



Growth and Nitrogen Uptake by Potato and Cassava Crops Can Be Improved by Azospirillum brasilense Inoculation and Nitrogen Fertilization

 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Site, Soil, Experimental Design, and Treatments
2.2. Planting and Management of Potato and Cassava
2.3. Sampling and Analyses
2.4. Statistical Analyses
3. Results
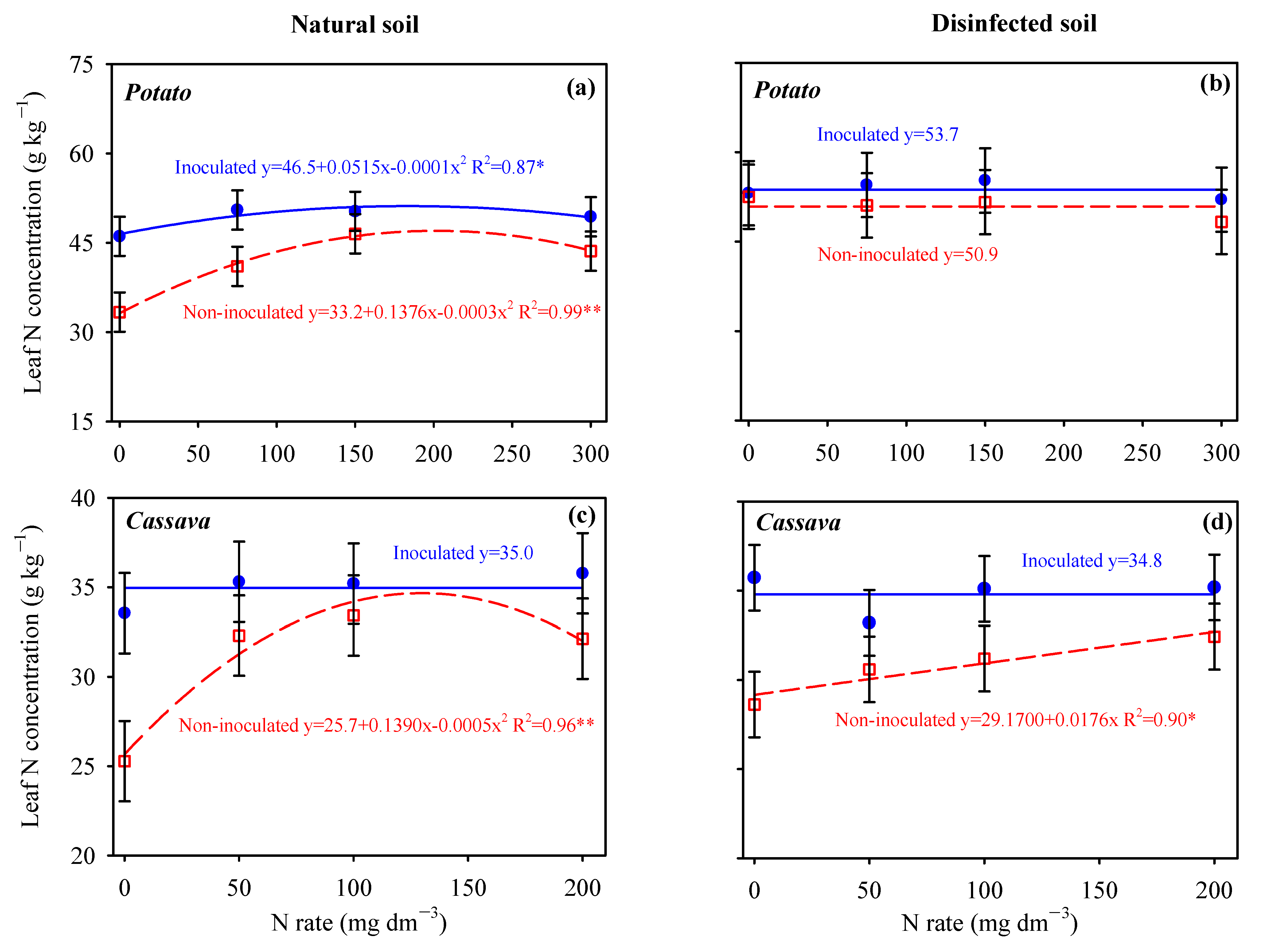
3.1. Nitrogen Status in the Leaves of Potato and Cassava
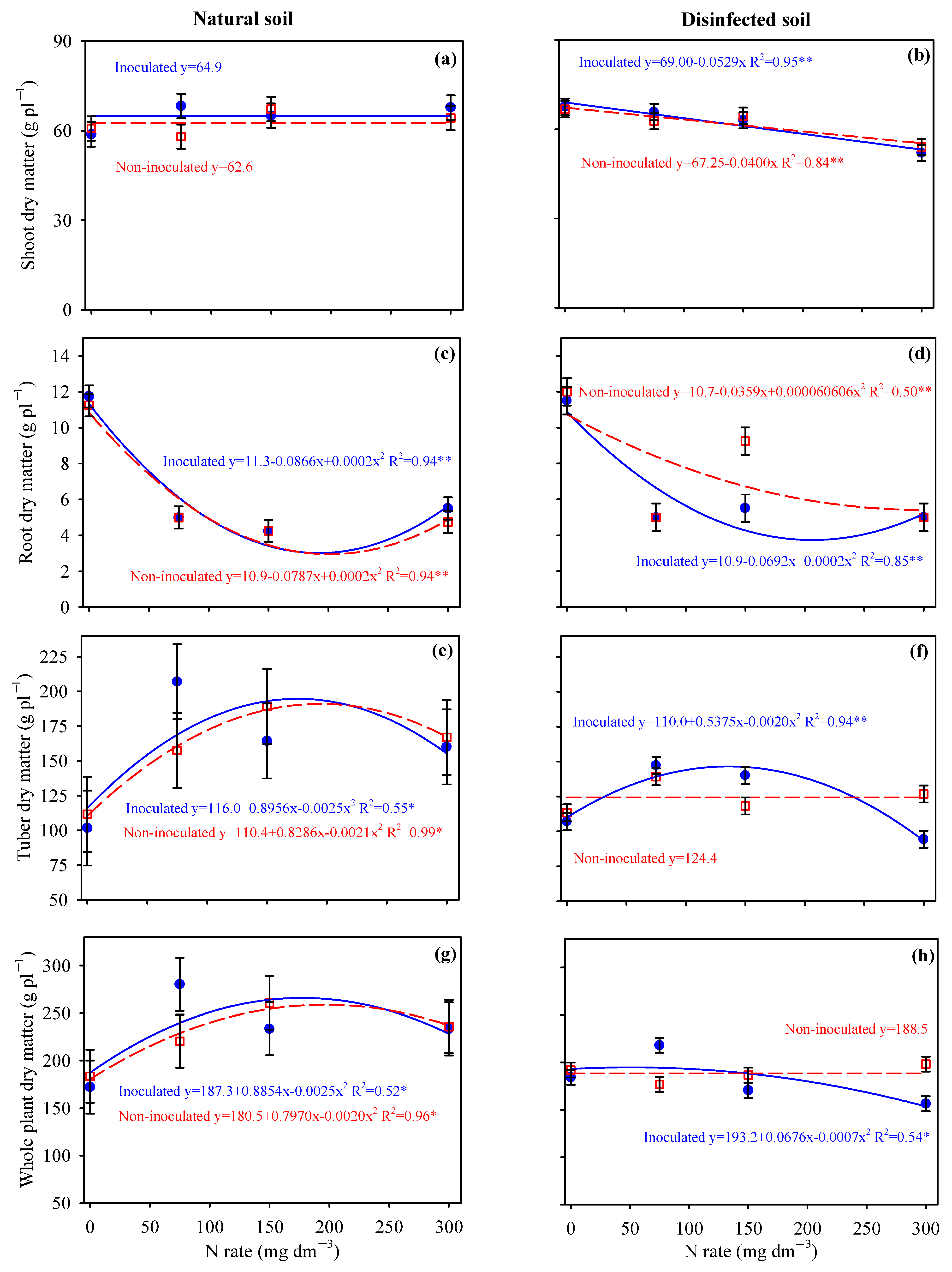
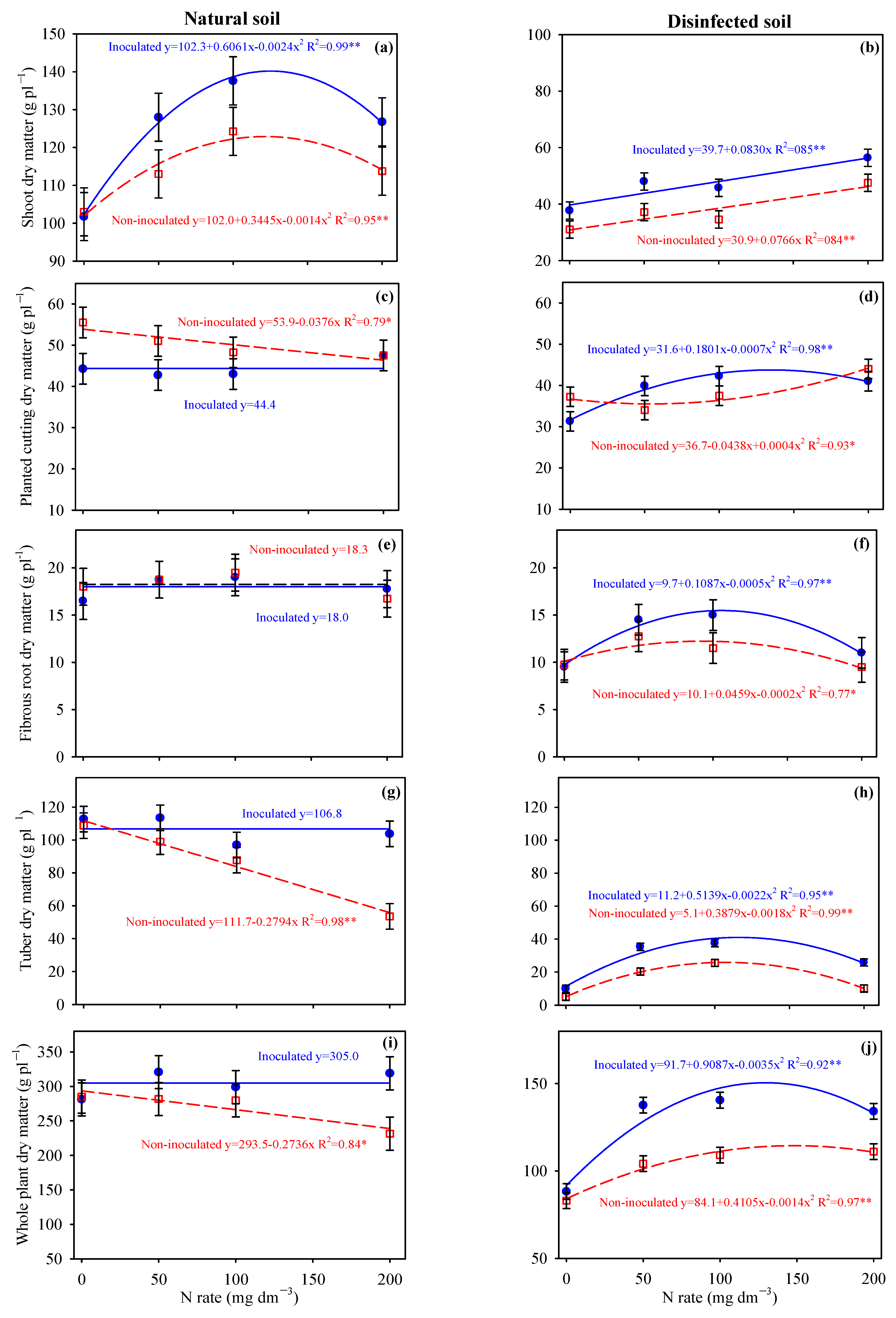
3.2. Biomass Accumulation in Potato and Cassava Plants
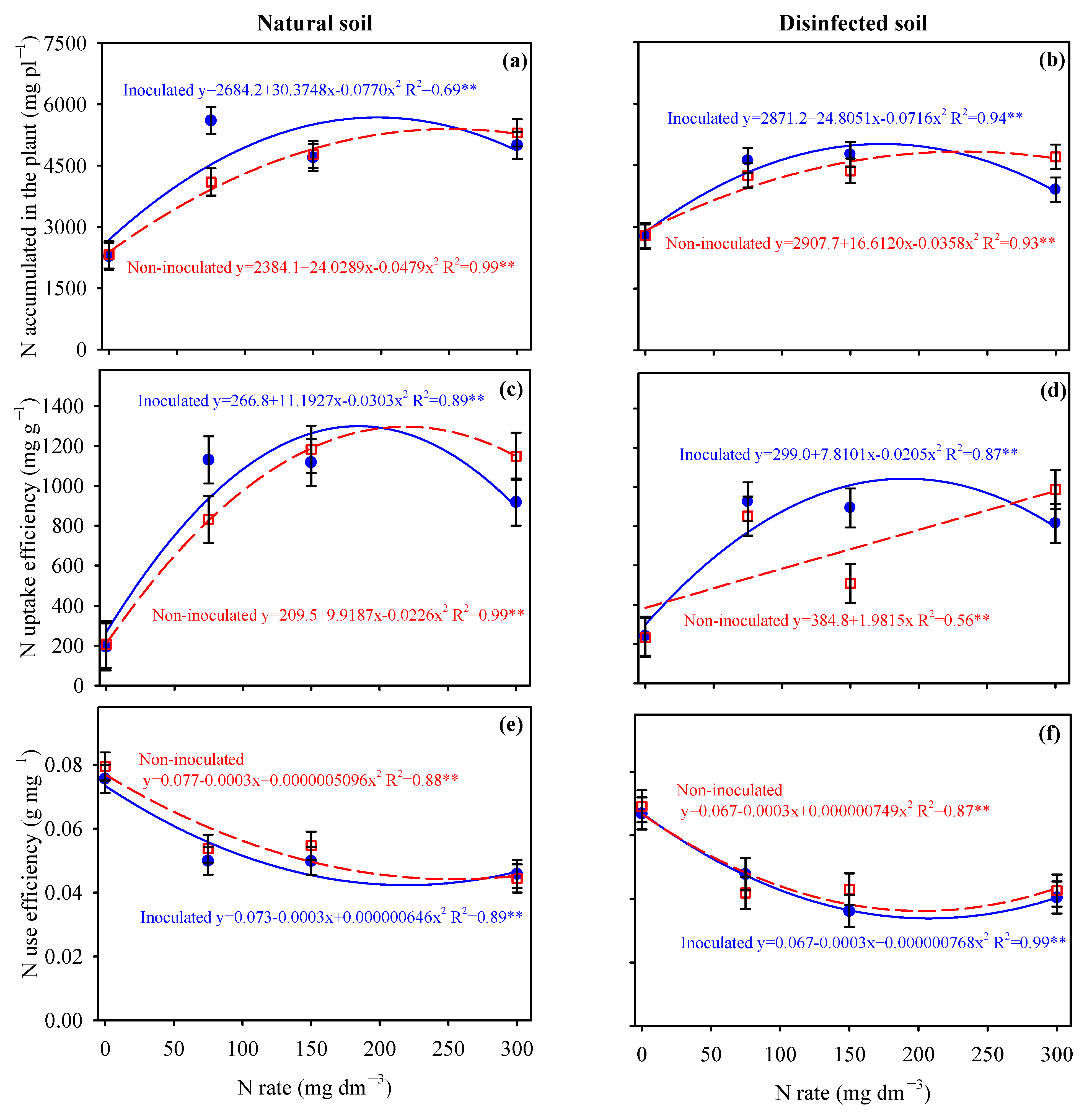
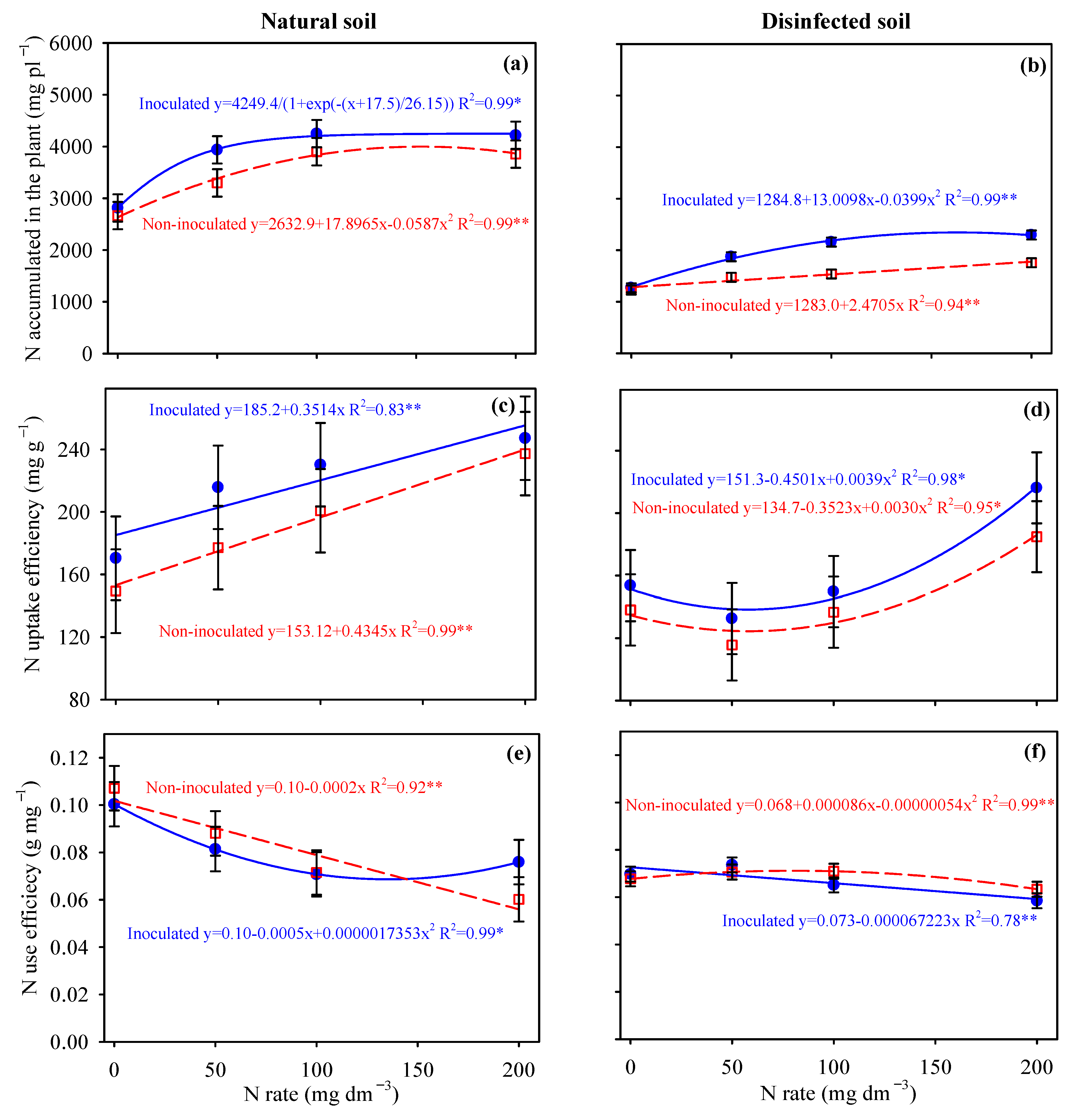
3.3. Nitrogen Uptake and N-Use Efficiency by Potato and Cassava Plants
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- FAOSTAT. Crops: Cassava and Potatoes. 2023. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 25 January 2023).
- Souza, E.F.C.; Soratto, R.P.; Fernandes, A.M.; Gupta, S.K. Performance of conventional and enhanced-efficiency nitrogen fertilizers on potato tuber mineral composition and marketability. J. Sci. Food Agric. 2022, 102, 3078–3087. [Google Scholar] [CrossRef] [PubMed]
- Mota, L.H.S.O.; Fernandes, A.M.; Assunção, N.S.; Leite, H.M.F. Leaf area development and yield of cassava in response to pruning of shoots and the late supply of nitrogen and potassium. Agron. J. 2020, 112, 1406–1422. [Google Scholar] [CrossRef]
