
Seaweed Extracts as Substitutes of Synthetic Hormones for Rooting Promotion in Rose Cuttings




Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Treatments
2.2. Biometric Measures
2.3. Carbohydrates Analysis
2.4. Statistics
3. Results
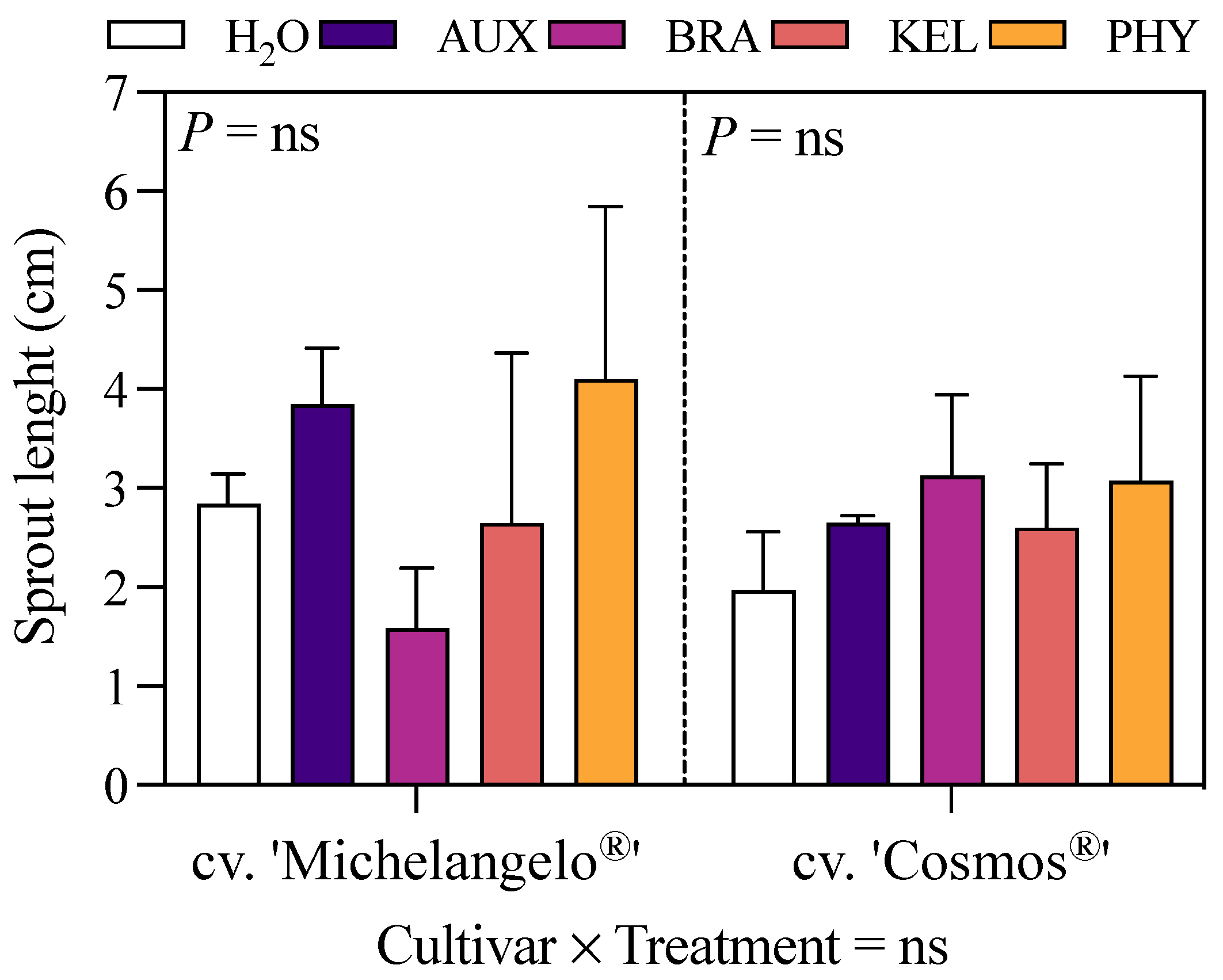
3.1. Survival Percentage and Biometric Measures
3.2. Soluble Sugars
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cardoso, J.C.; Vendrame, W.A. Innovation in propagation and cultivation of ornamental plants. Horticulturae 2022, 8, 229. [Google Scholar] [CrossRef]
- Kentelky, E.; Jucan, D.; Cantor, M.; Szekely-Varga, Z. Efficacy of different concentrations of NAA on selected ornamental woody shrubs cuttings. Horticulturae 2021, 7, 464. [Google Scholar] [CrossRef]
- Akhtar, M.S.; Khan, M.A.; Riaz, A.; Younis, A. Response of different rose species to different root promoting hormones. Pak. J. Agric. Sci. 2002, 39, 297–299. [Google Scholar]
- Nguyen, T.H.N.; Tänzer, S.; Rudeck, J.; Winkelmann, T.; Debener, T. Genetic analysis of adventitious root formation in vivo and in vitro in a diversity panel of roses. Sci. Hortic. 2020, 266, 109277. [Google Scholar] [CrossRef]
- Zhang, W.; Fan, J.; Tan, Q.; Zhao, M.; Cao, F. Mechanisms underlying the regulation of root formation in Malus hupehensis stem cuttings by using exogenous hormones. J. Plant Growth Regul. 2017, 36, 174–185. [Google Scholar] [CrossRef]
- Parađiković, N.; Teklić, T.; Zeljković, S.; Lisjak, M.; Špoljarević, M. Biostimulants research in some horticultural plant species—A review. Food Energy Secur. 2019, 8, e00162. [Google Scholar] [CrossRef]
- Blythe, E.K.; Sibley, J.L.; Tilt, K.M.; Ruter, J.M. Methods of auxin application in cutting propagation: A review of 70 years of scientific discovery and commercial practice. J. Environ. Hortic. 2007, 25, 166–185. [Google Scholar] [CrossRef]
- Betti, C.; Della Rovere, F.; Piacentini, D.; Fattorini, L.; Falasca, G.; Altamura, M.M. Jasmonates, ethylene and brassinosteroids control adventitious and lateral rooting as stress avoidance responses to heavy metals and metalloids. Biomolecules 2021, 11, 77. [Google Scholar] [CrossRef]
- Vardhini, B.V. Modifications of morphological and anatomical characteristics of plants by application of brassinosteroids under various abiotic stress conditions—A review. Plant Gene 2017, 11, 70–89. [Google Scholar] [CrossRef]
