4.1. Molecular Variability
Identification of genetic diversity patterns is of significant importance for effective management and species conservation especially of endemic and threatened taxa [
41,
42,
43,
44]. There are no larger differences in the level of gene diversity, private alleles, and DW among the
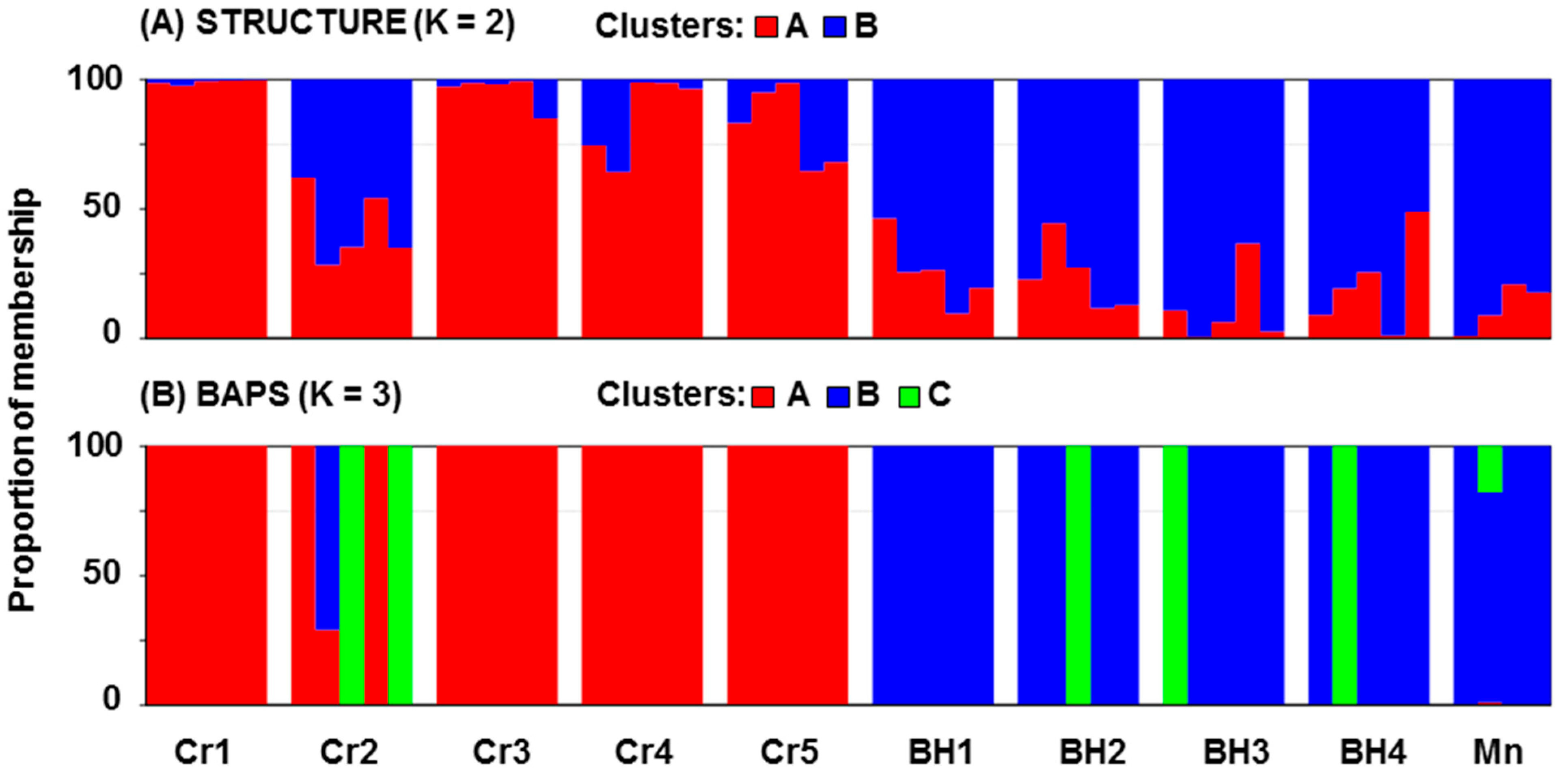
Micromeria croatica populations included in this study (
Table 2). The most diverse population is Cr2 which possesses the most private bands (15) but also contains individuals that belong to the eastern genetic group revealed by STRUCTURE (
Figure 3A). It is possible that it is caused by gene flow from some other nearby populations (not included in this study) which maybe belong to a third genetic group revealed by BAPS (
Figure 3B). Two spatially close western populations have relatively different DW values, which are 1262.15 (Cr2) and 887.47 (Cr3). Stronger genetic differentiation at short distances can indicate a much reduced power of seed dispersal and limited pollination over large distances. A similar result has been observed in some other endemic species [
45,
46,
47].