- Fontes, P.C.R.; Braun, H.; Busato, C.; Cecon, P.R. Economic optimum nitrogen fertilization rates and nitrogen fertilization rate effects on tuber characteristics of potato cultivars. Potato Res. 2010, 53, 167–179. [Google Scholar] [CrossRef]
- Souza, E.F.C.; Soratto, R.P.; Fernandes, A.M.; Rosen, C.J. Nitrogen source and rate effects on irrigated potato in tropical sandy soils. Agron. J. 2019, 111, 378–389. [Google Scholar] [CrossRef]
- Uwah, D.F.; Effa, E.B.; Ekpenyong, L.E.; Akpan, I.E. Cassava (Manihot esculenta Crantz) performance as influenced by nitrogen and potassium fertilizers in Uyo, Nigeria. J. Anim. Plant Sci. 2013, 23, 550–555. [Google Scholar]
- Howeler, R.H.; Cadavid, L.F. Short- and long-term fertility trials in Colombia to determine the nutrient requirements of cassava. Fertil. Res. 1990, 26, 61–80. [Google Scholar] [CrossRef]
- Oliveira, N.T.; Uchôa, S.C.P.; Alves, J.M.A.; Albuquerque, J.A.A.; Rodrigues, G.S. Effect of harvest time and nitrogen doses on cassava root yield and quality. Rev. Bras. Cienc. Solo. 2017, 41, e0150204. [Google Scholar] [CrossRef]
- Galindo, F.S.; Buzetti, S.; Rodrigues, W.L.; Boleta, E.H.M.; Silva, V.M.; Tavanti, R.F.R.; Fernandes, G.C.; Biagini, A.L.C.; Rosa, P.A.L.; Teixeira Filho, M.C.M. Inoculation of Azospirillum brasilense associated with silicon as a liming source to improve nitrogen fertilization in wheat crops. Sci. Rep. 2020, 10, 6160. [Google Scholar] [CrossRef]