- Pacholczak, A.; Zajączkowska, M.; Nowakowska, K. The Effect of brassinosteroids on rooting of stem cuttings in two barberry (Berberis thunbergii L.) cultivars. Agronomy 2021, 11, 699. [Google Scholar] [CrossRef]
- Moreno-Castillo, E.; Ramírez-Echemendía, D.P.; Hernández-Campoalegre, G.; Mesa-Tejeda, D.; Coll-Manchado, F.; Coll-García, Y. In silico identification of new potentially active brassinosteroid analogues. Steroids 2018, 138, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Geelen, D. Developing biostimulants from agro-food and industrial by-products. Front. Plant Sci. 2018, 9, 1567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brzozowski, L.; Mazourek, M.A. Sustainable agricultural future relies on the transition to organic agroecological pest management. Sustainability 2018, 10, 2023. [Google Scholar] [CrossRef] [Green Version]
- Baltazar, M.; Correia, S.; Guinan, K.J.; Sujeeth, N.; Bragança, R.; Gonçalves, B. Recent advances in the molecular effects of biostimulants in plants: An overview. Biomolecules 2021, 11, 1096. [Google Scholar] [CrossRef]
- Dmytryk, A.; Chojnacka, K. Algae as fertilizers, biostimulants, and regulators of plant growth. In Algae biomass: Characteristics and Applications; Chojnacka, K., Wieczorek, P.P., Schroeder, G., Michalak, I., Eds.; Springer: Cham, Switzerland, 2018; pp. 115–122. [Google Scholar]
- Patel, J.S.; Mukherjee, A. Seaweed and associated products: Natural biostimulant for improvement of plant health. In Emerging Trends in Plant Pathology; Singh, K.P., Jahagirdar, S., Sarma, B.K., Eds.; Springer: Singapore, 2021; pp. 317–330. [Google Scholar]
- Ali, O.; Ramsubhag, A.; Jayaraman, J. Biostimulant properties of seaweed extracts in plants: Implications towards sustainable crop production. Plants 2021, 10, 531. [Google Scholar] [CrossRef]
- Kisvarga, S.; Farkas, D.; Boronkay, G.; Neményi, A.; Orlóci, L. Effects of biostimulants in horticulture, with emphasis on ornamental plant production. Agronomy 2022, 12, 1043. [Google Scholar] [CrossRef]
- Otiende, M.A.; Nyabundi, J.O.; Ngamau, K.; Opala, P. Effects of cutting position of rose rootstock cultivars on rooting and its relationship with mineral nutrient content and endogenous carbohydrates. Sci. Hortic. 2017, 225, 204–212. [Google Scholar] [CrossRef]
- Vazquez-Iglesias, I.; Ochoa-Corona, F.M.; Tang, J.; Robinson, R.; Clover, G.R.; Fox, A.; Boonham, N. Facing Rose rosette virus: A risk to European rose cultivation. Plant Pathol. 2020, 69, 1603–1617. [Google Scholar] [CrossRef]