The results of STRUCTURE and BAPS analysis show the genetic differentiation of ten
M. croatica populations into two and three genetic groups, respectively. It is consistent with the known west–east separation in the range of this species and the presence of many isolated populations between and around these two groups [
5]. The STRUCTURE analysis reveals the existence of west–east split (
Figure 2 and
Figure 3A). This split is relative because some individuals from western populations were grouped into the eastern group and vice versa. Most of these ‘’transitional’’ individuals were grouped by BAPS in the third group (
Figure 3B). Such results suggest the presence of weak gene flow barriers among the western and eastern groups of
M. croatica populations. It is known that the differentiation of populations may have been fostered by natural habitat fragmentation. Hilpold et al. 2011 [
48] found that sandy beaches and river estuaries have probably acted as barriers for members of the Sicily group of
Centaurea cineraria L. group (family Asteraceae), which are restricted to rock faces close to the sea. The disjunction seen in
Centaurea aeolica Guss. ex Lojac. between Lipari and Ventotene islands (Italy) was best explained by long-distance (over about 250 km) for seed dispersal [
48]. According to Bittkau and Comes [
49], speciation within
Nigella L. s. lat. (Ranunculaceae) predominantly took place in allopatry. No significant effect on diversification rate was found regarding the establishment of a Mediterranean-type climate or the onset of the Quaternary climatic oscillations. The accelerated rate of speciation in the
Nigella arvensis L. complex is rather plausibly related to increased opportunities for allopatric speciation afforded by the (palaeo)geographical complexity of the Aegean archipelago combined with Late Pleistocene changes in climate and sea level [
49]. Our results suggested that 31.4% of the genetic differentiation between
M. croatica populations could be explained by geographical distances. Similarly, 26% of the genetic differentiation between the populations of
Helichrysum italicum (Roth) G. Don (Asteraceae) can be explained by their spatial distance [
50]. Even 66.61% of the genetic differentiation between
Degenia velebitica Hayek (Brassicaceae) populations was explained by geographical distance [
47]. On the other hand, only 5.6% of the genetic differentiation between
Tanacetum cinerariifolium (Trevir.) Sch. Bip. (Asteraceae) populations could be explained by geographical distance [
51]. It suggests that in addition to the factor of geographical isolation, other types of factors (height of growth above sea level, exposure, type and illumination of the slope, and biological features of the species) play a significant role in determining taxonomic diversity. The universal sets of adaptive markers include lipid composition [
52], which was previously shown [
53]. Hilpold et al. [
48] also mentioned the influence of the geological substratum and soil on the taxonomic diversity of
Centaurea aeolica. They found that
C. aeolica grows on volcanic soil, while the rocks along the coast south of Naples (Italy) are almost exclusively calcareous or granitic, rendering dispersal across the mainland difficult. According to Rešetnik et al. [
54],
Campanula fenestrellata Feer subsp.
istriaca (Feer) Damboldt (Campanulaceae) occupies narrower environmental space occurring in warmer and drier sites compared to
C. fenestrellata subsp.
fenestrellata which has a broader niche, tolerating a wider temperature range and generally occurring in colder and more humid sites. Local patterns of adaptation linked with altitude have been identified within
Arabidopsis halleri (L.) O’Kane et Al-Shehbaz,
A. thaliana (L.) Heynh., and
A. arenosa (L.) Lawalrée, suggests that altitude may be an important driver of genetic differentiation and adaptation in the entire genus [
55,
56,
57].