- Rodrigues, A.C.; Bonifacio, A.; Araujo, F.F.; Lira Junior, M.A.; Figueiredo, M.V.B. Azospirillum sp. as a challenge for agriculture. In Bacterial Metabolites in Sustainable Agroecosystem; Maheshwari, D.K., Ed.; Springer: Cham, Switzerland, 2015; Volume 12. [Google Scholar] [CrossRef]
- Boddey, R.M.; Döbereiner, J. Nitrogen fixation associated with grasses and cereals: Recent progress and perspectives for the future. Fertil. Res. 1995, 42, 241–250. [Google Scholar] [CrossRef]
- Pereg, L.; de-Bashan, L.E.; Bashan, Y. Assessment of affinity and specificity of Azospirillum for plants. Plant Soil. 2016, 399, 389–414. [Google Scholar] [CrossRef]
- Caires, E.F.; Bini, A.R.; Barão, L.F.C.; Haliski, A.; Duart, V.M.; Ricardo, K.S. Seed inoculation with Azospirillum brasilense and nitrogen fertilization for no-till cereal production. Agron. J. 2021, 113, 560–576. [Google Scholar] [CrossRef]
- Hungria, M.; Barbosa, J.Z.; Rondina, A.B.L.; Nogueira, M.A. Improving maize sustainability with partial replacement of N fertilizers by inoculation with Azospirillum brasilense. Agron. J. 2022, 114, 2969–2980. [Google Scholar] [CrossRef]
- Santos, M.S.; Nogueira, M.A.; Hungria, M. Outstanding impact of Azospirillum brasilense strains Ab-V5 and Ab-V6 on the Brazilian agriculture: Lessons that farmers are receptive to adopt new microbial inoculants. Rev. Bras. Ciênc. Solo. 2021, 45, e0200128. [Google Scholar] [CrossRef]
- Urrea-Valencia, S.; Etto, R.M.; Takahashi, W.Y.; Caires, E.F.; Bini, A.R.; Ayub, R.A.; Stets, M.I.; Cruz, L.M.; Galvão, C.W. Detection of Azospirillum brasilense by qPCR throughout a maize field trial. Appl. Soil Ecol. 2021, 160, 103849. [Google Scholar] [CrossRef]