- Al-Saqri, F.; Alderson, P.G. Effects of IBA, cutting type and rooting media on rooting of Rosa centifolia. J. Hortic. Sci. 1996, 71, 729–737. [Google Scholar] [CrossRef]
- Pati, P.K.; Prakash, O.; Sharma, M.; Sood, A.; Ahuja, P.S. Growth performance of cuttings raised from in vitro and in vivo propagated stock plants of Rosa damascena Mill. Biol. Plant. 2004, 48, 609–611. [Google Scholar] [CrossRef]
- Bannoud, F.; Bellini, C. Adventitious rooting in Populus species: Update and perspectives. Front. Plant Sci. 2021, 12, 918. [Google Scholar] [CrossRef]
- Farazi, E.; Afshari, H.; Hokm Abadi, H. Effect of different concentrations of brassinosteroid on physiomorphological characteristics of five pistachio genotypes (Pistacia vera L). J. Nuts 2015, 6, 143–153. [Google Scholar]
- Costa, J.M.; Heuvelink, E.; Van de Pol, P. Propagation by cuttings. In Reference Module in Life Sciences; Roitberg, B.D., Ed.; Elsevier: Amsterdam, The Netherlands, 2017. [Google Scholar]
- Shao, F.; Wang, S.; Huang, W.; Liu, Z. Effects of IBA on the rooting of branch cuttings of Chinese jujube (Zizyphus jujuba Mill.) and changes to nutrients and endogenous hormones. J. For. Res. 2018, 29, 1557–1567. [Google Scholar] [CrossRef]
- Krajnc, A.U.; Ivanus, A.; Kristl, J.; Susek, A. Seaweed extract elicits the metabolic responses in leaves and enhances growth of Pelargonium cuttings. Eur. J. Hortic. Sci. 2012, 77, 170–181. [Google Scholar]
- Stirk, W.A.; Tarkowská, D.; Turečová, V.; Strnad, M.; Van Staden, J. Abscisic acid, gibberellins and brassinosteroids in Kelpak®, a commercial seaweed extract made from Ecklonia maxima. J. Appl. Phycol. 2014, 26, 561–567. [Google Scholar] [CrossRef]
- Aremu, A.O.; Plačková, L.; Gruz, J.; Bíba, O.; Novák, O.; Stirk, W.A.; Doleza, K.; Van Staden, J. Seaweed-derived biostimulant (Kelpak®) influences endogenous cytokinins and bioactive compounds in hydroponically grown Eucomis autumnalis. J. Plant Growth Regul. 2016, 35, 151–162. [Google Scholar] [CrossRef]
- Szabó, V.; Sárvári, A.; Hrotkó, K. Treatment of stockplants with biostimulators and their effects on cutting propagation of Prunus marianna ‘GF 8-1’. Acta Hortic. 2011, 923, 277–282. [Google Scholar] [CrossRef]