It is also possible that the partial isolation of
M. croatica populations by river valleys and mountain ranges led to genetic differentiation among populations over a long period of time. Although it is speculative to conclude this from such a limited data set, we can assume inter-population contacts happened periodically during the last glaciation cycles. Since
M. croatica is mountainous species, and bearing in mind habitats for such species became limited during the glaciation cycles [
58], they could only survive by altitudinal range shifts and descent to lower altitudes during the glaciation maximums [
41,
59]. In colder climate periods, especially during repeated glaciations in the Pleistocene, populations of this species probably occupied larger and less fragmented areas at lower altitudes. Once the climate got warmer again, they reclaimed their pre-glaciation habitats. Nonetheless, these migrations also served as opportunities for inter-population gene flow, as fragmented and geographically isolated populations came into direct contact. The fingerprints of these contacts can still be observed today as a reduced level of differentiation among seemingly isolated populations and the lack of clear genetic structure. The positive and significant correlations revealed by the IBD could be the result of successive postglacial colonization from the refugia [
60]. The two populations characterized by somewhat higher levels of DW markers likely experienced prolonged isolation that enabled them to accumulate more private alleles if compared to other ones. Phylogeographical patterns were analyzed in some plant and animal species from Balkan Peninsula. A deep phylogeographic split within
Edraianthus tenuifolius (Waldst. et Kit.) A. DC. (Campanulaceae), indicated by AFLP data, separates southeastern from northwestern populations. The noticed split coincides with the lowermost Neretva Valley (Croatia) [
61]. Kučera et al. [
62] found a phylogeographic and taxonomic split within the
Cardamine maritima Port. ex DC. agg. (Brassicaceae) in the Neretva Valley (Croatia, and Bosnia and Herzegovina), a genetic pattern on which they based, along with morphological data, the segregation of
Cardamine maritima and
C. fialae Fritsch. A similar split was found between the allopatric northwestern Adriatic group of
Campanula pyramidalis L. s. str. (Campanulaceae) and the southeastern Adriatic group of
C. austroadriatica D. Lakušić et Kovačić populations of the
C. pyramidalis complex along the lower Neretva Valley [
63]. The canyons in Dinaric karst were also shown to limit gene flow in
Tanacetum cinerariifolium (Trevir.) Sch. Bip. [
51] and
Campanula secundiflora Vis. et Panč. s. l. [
64]. The separation often coincides with isolated mountain ranges as also seen in some Western Balkan mountain animals [
65,
66]. Surina et al. [
61] also noticed that the lower Neretva River valley does not coincide with any major phylogeographic split indicating that the
Edraianthus serpyllifolius gene flow has occurred across this deep but narrow valley. In some populations of
E.
serpyllifolius gene exchange is also evident between mountain ranges [
61]. It is probable that gene exchange also exists between eastern populations of
M. croatica where four of five are located around the Neretva River Valley. Obtained results also show that mountain ranges are not strong enough barriers to prevent the gene exchange between nearby both western and eastern
M. croatica populations. Although weak, present gene flow barriers among populations maybe have led to further differentiation among populations. Investigations of some other Eastern Adriatic plant species also showed clustering of their populations in two or three genetic groups [
51,
61,
67,
68]. This coincides with Médail et Diadema’s (2009) [
69] view of the existence of a northern and more southern local glacial refugia along the Eastern Adriatic coast. Finally, Kremer et al. [
53] investigated
Micromeria and
Clinopodium species and concluded that no single refugial area can be identified within the Balkan Peninsula. Instead, there appear to have been numerous microrefugia scattered over large areas. Such a refugia-within-refugia model was first developed by Gómes and Lunt [
70] for the Iberian Peninsula.