- Koul, V.; Kochar, M. A novel essential small RNA, sSp_p6 influences nitrogen fixation in Azospirillum brasilense. Rhizosphere 2021, 17, 100281. [Google Scholar] [CrossRef]
- Cassán, F.; Maiale, S.; Masciarelli, O.; Vidal, A.; Luna, V.; Ruiz, O. Cadaverine production by Azospirillum brasilense and its possible role in plant growth promotion and osmotic stress mitigation. Eur. J. Soil Biol. 2009, 45, 12–19. [Google Scholar] [CrossRef]
- Majeed, A.; Muhammad, Z.; Ahmad, H. Plant growth promoting bacteria: Role in soil improvement; abiotic and biotic stress management of crops. Plant Cell Rep. 2018, 37, 1599–1609. [Google Scholar] [CrossRef]
- Kargapolova, K.Y.; Burygin, G.L.; Tkachenko, O.V.; Evseeva, N.V.; Pukhalskiy, Y.V.; Belimov, A.A. Effectiveness of inoculation of in vitro-grown potato microplants with rhizosphere bacteria of the genus Azospirillum. Plant Cell Tissue Organ Cult. 2020, 141, 351–359. [Google Scholar] [CrossRef]
- Tkachenko, O.V.; Evseeva, N.V.; Terentyeva, E.V.; Burygin, G.L.; Shirokov, A.A.; Burov, A.M.; Matora, L.Y.; Shchyogolev, S.Y. Improved production of high-quality potato seeds in aeroponics with plant-growth-promoting rhizobacteria. Potato Res. 2021, 64, 55–66. [Google Scholar] [CrossRef]
- Naqqash, T.; Hameed, S.; Imran, A.; Hanif, M.K.; Majeed, A.; van Elsas, J.D. Differential response of potato toward inoculation with taxonomically diverse plant growth promoting rhizobacteria. Front. Plant Sci. 2016, 7, 144. [Google Scholar] [CrossRef]