- Gomes, E.N.; Vieira, L.M.; Tomasi, J.D.C.; Tomazzoli, M.M.; Grunennvaldt, R.L.; Fagundes, C.D.M.; Machado, R.C.B. Brown seaweed extract enhances rooting and roots growth on Passiflora actinia Hook stem cuttings. Ornam. Hortic. 2018, 24, 269–276. [Google Scholar] [CrossRef] [Green Version]
- Faize, M.; Faize, L.; Burgos, L.; Critchley, A.; Albuquerque, N. Application of Ascophyllum nodosum-based soluble extract on micropropagation and regeneration of Nicotiana benthamiana and Prunus domestica. Plants 2021, 10, 1354. [Google Scholar] [CrossRef]
- Fleming, T.R.; Fleming, C.C.; Levy, C.C.; Repiso, C.; Hennequart, F.; Nolasco, J.B.; Liu, F. Biostimulants enhance growth and drought tolerance in Arabidopsis thaliana and exhibit chemical priming action. Ann. Appl. Biol. 2019, 174, 153–165. [Google Scholar] [CrossRef]
- Majda, M.; Robert, S. The role of auxin in cell wall expansion. Int. J. Mol. Sci. 2018, 19, 951. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Day | Cv. | Treatment | p | |||||
|---|---|---|---|---|---|---|---|---|
| H2O | AUX | BRA | KEL | PHY | T | C × T | ||
| 0 | M | 18.0 ± 1.27 | − | − | − | − | − | − |
| C | 20.8 ± 1.79 | − | − | − | − | − | − | |
| 4 | M | 8.0 ± 0.95 | 7.2 ± 1.79 | 10.0 ± 5.60 | 8.8 ± 2.19 | 7.9 ± 3.77 | ns | ns |
| C | 12.3 ± 2.00 | 14.3 ± 2.71 | 10.7 ± 1.23 | 12.6 ± 1.10 | 14.8 ± 4.00 | ns | ||
| 16 | M | 2.9 ± 1.16 ab | 3.8 ± 1.23 a | 0.3 ± 0.16 b | 0.8 ± 0.82 ab | 3.3 ± 1.37 ab | * | *** |
| C | 9.7 ± 1.84 a | 11.0 ± 2.19 a | 8.2 ± 2.26 a | 9.5 ± 2.34 a | 1.6 ± 0.16 b | ** | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Traversari, S.; Cacini, S.; Nesi, B. Seaweed Extracts as Substitutes of Synthetic Hormones for Rooting Promotion in Rose Cuttings. Horticulturae 2022, 8, 561. https://doi.org/10.3390/horticulturae8070561
Traversari S, Cacini S, Nesi B. Seaweed Extracts as Substitutes of Synthetic Hormones for Rooting Promotion in Rose Cuttings. Horticulturae. 2022; 8(7):561. https://doi.org/10.3390/horticulturae8070561
Chicago/Turabian StyleTraversari, Silvia, Sonia Cacini, and Beatrice Nesi. 2022. "Seaweed Extracts as Substitutes of Synthetic Hormones for Rooting Promotion in Rose Cuttings" Horticulturae 8, no. 7: 561. https://doi.org/10.3390/horticulturae8070561
APA StyleTraversari, S., Cacini, S., & Nesi, B. (2022). Seaweed Extracts as Substitutes of Synthetic Hormones for Rooting Promotion in Rose Cuttings. Horticulturae, 8(7), 561. https://doi.org/10.3390/horticulturae8070561