
Optimization of Morphogenesis and In Vitro Production of Five Hyacinthus orientalis Cultivars


Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Sterilization
2.3. Micropropagation and Rooting
2.4. Environmental Culture Conditions
2.5. Chlorophyll Content
2.6. Growth Parameters
2.7. Statistical Design and Data Analysis
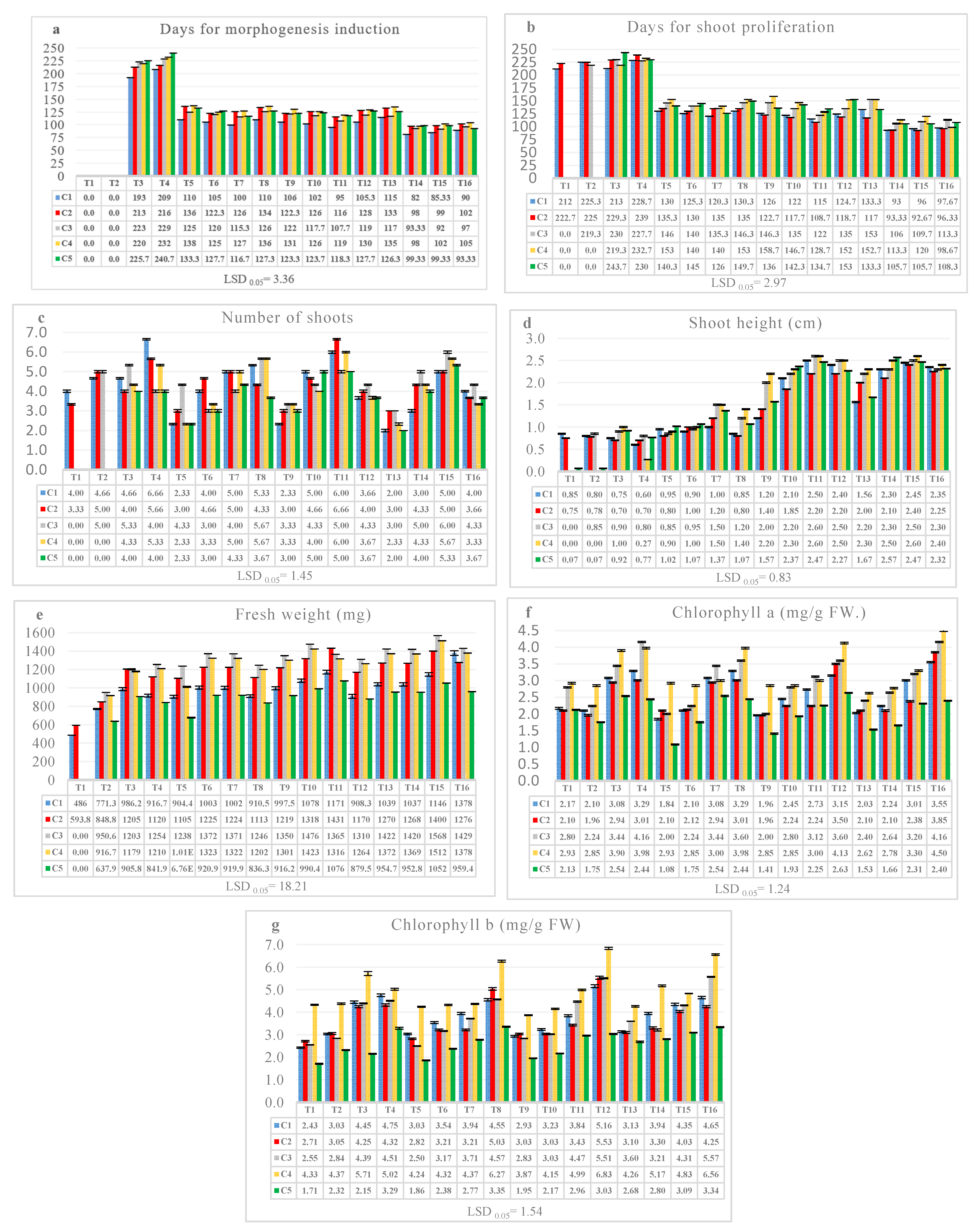
3. Results
3.1. Effects of the Five Cultivars, IAA, and Either BA or Kin in Experiments 1 and 2 on Growth Parameters
3.2. Influence of the Interactions between the Five Cultivars, IAA, and Either BA or Kin in Both Experiments 1 and 2 on Growth Parameters
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- The Angiosperm Phylogeny Group. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IIIboj_996 105. Bot. J. Linn. Soc. 2009, 161, 105–121. [Google Scholar] [CrossRef]
- Wong, K.H.; Wu, H.; Bobby, L.K.; Grace, W.B.; Siu, T.; Jerome, H.H.; Shaw, P.; David, T.L. Characterisation of the complete chloroplast genomes of seven Hyacinthus orientalis L. cultivars: Insights into cultivar phylogeny. Horticulturae 2022, 8, 453. [Google Scholar] [CrossRef]
- Young, B.Y.; Kyeoung, S.L.; Gung-Han, C. Protein variation and efficient in vitro culture of scale segments from Hyacinthus orientalis L. c.v. Carnegie. Sci. Hortic. 2002, 92, 367–374. [Google Scholar] [CrossRef]
- Miyamotoa, K.; Kotakeb, T.; Boncelac, A.J.; Saniewskic, M.; Uedad, J. Hormonal regulation of gummosis and composition of gums from bulbs of hyacinth (Hyacinthus orientalis). J. Plant Physiol. 2015, 174, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Saniewski, M.; Nowak, J.; Rudnicki, R. Studies on the physiology of Hyacinth bulbs (Hyacinthus orientalis L.) IV. Hormonal regulation of induction of roots and bulblets in Hyacinthus orientalis grown in culture. Plant Sci. Lett. 1974, 2, 373–376. [Google Scholar] [CrossRef]
- Pierik, R.L.M.; Post, A.J.M. Rapid vegetative propagation of Hyacinthus orientalis L. in vitro. Sci. Hortic. 1975, 3, 293–297. [Google Scholar] [CrossRef]
- Shinsaku, T.; Takako, A.; Mayumi, F. Rapid clonal propagation of Hyacinthus orientalis bulbs by shake culture. Sci. Hortic. 1991, 45, 315–321. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with Tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Moran, R. Formulae for determination of chlorophyllous pigments extracted with N,N-Dimethylformamide. Plant Physiol. 1982, 69, 1376–1381. [Google Scholar] [CrossRef]