4.2. Morphological Variability
Comparison of morphological with genetic traits showed that western populations have something longer, wider, and more orbicular leaves (
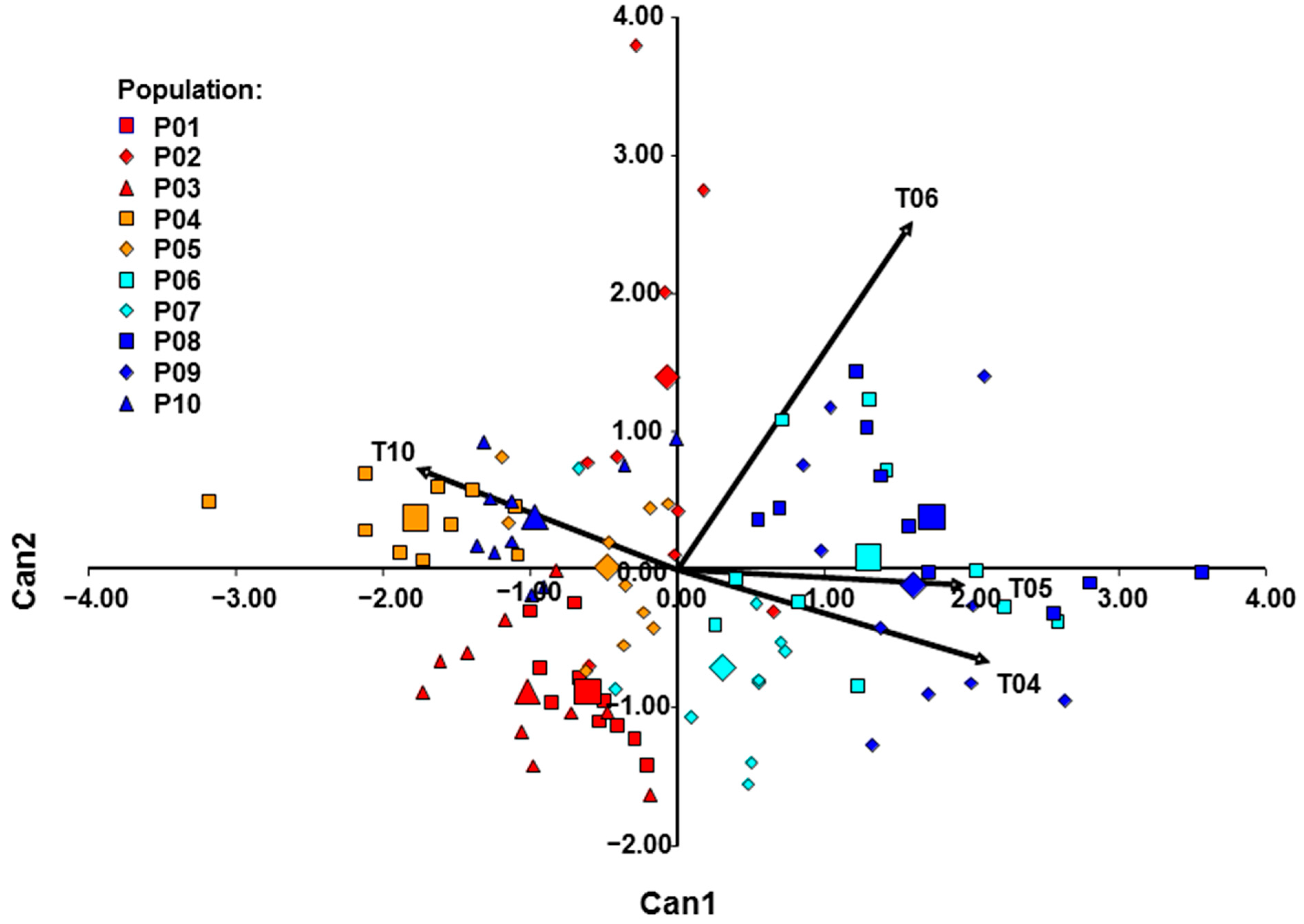
Table 5). The average number of flowers per shoot was 19.71 in the western and 40.98 in the eastern group. This indicates a more vigorous growth of the eastern group, which results in a higher number of flowers per shoot. The calyx length was approximately equal in both groups. On the other hand, calyx teeth were significantly longer in the eastern group. The discriminant function correctly classified 80.00% of plants into their respective clusters (
Figure 8). Most of the misclassified individuals belong to Mn, Cr2, and BH2 populations. It is evident that all plants from the Mn population were classified inside the western group. Similar to the results of STRUCTURE analysis (
Figure 3A), CANDISC analysis (
Figure 8) based on morphological traits showed that the Cr2 population is closer to the Bosnian population. This distinguishes the Cr2 population as a transitional population between the western and eastern groups of populations.