- Balota, E.L.; Lopes, E.S.; Hungria, M.; Döbereiner, J. Occurrence of diazotrophic bacteria and arbuscular mycorrhizal fungi on the cassava crop (in Portuguese, with English abstract). Pesq. Agropec. Bras. 1999, 34, 1265–1276. [Google Scholar] [CrossRef]
- Reinhardt, É.L.; Ramos, P.L.; Manfio, G.P.; Barbosa, H.R.; Pavan, C.; Moreira-Filho, C.A. Molecular characterization of nitrogen-fixing bacteria isolated from Brazilian agricultural plants at São Paulo state. Braz. J. Microbiol. 2008, 39, 414–422. [Google Scholar] [CrossRef] [PubMed]
- Balota, E.L.; Lopes, E.S.; Hungria, M.; Döbereiner, J. Interactions and physiological effects of diazotrophic bacteria and arbuscular mycorrhizal fungi in cassava plants (in Portuguese, with English abstract). Pesq. Agropec. Bras. 1995, 30, 1335–1345. [Google Scholar]
- Balota, E.L.; Lopes, E.S.; Hungria, M.; Döbereiner, J. Inoculation of diazotrophic bacteria and arbuscular mycorrhizal fungi on the cassava crop (in Portuguese; with English abstract). Pesq. Agropec. Bras. 1997, 32, 627–639. [Google Scholar]
- Hridya, A.C.; Byju, G.; Misra, R.S. Effect of biocontrol agents and biofertilizers on root rot, yield, harvest index and nutrient uptake of cassava (Manihot esculanta Crantz). Arch. Agron. Soil Sci. 2013, 59, 1215–1227. [Google Scholar] [CrossRef]
