) | 1.6 a (0.7) | 0.9 a (0.2) | 85.7 a (8.3) | 0.0 a (0.0) | 8.7 a (3.2) | 167.2 a (20.3) | 264.1 a (31.2) |
| B | 3.3 a (1.0) | 1.2 a (0.1) | 47.2 a (2.6) | 22.4 a (1.2) | 74.0 a (1.5) | 0.5 b (0.1) | 0.3 b (0.2) | 14.0 b (3.4) | 0.0 a (0.0) | 5.4 ab (2.4) | 25.9 b (6.4) | 46.1 b (12.6) |
| C | 3.1 a (0.7) | 0.9 b (0.1) | 47.0 a (3.2) | 20.6 ab (2.7) | 71.5 ab (3.1) | 0.5 b (0.1) | 0.2 b (0.1) | 7.4 bc (1.9) | 0.1 a (0.0) | 2.8 b (1.2) | 9.0 c (2.8) | 20 bc (4.2) |
| D | 2.6 a (1.0) | 0.8 b (0.2) | 43.2 b (1.2) | 19.6 b (1.1) | 66.1 b (2.2) | 0.5 b (0.1) | 0.2 b (0.2) | 3.9 c (0.8) | 0.4 a (0.1) | 1.6 b (0.4) | 5.2 c (1.2) | 11.8 c (3.2) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Mastro, F.; Traversa, A.; Matarrese, F.; Cocozza, C.; Brunetti, G. Influence of Growing Substrate Preparation on the Biological Efficiency of Pleurotus ostreatus. Horticulturae 2023, 9, 439. https://doi.org/10.3390/horticulturae9040439
De Mastro F, Traversa A, Matarrese F, Cocozza C, Brunetti G. Influence of Growing Substrate Preparation on the Biological Efficiency of Pleurotus ostreatus. Horticulturae. 2023; 9(4):439. https://doi.org/10.3390/horticulturae9040439
Chicago/Turabian StyleDe Mastro, Francesco, Andreina Traversa, Francesco Matarrese, Claudio Cocozza, and Gennaro Brunetti. 2023. "Influence of Growing Substrate Preparation on the Biological Efficiency of Pleurotus ostreatus" Horticulturae 9, no. 4: 439. https://doi.org/10.3390/horticulturae9040439
APA StyleDe Mastro, F., Traversa, A., Matarrese, F., Cocozza, C., & Brunetti, G. (2023). Influence of Growing Substrate Preparation on the Biological Efficiency of Pleurotus ostreatus. Horticulturae, 9(4), 439. https://doi.org/10.3390/horticulturae9040439