The genetic structuring of
M. croatica populations for horticultural purposes should be observed together with its morphological and ecological features. The introduction of new crops into cultivation includes many research stages that begin with the initial search and screening of germplasm [
71]. Initial screening of wild plants should include populations from climatologically different habitats. When studying different populations, one may well come across populations that have genetically adapted to survival in colder environments, to drought, or to other environmental constraints [
72].
M. croatica is a tiny plant adapted to the unfavorable water balance due to the consistent drying effect of strong wind and shallow, poorly developed soils that fill the crevices of carbonate rocks.
M. croatica stands are very distinct from the surrounding vegetation and limited to crevices and terraces of rocks, the foot of the rocks, and rocky slopes. It grows in open habitats with intense sun radiation and wide temperature fluctuations. Such stands are inhabited by a smaller number of plants and have low coverage of co-occurring taxa. Low above-ground competition in habitats of
M. croatica is in accordance with the ecological preferences of narrow endemics in the Mediterranean described by Lavergne et al. [
73]. It suggests the azonal occurrence of
M. croatica and restriction to nowadays spatially limited and fragmented microsites. Investigations of morphological traits showed that the most variable trait in
M. croatica was the number of flowers per shoot and the coefficient of variability ranged from 30.1% to 104.0% (
Table 5). Abundant flowering is one of the most common criteria for selecting plants for horticultural purposes. The great variability of this trait enables the potential selection of populations and individuals of
M. croatica with a larger number of flowers. With regard to the abundance of flowering, the populations BH1 with an average of 69.8, and BH3 with 75.9 flowers per shoot are the most promising populations. Considerable variability was also observed for the ratio between leaf length and leaf width as another feature of horticultural interest. This ratio gives insight into the leaf shape and the coefficient of variability for this trait ranged from 23.5% to 51% (
Table 5). A higher ratio between leaf length and width indicates more elongated, narrower leaves. On the other hand, a smaller ratio indicates more orbicular leaves. With regard to leaf shape, the Cr2 population with the most rotund leaves and the BH4 population with the narrowest leaves are the most promising. Finally, it should be mentioned that when we consider the cultivation of rare plants it should be kept in mind that commercial production of endemic and endangered species can prevent the collection from the wild and reduce the danger of a species becoming extinct [
74,
75].
Such as other
Micromeria and closely related
Clinopodium species which are grown as ornamental plants (
Table 1),
M. croatica can be planted in stony, dry soils in gardens. According to available bioclimatic data, precipitation of the driest month in investigated
M. croatica populations ranged from 53 to 89 mm, while the precipitation of the driest quarter ranged from 207 to 359 mm (
Table S1). Of course, there are some Mediterranean species that are more drought-tolerant than
M. croatica. Yet, unlike typical Mediterranean
Micromeria or
Clinopodium species, such as
Micromeria graeca or
M. juliana,
M. croatica is more resistant to low temperatures and can be cultivated in areas with a continental climate. The annual mean temperature in investigated
M. croatica populations ranged from 4.02 to 11.66 C, while the minimum temperature of the coldest month ranged from −3.40 to −7.00 C. Several plants of
M. croatica are cultivated from 2009 in Pharmaceutical Botanical Garden ‘’Fran Kušan’’ (Faculty of Pharmacy and Biochemistry, University of Zagreb, Croatia) in the continental part of Croatia. They bloom and fructify regularly, and young plants are ordinarily developing from the seeds. Planted plants do not require special attention in terms of watering and they were never fertilized from 2009. The only potential problem is that
M. croatica plants are sensitive to the overgrowth of weed species with which they are unable to compete for habitat. That is why the best place for their planting is garden rockeries. So, it can be said that the advantage of
M. croatica compared to Mediterranean
Micromeria species is that it not only tolerates drought but even better tolerates low temperatures. Under the framework of climate change, this makes it particularly suitable for planting in continental climate conditions.


 ,
, 












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}