- Hungria, M.; Campo, R.J.; Souza, E.M.; Pedrosa, F.O. Inoculation with selected strains of Azospirillum brasilense and A. lipoferum improves yields of maize and wheat in Brazil. Plant Soil. 2010, 331, 413–425. [Google Scholar] [CrossRef]
- Gazola, B.; Fernandes, A.M.; Hellmeister, G.; Abrami, L.S.; Silva, R.M.; Soratto, R.P. Potassium management effects on yield and quality of cassava varieties in tropical sandy soils. Crop. Pasture Sci. 2022, 73, 285–299. [Google Scholar] [CrossRef]
- Buensanteai, N.; Sompong, M.; Thamnu, K.; Athinuwat, D.; Brauman, A.; Plassard, C. The plant growth promoting bacterium Bacillus sp. CaSUT007 produces phytohormone and extracellular proteins for enhanced growth of cassava. Afr. J. Microbiol. Res. 2013, 7, 4949–4954. [Google Scholar] [CrossRef]
- Ferreira, D.F. Sisvar: A computer statistical analysis system. Ciênc. Agrotec. 2011, 35, 1039–1042. [Google Scholar] [CrossRef]
- Zebarth, B.J.; Rosen, C.J. Research perspective on nitrogen BMP development for potato. Am. J. Potato Res. 2007, 84, 3–18. [Google Scholar] [CrossRef]
- Saubidet, M.I.; Fatta, N.; Barneix, A.J. The effect of inoculation with Azospirillum brasilense on growth and nitrogen utilization by wheat plants. Plant Soil. 2002, 245, 215–222. [Google Scholar] [CrossRef]
- Howeler, R. Mineral Nutrition and Fertilization of Cassava; Centro Internacional de Agricultura Tropical: Cali, Colombia, 1981; p. 52. [Google Scholar]
- Tsay, J.S.; Fukai, S.; Wilson, G.L. Growth and yield of cassava as influenced by intercropped soybean and by nitrogen application. Field Crop. Res. 1989, 21, 83–94. [Google Scholar] [CrossRef]