- Gomez, A.A.; Gomez, K.A. Statistical Procedures for Agricultural Research, 2nd ed.; John Wiley and Sons: Hoboken, NJ, USA, 1984; 680p. [Google Scholar] [CrossRef]
- Tukey, J.W. Comparing individual means in the analysis of variance. Biometrics 1949, 2, 99–114. [Google Scholar] [CrossRef]
- Marijana, S.; Suzana, Z.; Jelena, S.; Branislav, S.; Aneta, S.; Sladjana, T.; Dragoljub, G. Efficient one-step tissue culture protocol for propagation of endemic plant, Lilium martagon var. cattaniae Vis. Afr. J. Biotechnol. 2012, 11, 1862–1867. [Google Scholar] [CrossRef]
- Karakas, P.F. Efficient plant regeneration and callus induction from nodal and hypocotyl explants of goji berry (Lycium barbarum L.) and comparison of phenolic profiles in calli formed under different combinations of plant growth regulators. Plant Physiol. Biochem. 2020, 146, 384–391. [Google Scholar] [CrossRef] [PubMed]
- Volfová, A.L.; Chvojka, A.F. The effect of kinetin and auxin on the chloroplast structure and chlorophyll content in wheat coleoptiles. Biol. Plant. 1978, 20, 440–445. [Google Scholar] [CrossRef]
- Liorente, B.E.; Apóstolo, N.M. Effect of different growth regulators and genotype on in vitro propagation of jojoba. N. Z. J. Crop Hortic. Sci. 1998, 26, 55–62. [Google Scholar] [CrossRef]
- Bhat, M.S.; Zahoor, A.R.; Imtiyaz, T.N.; Neelofar, B.; Tabinda, W.; Sadaf, R.; Iqra, F.; Ahmed, N.; Hadeer, D. Standardization of in vitro micropropagation of winter jasmine (Jasminum nudiflorum) using nodal explants. Saudi J. Biol. Sci. 2022, 29, 3425–3431. [Google Scholar] [CrossRef]
- El-Naggar, H.; Osman, A.; Sewedan, E. In vitro propagation and organogenesis of Lilium ‘Prato’. Afr. J. Biotechnol. 2012, 11, 14771–14776. [Google Scholar] [CrossRef]
- Han, B.H.; Hee, J.Y.; Byeoung, W.Y.; Kee, Y.P. In vitro micropropagation of Lilium longiflorum ‘Georgia’ by shoot formation as influenced by addition of liquid medium. Sci. Hortic. 2004, 103, 39–49. [Google Scholar] [CrossRef]
- Loretta, B.; Patrizio, C.R.; Claudia, B.; Francesco, S. Adventitious shoot regeneration from leaf explants and stem nodes of Lilium. Plant Cell Tiss. Org. Cult. 2003, 74, 37–44. [Google Scholar] [CrossRef]