- Groppa, M.D.; Benavides, M.P.; Zawoznik, M.S. Root hydraulic conductance; aquaporins and plant growth promoting microorganisms: A revision. Appl. Soil Ecol. 2012, 61, 247–254. [Google Scholar] [CrossRef]
- Richardson, A.E.; Barea, J.M.; McNeill, A.M.; Prigent-Combaret, C. Acquisition of phosphorus and nitrogen in the rhizosphere and plant growth promotion by microorganisms. Plant Soil. 2009, 321, 305–339. [Google Scholar] [CrossRef]
- Bashan, Y.; Holguin, G.; de-Bashan, L.E. Azospirillum-plant relationships: Physiological, molecular, agricultural, and environmental advances. 1997–2003. Can. J. Microbiol. 2004, 50, 521–577. [Google Scholar] [CrossRef]
- Fukami, J.; Ollero, F.J.; Megías, M.; Hungria, M. Phytohormones and induction of plant-stress tolerance and defense genes by seed and foliar inoculation with Azospirillum brasilense cells and metabolites promote maize growth. AMB Express 2017, 7, 153. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Characteristic | Values | |
|---|---|---|
| Potato | Cassava | |
| pH(CaCl2) | 6.0 | 5.6 |
| Content of oxidizable carbon (Cox) (%) | 0.30 | 0.73 |
| Presin-extractable (mg dm−3) | 161.0 | 141.0 |
| H+Al (mmolc dm−3) | 23.0 | 25.0 |
| K (mmolc dm−3) | 3.7 | 3.8 |
| Ca (mmolc dm−3) | 40.0 | 45.0 |
| Mg (mmolc dm−3) | 20.0 | 14.0 |
| Cation exchange capacity (mmolc dm−3) | 86.7 | 87.8 |
| Base saturation (%) | 73.5 | 71.5 |
| B (mg dm−3) | 0.6 | 1.6 |
| Cu (mg dm−3) | 4.6 | 3.5 |
| Fe (mg dm−3) | 14.0 | 10.0 |
| Mn (mg dm−3) | 4.0 | 2.8 |
| Zn (mg dm−3) | 0.8 | 1.7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernandes, A.M.; da Silva, J.A.; Eburneo, J.A.M.; Leonel, M.; Garreto, F.G.d.S.; Nunes, J.G.d.S. Growth and Nitrogen Uptake by Potato and Cassava Crops Can Be Improved by Azospirillum brasilense Inoculation and Nitrogen Fertilization. Horticulturae 2023, 9, 301. https://doi.org/10.3390/horticulturae9030301
Fernandes AM, da Silva JA, Eburneo JAM, Leonel M, Garreto FGdS, Nunes JGdS. Growth and Nitrogen Uptake by Potato and Cassava Crops Can Be Improved by Azospirillum brasilense Inoculation and Nitrogen Fertilization. Horticulturae. 2023; 9(3):301. https://doi.org/10.3390/horticulturae9030301
Chicago/Turabian StyleFernandes, Adalton Mazetti, Jessica Aparecida da Silva, Juliana Aparecida Marques Eburneo, Magali Leonel, Francisca Gyslane de Sousa Garreto, and Jason Geter da Silva Nunes. 2023. "Growth and Nitrogen Uptake by Potato and Cassava Crops Can Be Improved by Azospirillum brasilense Inoculation and Nitrogen Fertilization" Horticulturae 9, no. 3: 301. https://doi.org/10.3390/horticulturae9030301
APA StyleFernandes, A. M., da Silva, J. A., Eburneo, J. A. M., Leonel, M., Garreto, F. G. d. S., & Nunes, J. G. d. S. (2023). Growth and Nitrogen Uptake by Potato and Cassava Crops Can Be Improved by Azospirillum brasilense Inoculation and Nitrogen Fertilization. Horticulturae, 9(3), 301. https://doi.org/10.3390/horticulturae9030301