- Godo, T.; Katsunori, K.; Tomoyuki, T.; Kazuhiko, M.; Takao, K. In vitro propagation utilizing suspension cultures of meristematic nodular cell clumps and chromosome stability of Lilium formolongi. Sci. Hortic. 1998, 72, 193–202. [Google Scholar] [CrossRef]
- Economou, A.S.; Read, P.E. Effect of benzyladenine pretreatments on shoot proliferation from petunia leaf segments cultured in vitro. In Proceedings of the Plant Growth Regulator Working Group; Annual meeting (USA); FAO: Rome, Italy, 1980. [Google Scholar]
- Hosokawa, K. Hyacinthus orientalis L.: In vitro culture and the production of anthocyanin and other secondary metabolites. Biotechnol. Agric. Med. Aromat. Plants XI 1999, 43, 177–198. [Google Scholar] [CrossRef]
- Hassan, K.; Hosni, A.; Hewidy, M.; Abd Elrazik, A. Micropropagation and evaluation of genetic stability of foxglove tree (Paulownia tomentosa). Arab Univ. J. Agri. Sci. 2019, 26, 2287–2296. [Google Scholar] [CrossRef]
- Nhut, D.T.; Nguyen, T.D.; Vu, Q.L.; Nguyen, Q.T.; Nguyen, T.M.; Thai, X.D.; Bui, V.L. Standardization of in-vitro lily (Lilium spp.) plantlets for propagation and bulb formation. Proc. Int. Workshop Biotechnol. Agric. 2006, 20–21, 134–138. [Google Scholar]
- Arab, M.M.; Yadollahi, A.; Shojaeiyan, A.; Shokri, S.; Ghojah, S.M. Effects of nutrient media, different cytokinin types and their concentrations on in vitro multiplication of G × N15 (hybrid of almond × peach) vegetative rootstock. J. Genetic Eng. Biotechnol. 2014, 12, 81–87. [Google Scholar] [CrossRef]
- Zhoua, J.; Kun, C.; Guomin, H.; Guangcai, C.; Shoubiao, Z.; Yongjie, H.; Jie, Z.; Honglang, D.; Houbao, F. Effects of exogenous 3-indoleacetic acid and cadmium stress on the physiological and biochemical characteristics of Cinnamomum camphora. Ecotoxicol. Environ. Saf. 2020, 191, 15. [Google Scholar] [CrossRef]
- Nana, L. Effects of IAA and ABA on the immature peach fruit development process. Hortic. Plant J. 2019, 5, 145–154. [Google Scholar] [CrossRef]
- Sadaf, R.; Rather, Z.A.; Reyaz, A.B.; Nazki, I.T.; Mohammad, S.A.; Neelofar, B.; Iqra, F.; Bassem, N.S.; Khan, M.H.; Atef, F.; et al. Standardization of in vitro micropropagation procedure of Oriental Lilium Hybrid Cv. Saudi J. Bio. Sci. 2021, 28, 7581–7587. [Google Scholar] [CrossRef]
- Gheisari, M.; Miri, S.M. In vitro Callus induction and bulblet regeneration of hyacinth (Hyacinthus orientalis L.). Plant Cell Biotechnol. Mol. Biol. 2017, 18, 145–155. [Google Scholar]
- Linlin, Y.; Liqin, C.; Xiuhong, S.; Chunyan, W.; Chengming, D.; Suiqing, C. High-frequency in vitro plantlet regeneration in Lilium davidii var. unicolour Salisb, an important edible and medicinal plant, and confirmation of genetic fidelity of regeneration plantlets using ISSR markers. Plant Biotechnol. Rep. 2021, 15, 435–446. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Days for Morphogenesis | Days for Shoot Proliferation | Number of Shoots | Shoot Height (cm) | FW. (mg) | Chl. a (mg/g FW) | Chl. b (mg/g FW) | |
|---|---|---|---|---|---|---|---|
| Main effect of the 5 cultivars | |||||||
| Pink Pearl | 132.97 ± 3.8 d | 143.29 ± 3.0 d | 4.45 ± 0.10 a | 1.47 ± 0.05 c | 953.10 ± 23.0 c | 2.64 ± 0.04 c | 3.81 ± 0.06 b |
| Jan Bos | 134.48 ± 2.1 d | 144.90 ± 3.3 d | 4.45 ± 0.08 a | 1.44 ± 0.04 c | 1136.95 ± 20.9 b | 2.53 ± 0.04 c | 3.64 ± 0.06 b |
| Blue Pearl | 157.75 ± 5.6 c | 164.06 ± 4.2 c | 4.17 ± 0.10 a | 1.60 ± 0.05 a | 1248.40 ± 22.8 a | 3.04 ± 0.05 b | 3.79 ± 0.06 b |
| Serene Blue | 165.25 ± 5.5 a | 176.79 ± 5.0 a | 4.17 ± 0.13 a | 1.59 ± 0.06 ab | 1193.59 ± 22.2 ab | 3.28 ± 0.04 a | 4.96 ± 0.07 a |
| White Pearl | 162.67 ± 5.5 b | 173.35 ± 5.2 b | 3.31 ± 0.10 b | 1.50 ± 0.05 bc | 845.04 ± 16.0 d | 2.03 ± 0.03 d | 2.60 ± 0.04 c |
| Main effect of IAA | |||||||
| 0.0 mg/L | 258.07 ± 4.7 a | 259.90 ± 3.8 a | 3.50 ± 0.14 c | 0.58 ± 0.02 d | 747.35 ± 30.2 d | 2.61 ± 0.05 b | 3.60 ± 0.08 b |
| 0.5 mg/L | 123.03 ± 0.7 b | 137.80 ± 0.6 b | 4.08 ± 0.08 b | 1.06 ± 0.02 c | 1096.1 ± 13.9 c | 2.64 ± 0.05 b | 3.64 ± 0.07 ab |
| 1.0 mg/L | 118.30 ± 0.6 c | 132.28 ± 0.9 c | 4.46 ± 0.09 a | 2.17 ± 0.03 b | 1198.0 ± 13.8 b | 2.72 ± 0.05 b | 3.80 ± 0.09 ab |
| 1.5 mg/L | 103.10 ± 0.9 d | 111.95 ± 1.2 d | 4.20 ± 0.09 b | 2.27 ± 0.02 a | 1260.2 ± 14.6 a | 2.87 ± 0.05 a | 4.00 ± 0.07 a |
| Main effect of BA | |||||||
| 0.0 mg/L | 167.33 ± 5.6 a | 179.90 ± 5.2 a | 2.50 ± 0.09 c | 1.20 ± 0.04 d | 838.8 ± 34.0 c | 2.13 ± 0.04 d | 2.97 ± 0.05 d |
| 1.0 mg/L | 157.63 ± 6.0 b | 162.73 ± 5.0 b | 3.80 ± 0.10 b | 1.49 ± 0.05 c | 1115.0 ± 16.6 b | 2.42 ± 0.03 c | 3.31 ± 0.06 c |
| 2.0 mg/L | 134.67 ± 3.1 d | 146.25 ± 3.2 c | 5.10 ± 0.07 a | 1.78 ± 0.05 a | 1217.7 ± 13.9 a | 2.84 ± 0.04 b | 3.96 ±0.06 b |
| 3.0 mg/L | 142.86 ± 3.2 c | 153.50 ± 3.2 d | 5.00 ± 0.07 a | 1.60 ± 0.05 b | 1130.1 ± 14.4 b | 3.42 ± 0.04 a | 4.80 ± 0.08 a |
| Days for Morphogenesis | Days for Shoot Proliferation | Number of Shoots | Shoot Height (cm) | FW. (mg) | Chl. a (mg/g FW) | Chl. b (mg/g FW) | |
|---|---|---|---|---|---|---|---|
| Main effect of the 5 cultivars | |||||||
| Pink pearl | 165.27 ± 5.9 d | 156.39 ± 3.8 c | 3.13 ± 0.08 a | 1.27 ± 0.04 a | 727.69 ± 9.6 c | 2.20 ± 0.04 b | 3.33 ± 0.07 b |
| Jan Bos | 182.08 ± 5.6 c | 157.90 ± 4.1 c | 3.27 ± 0.07 a | 1.24 ± 0.04 a | 799.40 ± 10.7 b | 2.20 ± 0.04 b | 3.19 ± 0.07 b |
| Blue Pearl | 189.87 ± 5.4 ab | 178.10 ± 4.4 b | 2.99 ± 0.08 ab | 1.38 ± 0.05 a | 949.40 ± 10.5 a | 2.70 ± 0.05 a | 3.35 ± 0.08 b |
| Serene Blue | 191.30 ± 5.3 a | 184.40 ± 5.4 a | 2.67 ± 0.10 bc | 1.36 ± 0.06 a | 943.51 ± 16.2 a | 2.82 ± 0.04 a | 3.97 ± 0.05 a |
| White Pearl | 188.83 ± 5.5 b | 185.02 ± 5.3 a | 2.39 ± 0.08 c | 1.23 ± 0.05 a | 663.02 ± 10.7 c | 1.91 ± 0.04 b | 2.27 ± 0.04 c |
| Main effect of IAA | |||||||
| 0.0 mg/L | 318.26 ± 2.8 a | 280.64 ± 3.0 a | 2.62 ± 0.10 b | 0.48 ± 0.02 c | 652.05 ± 13.9 c | 2.53 ± 0.06 a | 3.42 ± 0.08 a |
| 0.5 mg/L | 158.02 ± 0.9 b | 156.52 ± 0.7 b | 2.91 ± 0.07 ab | 0.90 ± 0.01 b | 809.93 ± 10.8 b | 2.31 ± 0.05 a | 3.15 ± 0.07 a |
| 1.0 mg/L | 152.32 ± 0.8 c | 145.97 ± 1.7 c | 3.13 ± 0.08 a | 1.85 ± 0.03 a | 898.08 ± 12.1 a | 2.30 ± 0.05 a | 3.14 ± 0.07 a |
| 1.5 mg/L | 105.28 ± 2.2 d | 106.31 ± 2.1 d | 2.91 ± 0.07 ab | 1.94 ± 0.02 a | 906.36 ± 12.1 a | 2.30 ± 0.05 a | 3.17 ± 0.07 a |
| Main effect of Kin | |||||||
| 0.0 mg/L | 210.24 ± 5.7 a | 196.69 ± 4.8 a | 1.69 ± 0.06 c | 1.03 ± 0.04 c | 784.51 ± 18.7 a | 1.87 ± 0.04 c | 2.54 ± 0.05 c |
| 1.0 mg/L | 188.04 ± 6.8 b | 171.35 ± 5.3 b | 2.80 ± 0.08 b | 1.28 ± 0.05 b | 835.83 ± 12.2 a | 2.11 ± 0.04 c | 2.89 ± 0.06 c |
| 2.0 mg/L | 162.58 ± 4.4 d | 153.06 ± 4.3 d | 3.72 ± 0.06 a | 1.52 ± 0.04 a | 809.73 ± 11.6 a | 2.45 ± 0.04 b | 3.38 ± 0.06 b |
| 3.0 mg/L | 173.02 ± 4.6 c | 168.35 ± 3.9 c | 3.35 ± 0.06 a | 1.35 ± 0.05 b | 836.35 ± 11.8 a | 3.02 ± 0.05 a | 4.08 ± 0.08 a |
| Percentage of Root Formation (%) | Number of Roots per Plantlet | Root Length (cm) | |
|---|---|---|---|
| Pink Pearl | 81.1 ± 13.6 a | 3.00 ± 2.12 a | 0.76 ± 0.41 ab |
| Jan Bos | 83.3 ± 12.0 a | 3.66 ± 2.34 a | 0.77 ± 0.46 a |
| Blue Pearl | 74.4 ± 14.2 ab | 3.22 ± 1.92 a | 0.62 ± 0.31 ab |
| Serene Blue | 82.2 ± 13.9 a | 2.88 ± 1.53 ab | 0.70 ± 0.32 b |
| White Pearl | 71.1 ± 10.5 b | 2.00 ± 1.58 b | 0.39 ± 0.27 c |
| Main effect of IBA | |||
| 1.0 mg/L | 68.6 ± 7.4 b | 1.40 ± 0.82 a | 0.43 ± 0.13 a |
| 2.0 mg/L | 73.3 ± 9.0 b | 2.20 ± 2.20 b | 0.42 ± 0.14 a |
| 3.0 mg/L | 93.3 ± 8.0 a | 5.26 ± 5.26 c | 1.10 ± 0.26 b |
| IBA (mg/L) | Percentage of Root Formation (%) | Number of Roots per Plantlet | Root Length (cm) | |
|---|---|---|---|---|
| Pink Pearl | 1.0 | 70.0 ± 10.0 g | 1.33 ± 0.58 fg | 0.43 ± 0.05 cd |
| 2.0 | 76.6 ± 5.77 e | 2.00 ± 1.00 ef | 0.50 ± 0.10 cd | |
| 3.0 | 96.6 ± 5.77 b | 5.66 ± 0.58 ab | 1.39 ± 0.06 a | |
| Jan Bos | 1.0 | 73.3 ± 5.77 f | 1.66 ± 0.58 ef | 0.53 ± 0.11 cd |
| 2.0 | 76.6 ± 5.77 e | 2.66 ± 0.58 de | 0.46 ± 0.05 cd | |
| 3.0 | 100 ± 0.00 a | 6.66 ± 0.58 a | 1.29 ± 0.08 a | |
| Blue Pearl | 1.0 | 70.0 ± 10.0 g | 2.00 ± 0.00 ef | 0.40 ± 0.10 cd |
| 2.0 | 63.3 ± 5.77 i | 2.00 ± 1.00 ef | 0.46 ± 0.15 cd | |
| 3.0 | 90.0 ± 10.0 c | 5.66 ± 0.58 ab | 0.99 ± 0.19 ab | |
| Serene Blue | 1.0 | 66.6 ± 5.77 h | 1.66 ± 1.15 ef | 0.53 ± 0.05 cd |
| 2.0 | 83.3 ± 5.77 d | 2.33 ± 0.58 ef | 0.46 ± 0.05 cd | |
| 3.0 | 96.6 ± 5.77 b | 4.66 ± 0.58 bc | 1.11 ± 0.18 ab | |
| White Pearl | 1.0 | 63.3 ± 5.77 i | 0.33 ± 0.58 g | 0.26 ± 0.15 d |
| 2.0 | 66.6 ± 5.77 h | 2.00 ± 1.00 ef | 0.20 ± 0.10 d | |
| 3.0 | 83.3 ± 5.77 d | 3.66 ± 0.58 cd | 0.73 ± 0.06 bc | |
| LSD 0.05 | 3.20 | 1.05 | 0.41 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Naggar, H.M.; Shehata, A.M.; Moubarak, M.; Osman, A.R. Optimization of Morphogenesis and In Vitro Production of Five Hyacinthus orientalis Cultivars. Horticulturae 2023, 9, 176. https://doi.org/10.3390/horticulturae9020176
El-Naggar HM, Shehata AM, Moubarak M, Osman AR. Optimization of Morphogenesis and In Vitro Production of Five Hyacinthus orientalis Cultivars. Horticulturae. 2023; 9(2):176. https://doi.org/10.3390/horticulturae9020176
Chicago/Turabian StyleEl-Naggar, Hany M., Ashraf M. Shehata, Maneea Moubarak, and Amira R. Osman. 2023. "Optimization of Morphogenesis and In Vitro Production of Five Hyacinthus orientalis Cultivars" Horticulturae 9, no. 2: 176. https://doi.org/10.3390/horticulturae9020176
APA StyleEl-Naggar, H. M., Shehata, A. M., Moubarak, M., & Osman, A. R. (2023). Optimization of Morphogenesis and In Vitro Production of Five Hyacinthus orientalis Cultivars. Horticulturae, 9(2), 176. https://doi.org/10.3390/horticulturae9020176